

Zachet_Abramova
.pdfтакже для вступления в S-фазу. ЭФР занимает промежуточное положение: он необходим в интервале между 2 и 6 ч после стимуляции ФР тромбоцитов выхода клетки из фазы G0. В отличие от этого ингибитор роста ТФР-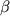 блокирует переход из фазы G0 в фазу G1.
блокирует переход из фазы G0 в фазу G1.
Билет 8
1. Как устроены эукариотические клетки
Клетка эукариот состоит из трех основных частей: ядра, цитоплазмы и клеточной стенки. К эукариотам относятся простейшие, беспозвоночные и позвоночные животные, высшие растения, грибы и водоросли (без сине-зеленых и прохлорофитовых). Клетки животных и растений отличаются по следующим параметрам. В клетках высших растений отсутствуют центриоли, они имеют жесткую клеточную стенку, плазмодесмы, вакуоль с клеточным соком, пластиды. В клетках водорослей, относящихся к разным таксонам, могут присутствовать или отсутствовать центриоли, клеточная стенка, пластиды и вакуоль с клеточным соком. Клетки грибов объединяют в себе некоторые признаки животных и растительных клеток. Как и клетки растений, они имеют жесткую клеточную стенку, но в ее состав входит хитин, как в наружном скелете у членистоногих. В клетках грибов отсутствуют пластиды, в обмене веществ у них присутствует мочевина, и запасают они не крахмал, а, как в клетках печени животных, гликоген. Оболочку, покрывающую клетку снаружи, называют клеточной мембраной. Внутри клетки часто встречаются пузырьки, оболочка которых очень похожа на клеточную мембрану. Их называют мембранными пузырьками, или вакуолями. Различные части клетки называются органоидами. На рисунке видны срезы нескольких органоидов: ядра, эндоплазматической сети (ЭПС), комплекса Гольджи, митохондрий, двух центриолей (вместе они имеют название
"клеточный центр"). Внутреннее содержимое клетки, за исключением ядра,
называют цитоплазмой.
Клетка живет активной жизнью. Шевелится мембрана, разные органоиды перемещаются с места на место, некоторые мембранные пузырьки сливаются в один пузырек, другие, наоборот, разделяются на несколько новых пузырьков. Если в роли большого пузырька выступает вся клетка, то при слиянии с ней маленького пузырька его содержимое выбрасывается наружу. Эту ситуацию выброса из клетки "начинки" мембранного пузырька называют экзоцитозом, а ситуацию захвата чего-либо внутрь клетки - эндоцитозом (от слов "эндо-" - "внутрь" и "экзо-" - "наружу").
Любая живая клетка питается, т.е. захватывает из внешней среды съедобные для себя вещества (в виде отдельных молекул или больших групп молекул - пищевых частиц, иногда даже целых клеток меньшего размера), и так или иначе использует эти вещества.
2. Что такое митотический коэффициент? Как он определяется?
Митотический индекс (Mitotic index,MI, %) — процент делящихся клеток от общего числа проанализированных клеток. Данный индекс можно вычислить используя световой микроскоп, просчитав в поле зрения клетки с видимыми хромосомами и разделив его на общее число клеток в поле зрения.
11
Если вы управляете колхицином или другим лекарством — производным колхицина (например, colcemid), вы можете остановить клеточный цикл в этот момент и оставить хромосом в их видимой формы. Колхицин нарушает образование микротрубочек, которое необходимы для шпинделя волокон отдельных хромосом в анафазе.
Рост клеточной культуры происходит когда клетки проходят через интерфазу и митоз для завершения клеточного цикла. Многие клетки теряют способность делиться когда стареют или делятся реже. Другие клетки способны к быстрому делению. Например, как корни растений, клетки вблизи от кончика корня, в апикальной меристеме, делятся быстро внедряя корень в почву. Корневой чехлик определяет направление гравитации и устремляет свою вершину с интенсивно делящимися клетками вниз.
Количественно мы можем сравнить группы клеток по их способности к делению. В экспериментальных условиях мы можем изменять условия среды и количественное определить это воздействие на пролиферативную способность клеток.
Для группы клеток, редко совершающих клеточный цикл мы прогнозируем что значительная часть будет находиться в стадии покоя клеточного цикла (G1). Однако в быстро делящейся клеточной культуре мы можем ожидать увидеть большую долю клеток в стадии митотического деления. Одним из верных способов количественного анализа интенсивности деления служит митотический индекс.
Показывает интенсивность деления по наличию клеток в фазе роста (делящихся клеток). Чем выше значение, тем интенсивнее происходит процесс деления клеток и наоборот. Индекс может говорить о нормальном протекании митоза, об угнетении процесса деления клеток или, напротив, усилении митотической активности тканей. На основании этого делается заключение о митотическом или митозстимулирующем действии изучаемого фактора.
Билет 9
1. Гипотезы происхождения эукариотических клеток
Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой (рис. 1.4) основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней.
Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза.
Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клеткихозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли.
Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот (рис. 1.4). Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной
12

оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран. Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки.
2. Чем отличаются жизненные циклы нормальных и опухолевых клеток?
Спонтанно или при действии канцерогенных факторов могут происходить мутации протоонкогенов или генов супрессоров, регулирующих размножение клеток. Протоонкогены превращаются в онкогены, которые не реагируют на регуляторные факторы и образуют большое количество факторов роста. Повреждение геновсупрессоров не позволяет сдерживать избыточное размножение клеток - возникает опухоль. Для клеток опухоли характерна генетическая нестабильность - в них возникают новые мутации, которые еще больше нарушают регуляцию клеточной пролиферации.
Доброкачественная опухоль может трансформироваться в злокачественную.
Параметры |
Нормальные клетки |
Опухолевые клетки |
||
|
Генетический |
|
Осуществляется |
Трансформация протоонкогенов в |
|
|
генамиактиваторами |
||
|
контроль |
|
онкогены; подавление |
|
|
|
(протоонкогенами) и генами |
||
|
пролиферации |
|
геновсупрессоров |
|
|
|
супрессорами |
||
|
|
|
|
|
|
|
|
Сбалансированная работа |
|
|
Регуляция |
|
генов-активаторов и |
Нарушается: происходит |
|
клеточного |
|
супрессоров поддерживает |
прогрессивное увеличение |
|
цикла |
|
оптимальный уровень |
количества клеток |
|
|
|
пролиферации |
|
|
Пролиферативный |
Постоянный для каждой ткани |
Прогрессивно увеличивается |
|
|
пул |
|
||
|
|
|
|
|
|
Межклеточные |
|
Ограничивают увеличение |
Нарушены: нет контактного |
|
|
количества клеток при |
||
|
контакты |
|
торможения пролиферации |
|
|
|
контактном торможении |
||
|
|
|
|
|
|
|
|
Обеспечивает возможность |
|
|
|
|
размножения клеток при |
Изменена: возможно размножение |
|
Клеточная мембрана |
контакте с базальной |
клеток без контакта с опорными |
|
|
|
|
мембраной или другими |
структурами |
|
|
|
опорными структурами |
|
|
Адгезия клеток |
|
Нормальная |
Снижена: возможен отрыв клеток |
|
|
и метастазирование |
||
|
|
|
|
|
|
Временная |
|
Одновершинный суточный |
Нарушение ритма митозов: |
|
|
двувершинный, инвертированный, |
||
|
характеристика |
|
ритм митозов |
|
|
|
отсутствие ритма |
||
|
|
|
|
|
|
Пространственная |
|
Нарушена вследствие утраты |
|
|
Строго определенная |
контроля пролиферации и |
||
|
организация |
|
||
|
|
|
изменения клеточных контактов |
|
|
|
|
|
|
|
Деление клеток |
|
Митоз |
Значительное количество |
|
|
нарушений митозов, амитозы |
||
|
|
|
|
|
|
|
|
|
13 |
Набор хромосом |
Строго определенный |
Значительные изменения числа и |
|
(кариотип) |
структуры хромосом |
||
|
Билет 10
1. Особенности наследственного материала у про- и эукариот
Геном современных прокариотических клеток характеризуется относительно небольшими размерами. У кишечной палочки (Е. coli) он представлен кольцевой молекулой ДНК длиной около 1 мм, которая содержит 4·106 пар нуклеотидов, образующих около 4000 генов. Основная масса ДНК прокариот (около 95%) активно транскрибируется в каждый данный момент времени. Как было сказано выше, геном прокариотической клетки организован в виде нуклеоида — комплекса ДНК с негистоновыми белками (см. разд.
3.5.2.4).
У эукариот объем наследственного материала значительно больше. У дрожжей он составляет 2,3 · 107 п.н., у человека общая длина ДНК в диплоидном хромосомном наборе клеток — около 174 см. Его геном содержит 3·109 п.н. и включает по последним данным 30—40 тыс. генов.
Большой объем наследственного материала эукариот объясняется существованием в нем помимо уникальных также умеренно и высоко повторяющихся последовательностей. Так, около 10% генома мыши составляют тандемно расположенные (друг за другом) короткие нуклеотидные последовательности, повторенные до 106 раз. Эти высоко повторяющиеся последовательности ДНК располагаются в основном в гетерохроматине, окружающем центромерные участки. Они не транскрибируются. Около 20% генома мыши образовано умеренными повторами, встречающимися с частотой 103—105 раз. Такие повторы распределены по всему геному и транскрибируются в РНК. К ним относятся гены, контролирующие синтез гистонов, тРНК, рРНК и некоторые другие. Остальные 70% генома мыши представлены уникальными нуклеотидными последовательностями. У растений и амфибий на долю умеренно и высоко повторяющихся последовательностей приходится до 60% генома.
Избыточность генома эукариот объясняется также экзон-интронной организацией большинства эукариотических генов, при которой значительная часть транскрибированной РНК удаляется в ходе следующего за синтезом процессинга и не используется для кодирования аминокислотных последовательностей белков (см. разд.
3.4.3.2).
Характеризуя наследственный материал прокариотической клетки в целом, необходимо отметить, что он заключен не только в нуклеоиде, но также присутствует в цитоплазме в виде небольших кольцевых фрагментов ДНК — плазмид.
Плазмиды — это широко распространенные в живых клетках внехромосомные генетические элементы, способные существовать и размножаться в клетке автономно от геномной ДНК. Описаны плазмиды, которые реплицируются не автономно, а только в составе геномной ДНК, в которую они включаются в определенных участках. В этом случае их называют эписомами.
Впрокариотических (бактериальных) клетках обнаружены плазмиды, которые несут наследственный материал, определяющий такие свойства, как способность бактерий к конъюгации, а также их устойчивость к некоторым лекарственным веществам.
Вэукариотических клетках внехромосомная ДНК представлена генетическим аппаратом органелл — митохондрий и пластид, а также нуклеотидными последовательностями, не являющимися жизненно необходимыми для клетки (вирусоподобными частицами).
Наследственный материал органелл находится в их матриксе в виде нескольких копий
14

кольцевых молекул ДНК, не связанных с гистонами. В митохондриях, например содержится от 2 до 10 копий мтДНК.
Внехромосомная ДНК составляет лишь небольшую часть наследственного материала эукариотической клетки. Например, мтДНК человека содержит 16569 п.н. и на её долю приходится менее 1% всей клеточной ДНК.
В отличие от хромосомной ДНК, мтДНК характеризуется высокой «плотностью генов». В них нет интронов, а межгенные промежутки невелики. В кольцевой мтДНК человека содержится 13 генов, кодирующих белки (3 субъединицы цитохром С-оксидазы, 6 компонентов АТФазы и др.) и 22 гена тРНК. Значительная часть белков митохондрий и пластид синтезируется в цитоплазме под контролем геномной ДНК.
Если большинство ядерных генов представлены в клетках организма в двойной дозе (аллельные гены), то митохондриальные гены представлены многими тысячами копий па клетку.
Для генома митохондрий характерны межиндивидуальные различия, но в клетках одного индивида, как правило, мтДНК идентична.
Совокупность генов, расположенных в цитоплазматических молекулах ДНК, называют плазмоном. Он определяет особый тип наследования признаков — цитоплазматическое наследование (см. разд. 6.3.2.).
2. Механизмы регуляции клеточного деления
Прохождение клетки по всем периодам клеточного цикла строго контролируется. При движении клеток по клеточному циклу в них появляются и исчезают, активируются и ингибируются специальные регуляторные молекулы, которые обеспечивают: 1) прохождение клетки по определенному периоду клеточного цикла и 2 переход из одного периода в другой. Причем прохождение по каждому периоду, а также переход из одного периода в другой контролируется различными веществами. Сейчас мы попробуем выяснить, что же это за вещества и что они делают.
Общая ситуация выгладит так. В клетке постоянно присутствуют специальные белки-ферменты, которые путем фосфорилирования других белков (по остаткам серина, тирозина или треонина в полипептидной цепи), регулируют активность генов, ответственных за прохождение клетки по тому или иному периоду клеточного цикла. Эти белки-ферменты называются циклин-зависимыми протеинкиназами (cdc). Имеется несколько их разновидностей, но они все обладают сходными свойствами. Хотя количество этих циклин-зависимых протеинкиназ может варьировать в различных периодах клеточного цикла, они присутствуют в клетке постоянно, независимо от периода клеточного цикла, то есть они имеются в избытке. Другими словами, их синтез или количество не лимитирует или не регулирует прохождение клеток по клеточному циклу. Однако при патологии, если синтез их нарушен, снижено их количество или имеются мутантные формы с измененными свойствами, то это, конечно же, может повлиять на течение клеточного цикла.
Почему же такие циклин-зависимые протеинкиназы сами не могут регулировать прохождение клеток по периодам клеточного цикла. Оказывается, что они находятся в клетках в неактивном состоянии, а для того чтобы они активировались и начали работать, необходимы специальные активаторы. Ими являются циклины. Их также много разных типов, но они присутствуют в клетках не постоянно: то появляются, то исчезают. В разные фазы клеточного цикла образуются разные циклины, которые связываясь с Cdk образуют различные Cdk-циклиновые комплексы. Эти комплексы регулируют разные фазы клеточного цикла и поэтому называются G1-, G1/S-, S- и М-Cdk (рис. из моих рис. циклины). Так, например, прохождение клетки по G1 периоду клеточного цикла обеспечивает комплекс циклин-зависимой протеинкиназы-2 (cdk2) и циклина D1, циклин-зависимой протеинкиназы-5 (cdk5) и циклина D3. Прохождение через специальную точку рестрикции (R-пункт) периода G1 контролирует комплекс cdc2 и циклина С. Переход клетки из G1 периода клеточного цикла в S период контролирует комплекс cdk2 и циклина Е. Для перехода клетки из S периода в G2 период необходим комплекс cdk2 и циклин А. Циклин-зависимая протеинкиназа-2 (cdc2) и циклин В участвуют в переходе клетки из G2 периода в митоз (М период). Циклин H в соединении с cdk7 необходим для фосфорилирования и активации cdc2 в комплексе с циклином В.
15

Билет 11 1. Что такое жизненный цикл клетки?
Клеточный цикл – это период жизни клетки от одного деления до другого или от деления до смерти. Клеточный цикл состоит из интерфазы (период вне деления) и самого клеточного деления.
Клеточный цикл разделяется на четыре периода:
*G1 фаза (от "gap 1", то есть интервал 1) - период высокой метаболической активности и роста клетки между митозом и репликацией ДНК.
*S фаза (от "synthesis") - период синтеза (репликации ДНК). Количество ядерной ДНК увеличивается в два раза от 2n до 4n.
*G2 фаза ("gap 2") - период подготовки к митозу. Продолжается клеточный рост и синтез необходимых белков
*M фаза (от "mitosis") - деление клетки на две дочерние с уменьшением в них количества ДНК от 4n до 2n. Митоз - сложный процесс, в ходе которого происходит конденсация хроматина с образованием узнаваемых хромосом, перемещение центриолей в противоположные части клетки, реорганизация и реконструкция микротрубочек в веретено деления, разрыв ядерной оболочки, вызванный деполимеризацией поддерживающих ее структуру белков - ламинов (профаза), прикрепления микротрубочек к кинетохорам хромосом(прометафаза), выстраивание хромосом по клеточному экватору (метафаза), расхождение сестринских хроматид к противоположным полюсам клетки(анафаза), реорганизация ядерных оболочек и
непосредственное деление цитоплазмы (телофаза) и клетки (цитокинез). Типичные быстро делящиеся клетки человека (клетки костного мозга, слизистой
оболочки кишечника, волосяных фолликул и др.) проходят клеточный цикл за 24 часа. Быстро делятся клетки эмбриона - сразу после оплодотворения яйцеклетки. Они проходят клеточный цикл за 30 мин, однако при этом не происходит клеточный рост, то есть зигота делится на большое количество маленьких клеток. Некоторые клетки взрослого организма либо прекращают делиться вообще (например, нервные клетки), либо делятся случайно, для замены погибших или поврежденных в результате ранения клеток при получении сигнала извне (например, фибробласты кожи). Такие клетки вышли из фазы G1 в фазу покоя G0, оставаясь при этом метаболически активными. Клеточный цикл других клеток может остановиться и в G2 фазе. В диплоидном мире эти клетки в дальнейшем обычно остаются тетраплоидными, как это бывает, например, на некоторых стадиях эмбриогенеза насекомых. В гаплоидном мире остановка клеточного цикла в G2 фазе происходит всегда, так как это позволяет предохранять ДНК от повреждений из-за наличия в клетке двух ее копий, вместо одной копии в G1 фазе.
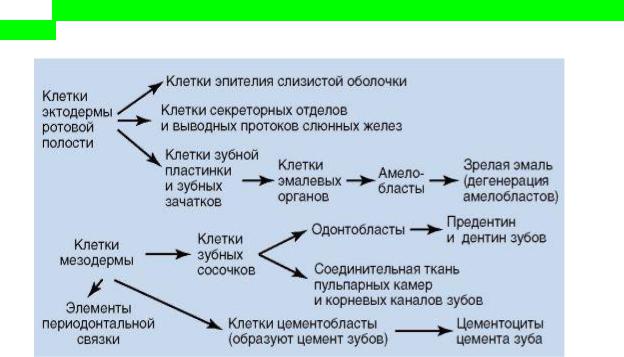
2. Что такое стволовые клетки? Виды и значения
Стволовые клетки являются основным источником клеточного материала быстро обновляющихся тканей, таких как кровь и эпителий. Но и в тканях, которым свойственна медленная смена клеточного состава, обнаружены стволовые клетки, которые отвечают за их восстановление, такие клетки находятся в состоянии покоя, активируясь лишь при определенных сигналах, например при повреждении органа. Стволовые клетки представлены в очень небольшом количестве и благодаря особому микроокружению, сохраняют
16
свой недифференцированный статус. При активации и делении, тканеспецифичные стволовые клетки дают начало дочерним клеткам – потомкам, которые пролиферируют (проходят несколько этапов деления) и дифференцируются, восстанавливая поврежденный участок, или восполняя клеточный состав активно обновляющейся ткани.
|
Способны давать |
С эмбриональных тотипотентных клеток |
|
|
начало любому виду |
начинается развитие организма при |
|
Тотипотентные |
клеток (бластомеры |
половом размножении. Соматические |
|
|
на ранних этапах |
дают начало новым организмам при |
|
|
дробления) |
вегетативном размножении |
|
|
Способны давать |
Формирование органов и тканей |
|
|
разные виды клеток |
развивающегося организма. Необходимы |
|
Полипотентные |
(клетки зародышевых |
для обновления или регенерации тканей, |
|
(плюрипотентные) |
листков; клетки |
в которых нет собственных стволовых |
|
|
красного костного |
клеток - эритроцитов и лейкоцитов, |
|
|
мозга) |
нейронов, кардиомиоцитов |
|
|
При размножении |
|
|
|
образуют клетки |
|
|
Унипотентные |
только одного вида |
Источник клеток для роста, обновления и |
|
(эпителий ротовой |
регенерации органов |
||
|
|||
|
полости, слюнных |
|
|
|
желез) |
|
|
|
Выделенные |
Использование в медицине позволяет |
|
|
эмбриональные |
выращивать органы и ткани с заданными |
|
Реконструированные |
стволовые клетки, в |
свойствами. Их применение для |
|
эмбриональные |
которых методами |
репродуктивного клонирования является |
|
|
генной инженерии |
источником генномодифицированных |
|
|
изменен состав генов |
организмов |
Билет 12
1. Молекулярная организация и функции нуклеиновых кислот?
Принципы строения ДНК
1.Нерегулярность. Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование нерегулярно.
2.Антипараллельность. ДНК состоит из двух полинуклеотидных цепей, ориентированных антипа-раллельно, т.е. идущих навстречу друг другу. 3'-конец одной цепи расположен напротив 5-конца другой.
3.Комплементарность (дополнительность). Каждому азотистому основанию одной цепи соответствует строго определенное азотистое снование другой цепи. Соответствие их друг другу задается химической структурой оснований. Пурины - более длинные основания, и поэтому для сохранения равномерной толщины двойной цепи они могут связываться только с более короткими пиримидинами. Пурин и пиримидин в паре образуют водородные связи. В паре А-Т две водородные связи, в паре Г-Ц — три.
17

4. Наличие регулярной вторичной структуры. Две комплементарные,
антипараллельно расположенные полинуклеотидные цепи образуют правые спирали с общей осью.
Формы двойной спирали ДНК
Существуют несколько форм двойной спирали ДНК.
В основной - В-форме на виток приходится 10 комплементарных пар.
Плоскости |
азотистых |
оснований |
|
перпендикулярны |
оси |
||
спирали. Соседние комплемен-тарные пары повернуты |
друг относительно друга на 36о. |
||||||
Диаметр |
спирали |
20Å, |
причем |
пуриновый |
нуклеотид |
занимает |
|
12Å, а пиримидиновый - 8Å. |
|
|
|
|
|
||
А-форма - 11 пар азотистых оснований на виток. |
Плоскости азотистых оснований |
||||||
отклонены от нормали к оси спирали на 20°. Отсюда следует наличие внутренней пустоты диаметром 5Å. Высота витка 28Å. Такие же параметры у гибрида из одной цепи ДНК и одной цепи РНК.
С-форма - шаг спирали 3lÅ, 9,3 пар оснований на виток, угол наклона к перпендикуляру 6°.
Все три формы - правозакрученные спирали.
Есть еще несколько форм правых спиралей и всего одна левая спираль (Z - форма). Высота витка в Z-форме -44.5Å, на виток приходится 12 пар нуклеотидов. Ни А-, ни Z- формы не могут существовать в водном растворе без дополнительных воздействий (белки или суперспирализация).
Структура рибонуклеиновых кислот (РНК)
Первичная структура РНК - порядок чередования рибонуклеозидмонофосфатов (НМФ) в полинуклеотидной
цепи. В РНК, как и в ДНК, нук-леотиды связаны между собой 3',5'-фосфодиэфирными связями. Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная ОН-группа 5'-углеродного атома, на другом конце - ОН-группа 3'- углеродного атома рибозы, поэтому концы называют 5'- и 3'-концами цепи РНК. Гидроксильная группа у 2'-углеродного атома рибозы делает молекулу РНК нестабильной. Так, в слабощелочной среде молекулы РНК гидролизуются даже при нормальной температуре, тогда как структура цепи ДНК не изменяется.
Вторичная структура РНК
Молекула рибонуклеиновой кислоты построена из одной полинуклеотидной цепи. Отдельные участки цепи РНК образуют спирализованные петли - "шпильки", за счёт водородных связей между комплементарными азотистыми основаниями A-U и G-C. Участки цепи РНК в таких спиральных структурах антипараллельны, но не всегда полностью комплементарны, в них встречаются неспаренные нуклеотидные остатки или даже одноцепочечные
146
18
петли, не вписьюающиеся в двойную спираль. Наличие спирализованных участков характерно для всех типов РНК.
Третичная структура РНК
Одноцепочечные РНК характеризуются компактной и упорядоченной третичной структурой, возникающей путём взаимодействия спирализованных элементов вторичной структуры. Так, возможно образование дополнительных водородных связей между нуклеотидными остатками, достаточно удалёнными друг от друга, или связей между ОНгруппами остатков рибо-зы и основаниями. Третичная структура РНК стабилизирована ионами двухвалентных металлов, например ионами Mg2+, связывающимися не только с фосфатными группами, но и с основаниями.
2. Клеточные циклы и направления дифференцировки при образовании тканей и органов
У животных дифференцировка интенсивно происходит при зародышевом развитии, а также в постэмбриональный период, пока организм растѐт и развивается. Клеточные дифференцировки идут и у взрослого организма, когда, напр., в кроветворных органах стволовые клетки дифференцируются в постоянно обновляющиеся клетки крови, а в половых органах первичные половые клетки – в гаметы. В отличие от животных, растения растут всю жизнь, и, следовательно, образование новых органов и тканей у них идѐт до тех пор, пока они существуют. Эти процессы обеспечиваютсяобразовательными тканями, или меристемами. Меристемы состоят из неспециализированных, внешне одинаковых клеток, которые в ходе многократных делений дифференцируются и дают начало различным тканям и органам растения.
Клеточные процессы дифференцировки определяются заключѐнными в генах программами. Так как все соматические клетки развивающегося зародыша содержат одну и ту же генетическую информацию, возникновение из генетически однотипных клеток таких различно специализированных клеток, как, напр., клетки мозга, мышц, кожи у животных или клетки листьев и корней у растений, можно объяснить только работой в них различных генов или т. н. дифференциальной экспрессией (активностью) генов. Сложные молекулярные и клеточные механизмы, регулирующие включение и выключение разных генов и направляющие клетки по различным путям дифференцировки, изучены недостаточно.
Ранее считалось, что дифференцировка соматических клеток, особенно клеток высших животных, необратима. Однако успехи таких методов, как культура клеток и тканей и клонирование,
19

показали, что в ряде случаев дифференцировка обратима: при определѐнных условиях из специализированной клетки можно вырастить полноценный организм.
Билет 13
1. Что такое ген и его определение?
Ген (от греч. génos — род, происхождение), элементарная единица наследственности, представляющая отрезок молекулы дезоксирибонуклеиновой кислоты — ДНК (у некоторых вирусов — рибонуклеиновой кислоты — РНК). Каждый Г. определяет строение одного из белков живой клетки и тем самым участвует в формировании признака или свойства организма.
2. Каковы биологическое значение и сущность мейоза?
Значение
У организмов, размножающихся половым путем, предотвращается удвоение числа хромосом в каждом поколении, так как при образовании половых клеток мейозом происходит редукция числа хромосом.
Мейоз создает возможность для возникновения новых комбинаций генов (комбинативная изменчивость), так как происходит образование генетически различных гамет.
Редукция числа хромосом приводит к образованию "чистых гамет", несущих только один аллель соответствующего локуса.
Расположение бивалентов экваториальной пластинки веретена деления в метафазе 1 и хромосом в метафазе 2 определяется случайным образом. Последующее расхождение хромосом в анафазе приводит к образованию новых комбинаций аллелей в гаметах. Независимое расхождение хромосом лежит в основе третьего закона Менделя.
Билет 14
1. Особенности строения генов у про и эукариот?
Геном современных прокариотических клеток характеризуется относительно небольшими размерами. У кишечной палочки (Е. coli) он представлен кольцевой молекулой ДНК длиной около 1 мм, которая содержит 4·106 пар нуклеотидов, образующих около 4000 генов. Основная масса ДНК прокариот (около 95%) активно транскрибируется в каждый данный момент времени. Как было сказано выше, геном прокариотической клетки организован в виде нуклеоида — комплекса ДНК с негистоновыми белками (см. разд.
3.5.2.4).
У эукариот объем наследственного материала значительно больше. У дрожжей он составляет 2,3 · 107 п.н., у человека общая длина ДНК в диплоидном хромосомном наборе клеток — около 174 см. Его геном содержит 3·109 п.н. и включает по последним данным 30—40 тыс. генов.
Большой объем наследственного материала эукариот объясняется существованием в нем помимо уникальных также умеренно и высоко повторяющихся последовательностей. Так, около 10% генома мыши составляют тандемно расположенные (друг за другом) короткие нуклеотидные последовательности, повторенные до 106 раз. Эти высоко повторяющиеся последовательности ДНК располагаются в основном в гетерохроматине, окружающем центромерные участки. Они не транскрибируются. Около 20% генома мыши образовано умеренными повторами, встречающимися с частотой 103—105 раз. Такие повторы
20