

Zachet_Abramova
.pdfБилет 1
1. основные свойства биологических систем
1.Самовозобновление, которое связано с постоянным обменом вещества и энергии, и в основе которого лежит способность хранить и использовать биологическую информацию в виде уникальных информационных молекул: белков и нуклеиновых кислот.
2.Самовоспроизведение, которое обеспечивает преемственность между поколениями биологических систем
3.Саморегуляция, которая основана на потоке вещества, энергии и информации
4.Большинство химических процессов в организме находятся не в динамичном состоянии
5.Живые организмы способны к росту
1.Обмен веществом и энергией
2.Обмен веществ – особый способ взаимодействия живых организмов со средой
3.Обмен веществ требует постоянного притока некоторых веществ и энергии из вне и выделения некоторых продуктов диссимиляции во внешнюю среду. Организм является открытой системой
4.Раздражимость – заключается в передаче информации от внешней среды к организму; на основе раздражимости осуществляется Саморегуляция и гомеостаз
5.Репродукция – воспроизведение себе подобных
6.Наследственность – поток информации между поколениями в результате чего обеспечивается преемственность
7.Изменчивость – появление новых признаков в процессе репродукции; основа эволюции
8.Онтогенез – индивидуальное развитие, реализация индивидуальной программы
9.Филогенез – историческое развитие, эволюционное развитие осуществляется в результате наследственной изменчивости, естественного отбора и борьбы за существование
10.Организмы включены в процесс эволюции
2. Что такое жизненный цикл клеток?
Закономерные изменения структурно-функциональных характеристик клетки во времени составляют содержание жизненного цикла клетки (клеточного цикла). Клеточный цикл — это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти.
Важным компонентом клеточного цикла является митотический (пролиферативный) цикл — комплекс взаимосвязанных и согласованных во времени событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления. Кроме того, в жизненный цикл включается период выполнения клеткоймногоклеточного организма специфических функций, а
также периоды покоя. В периоды покоя ближайшая судьба клетки не определена: она может либо начать подготовку к митозу, либо приступить к специализации в определенном функциональном направлении.
Билет 2 1. Генетический код и его свойства
Генети́ческий код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
Свойства генетического кода:
1.универсальность (принцип записи един для всех живых организмов)
2.триплетность (считываются три, рядом расположенные нуклеотида)
3.специфичность (1 триплет соответствует ТОЛЬКО ОДНОЙ аминокислоте)
4.вырожденность (избыточность) (1 аминокислота может кодироваться несколькими триплетами)
5.неперекрываемость (считывание происходит триплет за триплетом без "пробелов" и областей перекрывания, т.е. 1 нуклеотид НЕ может входить в состав двух триплетов).
1

2. Как образуются новые клетки? Чем заканчивается жизнь клеток?
Соматические клетки организма образуются в результате митоза. В дальнейшем возможны три варианта жизненного пути (цикла) клеток:
1.Клетки готовятся к делению и заканчивают свою жизнь митозом (митотический цикл).
2.Клетки дифференцируются, функционируют и погибают.
3.Клетки переходят в период G0, в котором могут находиться от нескольких часов до многих лет. При определенных условиях они могут перейти из этого периода в митотический цикл или терминальную дифференцировку.
Билет 3
1. Какие уровни организации биологических систем являются эволюционно обусловленными?
1.молекулярный
2.клеточный
3.тканевый
4.органный
5.системный
6.организменный
7.биогеоцианотический
8.ноосфера
2. Что такое митотический цикл? Из каких периодов он состоит? Что происходит в различные периоды митотического цикла?
Митоз— основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала. Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл. Ниже приводится краткая характеристика фаз цикла.
Интерфаза состоит из трех периодов: пресинтетического, или постмитотического,
— G1, синтетического — S, постсинтетического, или премитотического, — G2. Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул
ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду.
Синтетический период (2n 4c) — репликация ДНК.
Постсинтетический период (2n 4c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей.
Профаза (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом.
Метафаза (2n 4c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка),
2

прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза (4n 4c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами).
Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки.
Билет 4
1. Каковы особенности строения генов у про- и эукариот?
Признаки |
Прокариоты |
Эукариоты |
|
Количество генов |
4 тыс. (Е. coli) |
Около 30 тыс. (человек) |
|
Количество ДНК |
4 млн пар нуклеотидов |
3-7 млрд пар нуклеотидов |
|
Кодирующие |
|
|
|
последовательности |
Более 90% |
Менее 10% |
|
ДНК |
|
|
|
Связь ДНК с |
Отсутствует |
Формирует нуклеосомы |
|
гистонами |
|||
|
|
||
|
Кольцевая, содержит 100 |
Линейная с замкнутыми в |
|
Укладка ДНК |
петель по 40 тыс. пар |
теломеры концами, имеет 4 |
|
|
нуклеотидов |
уровня спирализации |
|
Количество |
Один |
50 тыс. |
|
репликонов |
|||
|
|
||
Активно работающие |
Более 90% генов |
Менее 10% генов |
|
участки |
|||
|
|
||
Процессинг |
Отсутствует |
Осуществляется при переходе |
|
пре-мРНК из ядра в цитоплазму |
|||
|
|
||
Регуляция |
Оперонная |
Сложная каскадная |
|
транскрипции |
|||
|
|
Основу генетического аппарата кишечной палочки составляет бактериальная хромосома, входящая в состав нуклеоида – ядерноподобной структуры. Нуклеоид по морфологии напоминает соцветие цветной капусты и занимает примерно 30% объема цитоплазмы. Бактериальная хромосома представляет собой кольцевую двуспиральную правозакрученную молекулу ДНК, которая свернута во вторичную спираль. Длина бактериальной хромосомы составляет примерно 4,7 млн. нуклеотидных пар (п.н.), или ~ 1,6 мм. Вторичная структура хромосомы поддерживается с помощью гистоноподобных (основных) белков и РНК. Точка прикрепления бактериальной хромосомы к мезосоме (складке плазмалеммы) является точкой начала репликации ДНК (эта точка носит название OriC). Бактериальная хромосома удваивается перед делением клетки, и сестринские копии распределяются по дочерним клеткам с помощью мезосомы. Репликация
3

ДНК идет в две стороны от точки OriC и завершается в точке TerC. Молекулы ДНК, способные себя воспроизводить путем репликации, называются репликоны.
Одна бактериальная хромосома содержит до 1000 известных генов. Обычно это гены «домашнего хозяйства», то есть необходимые для поддержания жизнедеятельности клетки.
Главная количественная особенность генетического материала эукариот – наличие избыточной ДНК. Этот факт легко выявляется при анализе отношения числа генов к количеству ДНК в геноме бактерий и млекопитающих. Если средний размер гена бактерий 1500 пар нуклеотидов (п.н.), а длина кольцевой молекулы ДНК хромосомы Е. coliи В. subtilis составляет свыше 1 мм, то в такой хромосоме могут разместиться около 3 тысяч генов. Примерно такое число генов было экспериментально определено у бактерий по числу типов иРНК. Если это число умножить на средний размер гена, то получится, что около 95% генома бактерий состоит из кодирующих (генных) последовательностей. Остальные 5%, повидимому, заняты регуляторными элементами. Иная картина наблюдается у эукариотических организмов. Например, у человека насчитывают приблизительно 50 тысяч генов (имеется в виду только суммарная длина кодирующих участков ДНК – экзонов). В то же время размер генома человека 3×109 (три миллиарда) п.н. Это означает, что кодирующая часть его генома составляет всего 15…20 % от тотальной ДНК. Существует значительное число видов, геном которых в десятки раз больше генома человека, например некоторые рыбы, хвостатые амфибии, лилейные. Избыточная ДНК характерна для всех эукариот. В этой связи необходимо подчеркнуть неоднозначность терминов генотип и геном. Под генотипом следует понимать совокупность генов, имеющих фенотипическое проявление, тогда как понятие генома обозначает количество ДНК, находящееся в гаплоидном наборе хромосом данного вида.
2. Каковы основные физико-химические свойства клетки?
Через оболочку и мембраны в клетку из внешней среды проникают различные растворенные вещества. Однако далеко не все вещества могут проникнуть в клетку. Живые клеточные оболочки и мембраны обладают избирательной проницаемостью: одни вещества проходят только в клетку, другие только из клетки, третьи вообще не проходят через этот барьер. Для демонстрации этого явления служит особый сосуд, называемый осмометром. Горлышко его плотно закрыто пробкой, в которой закреплена стеклянная трубка. Дно осмометра замещено полупроницаемой перепонкой (например, пленка из коллодия). Осмометр помешают в большой сосуд. В оба сосуда наливают жидкости, отличающиеся друг от друга концентрацией растворенных веществ, чистую воду и раствор какого-либо вещества, например красителя – метиленового синего.
Рассмотрим два случая. В первом случае дном осмометра является пленка с очень мелкими порами, пропускающими молекулы воды, но задерживающими молекулы метиленового синего)? В осмометр наливают 0,5% водный раствор метиленового синего, в большой сосуд – чистую воду. Поскольку в большом сосуде вода составляет 100%, а в осмометре только 99,5%, а концентрация растворов стремится к равновесию, будет происходить проникновение воды из большого сосуда в осмометр, т. е. будет совершаться осмос (жидкость в осмометре будет светлеть). Благодаря этому столбик жидкости в стеклянной
4
трубке будет подниматься до тех пор, пока его давление не сравняется с силой, заставляющей воду проходить через полупроницаемое дно осмометра (к).
Эту силу называют осмотическим давлением. Осмотическое давление прямо пропорционально концентрации растворенного вещества (в данном случае красителя), т. е. равно суммарному давлению всех молекул и ионов растворенного вещества на полупроницаемую перепонку.
Во втором случае попытаемся несколько приблизить опыт к явлениям, происходящим в живой клетке. В осмометре находится жидкость, в которой растворены крупные молекулы, например молекулы белка, а в большом сосуде – раствор метиленового синего. В этом опыте дно осмометра затянуто иной пленкой, с более крупными порами, пропускающими молекулы и воды,и метиленового синего, но задерживающими молекулы белка. Молекулы метиленового синего, проходя через поры этой пленки, распределяются равномерно по обе ее стороны. Общее число молекул белка и молекул метиленового синего в осмометре станет больше, вследствие чего в нем увеличится осмотическое давление (к).
В живом организме, как мы уже знаем, существуют полупроницаемые – клеточные и внутриклеточные мембраны, задерживающие большинство растворенных веществ.
Живая клетка в какой-то степени функционирует как осмотическая ячейка. Осмотическое давление в клетках и жидкостях определенного организма представляет собой более или менее постоянную величину – от 8 атм. у животных до 60 атм. у растений. У млекопитающих оно соответствует осмотическому давлению 0,85% раствора поваренной соли. Если концентрация солей в растворе, окружающем клетку, будет ниже этой величины, то и осмотическое давление в нем окажется ниже, чем в клетке. Такой раствор называется гипотоническим. Изза поступления воды клетка в нем будет набухать, а при достаточно большой разнице в осмотической концентрации внутри клеточной среды и вне ее она может вследствие разрыва погибнуть. Если клетка находится в растворе с повышенной концентрацией солей, в так называемом гипертоническом растворе, т. е. в растворе с более высоким осмотическим давлением, то из-за большой потери воды она съеживается. Подобное явление называется плазмолизом и приводит иногда даже к гибели клетки.
Если живую клетку или ткань поместить в солевой раствор, концентрация которого равна концентрации солей цитоплазмы (изотонический раствор), то она может некоторое время сохранять свою жизнеспособность. Такие изотонические растворы называют физиологическими. Для млекопитающих физиологическим раствором будет 0,85% раствор поваренной соли, для амфибий (лягушки)– 0,64% раствор этой же соли.
Клетка в нормальном состоянии обладает упругостью, зависящей от давления содержимого клетки на ее стенки, уравновешиваемого давлением эластичных стенок на содержимое (подобно надутой резиновой шине или мячу). Такое нормальное упругое состояние клетки называется тургором.
Тургор тем выше, чем выше осмотическое давление содержимого клетки.
5
Билет 5
1. Какие молекулярные процессы лежат в основе удвоения ДНК? Как происходит удвоение хромосом?
Репликация у прокариот.
Наиболее изучен процесс репликации у Escherichia coli. У этой бактерии (как и ещѐ некоторых исследованных видов) в области точки инициации репликации (ori C, длиной примерно 245 п.н.) находятся повторы размером в 13 и 9 пар оснований (Рис.5). При инициации 10-20 молекул белка инициации реплика¬ции Dna A связывается с четырьмя девятимерными повторами (9-mers) и расплетает ДНК в районе тандем¬ного набора тринадцатимеров, богатых АТ парами (что облегчает их расплетание, т.к. между А и Т только две водородные связи). Белок Dna C доставляет шестисубъединичный белок Dna B (геликаза) к матрице. На каж¬дую из одиночных цепей садится по одному Dna B и они затем двигаются в разных направлениях расплетая ДНК.
К геликазе присоединяется праймаза и синтезирует РНК-затравку. Две ДНК-полимеразы с помощью своих двух b-субъединиц прикрепляются к нити ДНК и начинают синтез ДНК. Расплетанию спирали способст¬вует SSB-белки, которые связываются с одноцепочечными участками ДНК, предотвращают образование шпи¬лек и тем са¬мым стабилизируют расплетѐнный дуплекс. Сбалансированное действие топоизомеразы II (гираза), способной индуцировать отрицательные сверхвитки(см.рис.4), и топоизомеразы I, снимающей отрицательные сверх¬витки(см.рис.3) регулирует степень сверхспиральности ДНК и таким образом влияет на скорость движе¬ния реп¬ликативной вилки.
У прокариот обнаружено три типа ДНК-полимераз. Их свойства приведены ниже.
ДНК-полимераза III осуществляет удлинение лидирующей цепи, а также удлинение РНКпраймеров с обра¬зова¬нием фрагментов Оказаки длиной от 1000 до 2000 нуклеотидов. Две ДНКполимеразы связаны между со¬бой субъединицей. Удаление сегментов РНК с 5 -конца каждого фрагмента Оказаки и заполнение пробелов между ними катализируетcя ДНК-полимеразой I ,способной удлинять цепь и осуществлять ник-трансляцию. Когда рас¬тущий 3 -гидро¬ксильный конец каждого фрагмента Оказаки до¬ходит до 5 –дезоксинуклеотид¬ного конца соседнего фраг¬мента, вступает в действие ДНКлигаза и образуется непре¬рыв¬ная отстающая цепь. Роль ДНК-полимеразы II в репликации не выяснена.
Обнаружен специальный белок терминации – Tus-белок. Он задерживает геликазу, в результате чего прекращается расплетение нити и происходит терминация репликации.
Репликация у эукариот.
Как и в случае с E.coli исследования репликации в эукариотических клетках сначала были сосредото¬чены на характеристике различных ДНК-полимераз (см. табл.2).
Следующим этапом стало создание систем для репликации хромосом вирусов животных in vitro. В ре¬зуль¬тате в настоящее время хромосома вируса SV40 может быть реплицирована in vitro с использованием всего лишь восьми компонентов клеток млекопитающих. По своим свойствам эти белки напоминают белки необхо¬димые для репликации в E.coli. Репликация ДНК эукариот также идѐт в двух направлениях; для син¬теза ДНК нужны праймеры синтезируемые праймазой; синтез лидирующей цепи непрерывен, а отстающей прерывистый. Как показано на рис.7, инициация репликации ДНК вируса SV40 происходит в уникальном сайте, точке начала репликации, путѐм связывания кодируемого вирусом белка, называемого T antigen, или Tag.
Этот полифункциональный белок расплетает дуплекс ДНК благодаря своей геликазной активности. Распле¬тание дуплекса требует также наличия АТФ и белка репликации A (RPA), кодируемого клеткой-хозяи¬ном и обладающего способностью связываться с однонитчатой ДНК (как SSB-белки в E.coli). Одна молекула ДНК-полимеразы α (Pol α) прочно связывается с праймазой и затем связывается с образовавшейся однонит¬чатой ДНК. Праймаза образует РНК-
6
праймеры, которые затем удлиняются на небольшую длину Pol α , обра¬зуя пер¬вую часть ведущих цепей, которые растут от точки ori в противоположных направлениях. Активность Pol α стимулируется фактором репликации C (RFC).
Затем c 3-концамb удлинѐнных Pol α РНК-праймеров связывается PCNA (proliferating cell nuclear antigen) и замещает Pol α на обоих растущих ведущих цепях, прерывая их синтез. На следующем этапе Pol δ связывается с PCNA на 3 -концах растущих цепей. PCNA повышает процессивность Pol δ так, что полимераза может непрерывно продолжать синтез ведущих цепей. Таким образом, функция PCNA аналогична функции β-субъединицы полимеразы III E.coli, т.к. оба белка образуют сходные структуры (―кольца‖), охватывающие ДНК и способствующие удержанию полимераз на цепи ДНК. Они, однако, имеют различные первичные структуры; кроме того PCNA-тример, а не димер как β-субъединица полимеразы III E.coli.
Комплекс праймазаPol α. садится на цепь, являющуюся матрицей для отстающей цепи и вместе с RFC осуще¬ствляют синтез запаздывающей цепи.
Наконец, как и в E.coli топоизомеразы сни¬мают меха¬ническое напряжение, возникающее при распле¬тании ДНК в репликативной вилке, и участ¬вуют в разделе¬нии двух дочерних хромосом. Од¬нако топои¬зомеразы эукариот имеют некоторые от¬личия от прокариотиче¬ских: 1.топоизомеразы I эукариот взаи¬модействуют с 3 -фосфорильным концом разорванной цепи (прока¬риотические --с 5 -фосфорильным кон¬цом) 2. топои-зомеразы I эукариот устраняют как отри¬цательные, так и положительные сверх витки (прокариотиче-ские—только отрицательные) 3.топоизомеразы II эу¬кариот не способны инду¬циро¬вать образование отрица¬тельных сверхвит¬ков (как это делает в релаксирован¬ных кольцевых ДНК гираза бактерий).
Итак, получено много данных об эукариотических белках, осуществляющих репликацию ДНК вируса SV40 in vitro. Как упоминалось ранее, инициация репликации ДНК SV40 in vitro требует наличие вирусного белка - T антигена. Для инициации же репликации у эукариот хромосомной ДНК необходим целый комплекс белков. Так, у дрожжей с сайтом ori в течение всего жизненного цикла связан комплекс из 6 разных белков (ORC), к которому в интерфазе присоединяется ещѐ целый ряд белков и образованный комплекс инициирует процесс репликации. Такие же белки синтезируются всеми эукариотическими клетками.
Хромосомы эукариот линейны и их концы представлены теломерами, со¬стоящими из повторяющихся олигомерных последо¬вательностей; у человека это 25-200 копий последовательности TTAGGG. Наличие специ¬альной области на концах эукариотических хро¬мосом абсолютно необходимо. Дело в том, что при удалении последнего РНК-праймера отстаю¬щей цепи, на 5- конце этой цепи остаѐтся брешь, которую не способна заполнить ни одна из ДНК-полимераз, т.к. всем им для работы необходим праймер со свободным 3-ОН концом. Без сущест¬вования какоголибо специального меха¬низма дочерняя нить ДНК, синтезируемая на от¬стающей цепи, укорачивалась бы с каждым клеточным де¬лением. Ферментом, предотвращаю¬щим такое укорочение, является теломераза. Этот фермент имеет ассоциированную с ним короткую нить РНК, комплиментарную шестичленной после¬до-вательности, повторяющейся в теломере и слу¬жащую матрицей для синтеза ДНК теломеров. Бла¬годаря этому механизму эукариотические хромо¬сомы могут реплицироваться полностью. Репликация в большинство соматических клеток проходит без участия теломеразы, поэтому с каж¬дым делением длина хромосом клетки укорачи¬ва¬ется и после определѐнного числа делений хромо¬сомы утрачивают теломеры и начинают терять смысловые участки , что приводит к гибели клетки. Теломераза активна в половых, раковых клетках и клетках одноклеточных эукариот.
G1 – 2n2c
S –
G2 – 2n4c
Gh – 2n2n
Митоз : 2n4n 2n2c
7

2. Основные принципы регуляции генной активности у эукариот?
Особенности регуляции генов у эукариот: 1)нет оперонной организации генов.2) Гены, определяющие синтез ферментов рассеяны в геноме. 3)Регуляция транскрипции является комбинационной, т.е. активность каждого гена регулируется большим числом генов-регуляторов. (промотор и энхансер) 4)белки-регуляторы контролируют транскрипцию генов, кодирующих другие белки-регуляторы 5)гормоны – индукторы транскрипции 6) процесс компактизации и декомпактизации хроматина 7) обратная связь между процессингом, сплайсингом и экзонинтронной организацией генов – например изменение схемы сплайсинга при синтезе антител.
Билет 6
1. Современное представление молекулярной организации биологической мембраны и ее функции?
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») часть. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Мембраны — структуры инвариабельные, весьма сходные у разных организмов. Некоторое исключение составляют, пожалуй, археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7—8 нм.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
2. Фазы митоза. Биологическая сущность и значение митоза. Биохимический процесс сопровождающий митоз.
Митоз— основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала. Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл. Ниже приводится краткая характеристика фаз цикла.
Интерфаза состоит из трех периодов: пресинтетического, или постмитотического,
— G1, синтетического — S, постсинтетического, или премитотического, — G2. Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул
ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду.
Синтетический период (2n 4c) — репликация ДНК.
Постсинтетический период (2n 4c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей.
Профаза (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом.
8
Метафаза (2n 4c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза (4n 4c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами).
Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки.
Биологическое значение митоза.
Митоз лежит в основе роста и вегетативного размножения всех организмов, имеющих ядро - эукариот. Благодаря митозу поддерживается постоянство числа хромосом в клеточных поколениях, т. е. дочерние клетки получают такую же генетическую информацию, которая содержалась в ядре материнской клетки.
По-видимому, на молекулярном уровне фаза М инициируется каскадомфосфорилирования белков, запускаемым при появлении М-стимулирующего фактора (MPF), и заканчивается при дефосфорилировании, которое возвращает белки в их интерфазное состояние (разд. 13.2.5). В свою очередь фосфорилирование белков в течение М-фазы, вероятно, ответственно за многие морфологические изменения, сопровождающие митоз, в том числе и за конденсацию хромосом, разрушение ядерной оболочки и изменения цитоскелета, описанные ниже. Первое хорошо видимое проявление наступающей фазы М состоит в постепенном уплотнении дисперсного интерфазного хроматина в нитевидные хромосомы. Эта конденсация хромосом необходима для их последующего упорядоченного расхождения в дочерние клетки и сопровождается фосфорилированием многочисленных молекул гистона П1, имеющихся в клетке (до шести фосфатных групп на одну молекулу Н1). Поскольку гистон П1 присутствует в количестве примерно одной молекулы на нуклеосому и известно, что он участвует в
упаковке нуклеосом (разд. 13.2.5), то его фосфорилирование киназой MPF (разд. 9.1.12) в начале фазы М должно бытьглавной причиной конденсации хромосом. Такое молекулярное объяснение, пока еще гипотетическое, показывает, на каком уровне в конечном счете должен описываться весь клеточный цикл.
Билет 7
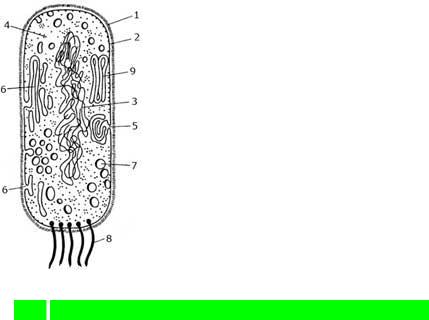
1. Как устроены прокариотические клетки?
Прокариотическая клетка устроена следующим образом. Главная особенность этих клеток — это отсутствие морфологически выраженного ядра, но имеется зона, в которой расположена ДНК (нуклеоид). В цитоплазме расположены рибосомы, цитоплазматические мембраны, но у них отсутствует набор других органелл, имеющихся в клетках эукариот, таких, как эндоплазматический ретикулюм, аппарат Гольджи, лизосомы, митохондрии, пластиды, центриоли, микротрубочки. Снаружи содержимое клетки прокариот одето цитоплазматической мембраной, которая играет барьерную функцию между собственно цитоплазмой клетки и внешней средой. Поверх цитоплазматической мембраны расположена клеточная стенка (оболочка). В то же время клетки прокариот и эукариот имеют и общие черты строения:
—одеты цитоплазматической мембраной, функционирующей как система для активного транспортирования веществ из клетки в клетку;
—синтез белка идет на рибосомах;
9
—сходны процессы синтеза РНК и репликации ДНК;
—похожи биоэнергетические процессы.
Прокариотическое строение клеток имеют все бактерии, включая архебактерии и цианобактерии (сине-зеленые водоросли). Клетки прокариот могут отличаться друг от друга по строению клеточной стенки, складчатости цитоплазматических мембран, количеству и свойствам внутриклеточных вакуолей, количеством и структурой цитоплазматических выростов и т. д., но общий план строения остается постоянным.
Прокариотическая клетка окружена мембраной, отделяющей цитоплазм образованной из сложного, высокополимерноголо,новеществаприсутств.В многочисленные мелкие рибосомы (бактериальные клетки соде
Строение прокариотической клетки
Цитоплазма прокариотической клетки пронизана мембранами, образующими эндоплазматическую сеть, в ней и находятся рибосомы, осуществляющие синтез белков.
Внутренняя часть клеточной стенки прокарио плазматической мембраной, выпячивания кото мезосомы, участвующие в построении клеточны являются местом прикрепления ДНК. Дыхание у мезосомах,-зеленыху синеводорослей в цитоплазматичес
У многих бактерий внутри клетки откладыв полисахариды, жиры, полифосфатывключаясь.Резервныеоб веществ, могут продлевать жизнь клетки в отсу
(1-клеточная стенка,-наружная 2 цитоплазматическаяхромосома(кольцевая молекула-рибосома,-мезосома,ДНК),5-впячивание4 6 наружной цитоплазмотической мембр-вакуоли,ны,-жгутики,7 8-стопки9 мембра которых осуществляется фотосинтез)
2. Роль факторов роста в функционировании клеточного цикла
Факторами роста называют группу белковых молекул, индуцирующих синтез ДНК в клетке (Goustin A.S. ea, 1986 ). Позднее было обнаружено, что спектр воздействий на клетки этих компонентов гораздо шире, чем предполагалось вначале. Так, некоторые белки этой группы в зависимости от типа клетокреспондентов могут индуцировать дифференцировку и подавлять пролиферацию . Кроме того, к ним относят регуляторные полипептиды, модулирующие подвижность клеток , но не
обязательно влияющие на деление клеток ( Stoker M. and Gherardi E., 1987 ).
ФР оказывают свое действие на различные фазы клеточного цикла. Известно, что клеточный цикл активно делящихся клеток состоит из четырех фаз *13+ (схема 1). Синтез ДНК происходит во время ограниченного периода интерфазы (S-фазы).
G1 - интервал между окончанием митоза и началом S-фазы. G2 - интервал между окончанием S-фазы и началом деления клетки, а М-фаза - период митоза и цитокинеза. Неделящиеся клетки находятся в состоянии покоя (G0-фазе). Клеточный цикл имеет две важнейшие точки регуляции, на которые оказывают воздействие ФР; эти точки различаются во времени и реагируют на разные митогены. Этими точками являются 1) ограничительная точка, в которой клетка нацеливается (становится обязанной) вступить в S-фазу; эта точка примерно на 2 ч предшествует началу синтеза ДНК; 2) присутствующая у неделящихся (с остановившимся ростом) клеток точка, расположенная в начале перехода от G0-фазы к G1-фазе *67+. ФРТ - фактор <дееспособности>, который стимулирует переход неделящихся клеток из фазы G0 в фазу G1 и их становление, таким образом, способными к делению. ИФР-I - фактор <прогрессии>, присутствие которого необходимо в G1-фазе, а
10