

7.5. Осіменіння корів-донорів, запліднення та ранні стадії розвитку ембріонів
F2cc
У корів, оброблених гонадотропінами на 15-17-й день циклу, охота наступає через 5-5 днів, а при комбінованій стимуляції гонадотропінами та ПГ-р2а в лютеїнову стадію циклу — через 75 годин після введення простагландину. При появі у тварин охоти а осіменяють.
Приблизно у 10 % донорів статевий цикл буває алібідним, хоча овуляція відбувається.
Осіменяють донорів цервікально, з ректальною фіксацією шийки матки, з використанням стерильних одноразових інструментів. У кожній дозі сперми повинно бути ж менше 50-100 млн рухливих сперміїв.
Оскільки охота та овуляція у оброблених гонадотропінами корів розтягується до 56-48 годин, то їх осіменяють 3^4 рази з інтервалами по 10-12 годин.
Оброблені гонадотропінами тварини відрізняються підвищеною чутливістю статевих органів до ректальної пальпації та інфекції, тому осіменяти їх слід обережно. Необережні маніпуляції можуть викликати зміщення бахромки яйцепроводу, тоді не зсі яйцеклітини попадають в його просвіт. Крім того, необережна пальпація яєчника ■ке викликати передчасні розриви передовуляційних фолікулів і вихід недозрілих
ЇЙЦЄКЛІТИН.
Введені в цервікальний канал спермії звичайно зберігають тут свою живучість до рак діб, проте під впливом гормональних обробок донорів змінюється якість церві-• ільного секрету, а отже і живучість, і запліднююча здатність сперміїв, що потрібно
211
Розділ 7
мати на увазі при осіменінні донорів. Просуваючись поступово в напрямку яйцепроводів спермії піддаються капацитації та селекції найбільш життєздатних.
Яйцеклітини, що попали після овуляції у яйцепровід, рухаються разом з течією рідини в напрямку рога матки. У верхній третині яйцепроводу вони зустрічаються зі сперміями і при сприятливих умовах переважна більшість з них запліднюється. Запліднена яйцеклітина містить подвійний набір хромосом, тому вона називається зиготою. Лімітуючим фактором відповідних змін сперміїв, яйцеклітин і самого запліднення є середовище яйцепроводу.
Процес дальшого розвитку зиготи полягає перш за все в поділі її ядра, спочатку на дві половинки (два бластомери), тоді на чотири, вісім і т. д., перетворюючись спочатку в ембріони, а тоді в плоди.
Програма розвитку ембріона закодована в його ДНК. Послідовність та специфічність всіх його змін залежить від часу зняття блокади окремих детермінант, що були раніше в неактивному стані. Кожна нова структура ембріона несе на своїй поверхні нову антигенну інформацію, від якої може залежати гальмування одних та стимуляція інших ділянок.
Приблизно через кожних 24 години кількість бластомерів у ембріоні подвоюється. Прозора та жовткова оболонки при цьому деякий час зберігаються, загальний діаметр ембріона залишається таким же, проте кожні наступні бластомери виявляються вдвічі меншими попередніх, чому цей їх поділ ще називають дробленням.
Ранні ембріони затримуються 3^4 дні в яйцепроводах, цьому сприяє наявність між яйцепроводом та рогом матки перешийка, що виконує роль адренергічного сфінктера. В постовуляційну стадію під впливом прогестерону та ПГЕ сфінктер розслаблюється.
За час перебування ембріонів у яйцепроводі матка за допомогою фагоцитозу звільнюється від залишків сперми, бактерій і т. п., в ендометрії відбуваються проліфера-тивні зміни, підвищується активність маткових залоз, формується гістіотроф.
На 3-4-й день ембріон переміщається з яйцепроводу у матку - у великої рогатої худоби на стадії 8-16-ти бластомерів; у свиней- на стадії 4-х бластомерів; кобил на стадії бластоцисти. Бластомери в цей час тісно прилягають один до одного у вигляді шовковиці чи тутової ягоди, тому їх називають (ранньою) морулою. Кількість бластомерів збільшується і на 4-5-ту добу - це морула (16-32 бластомери), а на 6-ту-7-му - пізня морула (32-90 бластомерів).
На ранніх стадіях розвитку ембріона всі бластомери знаходяться в однакових умовах, проте з часом вони диференціюються стосовно до їх локалізації, дещо змінюється темп їх дроблення. Із зовнішнього шару бластомерів, що діляться дещо швидше, формується живильний шар - трофобласт, що дасть початок судинній оболонці, а з розміщеного в центрі грудки бластомерів, що діляться повільніше, - ембріобласт, з якого з часом - розів'ється власне зародок (ембріон, а згодом плід) з водною та сечовою оболонками.
На 7-8-му добу між трофобластом та ембріобластом з'являється щілина, наповнена рідиною - бластоцель, навколо якої перегруповуються бластомери: більші з них (ембріобласт) зосереджуються на одному полюсі, а дрібніші - на протилежному. Ран-
212
Трансплантація ембріонів
ня бластоциста містить від 90 до 120-ти бластомерів, поверхня клітинної маси гладенька, рівномірна, товщина прозорої оболонки - біля 12 мк.
В наступні дні відбувається дальше збільшення кількості бластомерів до 300-^-80 на стадії бластоцисти і 1 200-1 500 на стадії пізньої бластоцисти. Дещо збільшується їх діаметр (до 140-200 мк), тоншає прозора оболонка, розширюється порожнина бластоцисти, вона займає всю прозору оболонку і нарешті на 9-11-ту добу прозора оболонка лопається (денудація) і ембріон "вилуплюється". У овець це буває на 7-8-му, свиней - на 6-ту добу розвитку ембріона. Незабаром після денудації зародка відбувається обширна проліферація тг>офобласта і помітний ріст в довжину бластоцисти. Такий зародок може тривалий час вільно переміщуватися в просвіті матки, змінюючи поступово свій розмір і форму. З 16-17-го дня у великої рогатої худоби розпочинається імплантація зародка.
Отже, найбільш придатним для вибивання періодом розвитку ембріона є Р-Я-н день, коли він ще захищений про-:орою оболонкою.
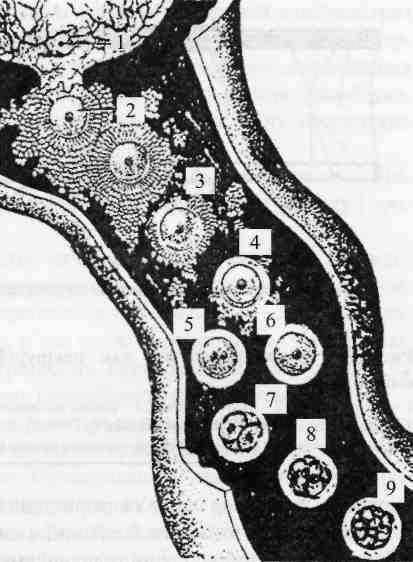
Рис. 43. Запліднення і ранні стадії розвитку
ембріона у яйцепроводі самки:
1 - фолікул, що овулював; 2 - яйцеклітина, оточена променевим вінцем; 3 - розсіювання променевого вінця; 4 - проникнення спермія крізь прозору оболонку; 5 - стадія двох пронуклеусів; 6 - зигота; 7—9 - стадії двох, чотирьох та восьми бластомерів.