

2.4.8. Отражение в характеристиках ээг уровня бодрствовапия
Между уровнем активации мозга и уровнем бодрстнонанни существует достаточно выраженная взаимосвязь: повышение уровня активации ЦНС обуславливает повышение уровня бодрствования. Графически эта взаимосвязь может быть проиллюстрирована схемой, предложенной В.Блоком (1970, Рис. 17). О повышении уровня активации может свидетельствовать, в частности, повышение частоты и снижение амплитуды и регулярности волн ЭЭГ (Рис. 18).
2.4.8.1. ЭЭГ-характеристики спокойного бодрствования
Характер ЭЭГ покоя определяется, прежде всего, индивидуальными особенностями структурно-функциональной организации ЦНС. Наряду с ними существенное значение имеет состояние зрительного анализатора, вследствие чего выделяю! состояние покоя с закрытыми и открытыми глазами.
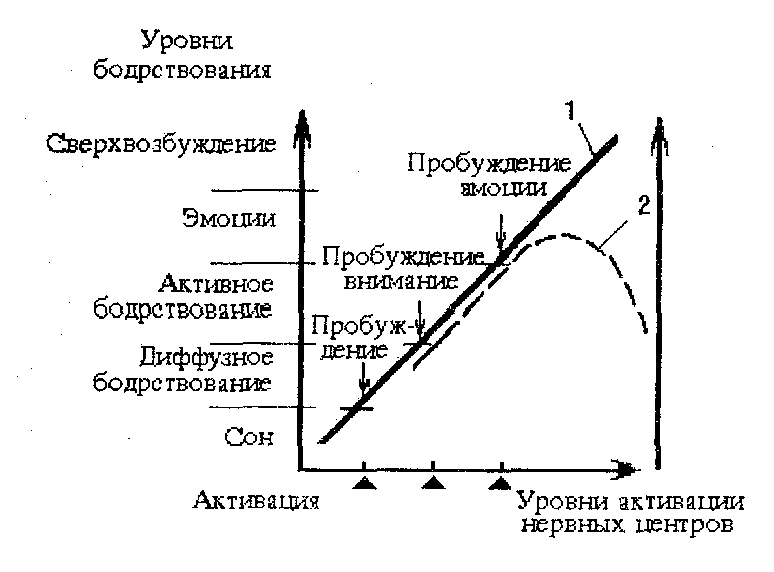
Рис. 17 . Зависимость между уровнем активации нервных центров, уровнем бодрствования (1) и эффективностью поведения (2).
Доминирующей в электрограммах спокойного бодрствования с закрытыми глазами более чем у 85% людей является альфа-активность, которая может быть как модулирована по амплитуде, так и не модулирована. Амплитуда и мощность альфа-частот, как правило, выше в ЭЭГ затылочных отведений, несколько ниже - теменных и центрачьных, и минимальна - лобно-височных, что формирует так называемый лобно-затылочный градиент.
Состояние покоя с открытыми глазами характеризуется, прежде всего, снижением амплитуды и мощности альфа-частот (Рис. 19). Его преимущественная выраженность в ЭЭГ затылочных отведений приводит к нарушению характерного для состояния покоя с закрытыми глазами градиента амплитуд; в бодрствовании с открытыми глазами амплитуда альфа-колебаний, как правило, выше в электрограммах теменных отведений. Лишь затемнение помещения и адаптация к условиям обследования способны восстановить профиль амплитуд, характерный для спокойного бодрствования с закрытыми глазами. Снижение амплитуды альфа-волн несколько нивелирует индивидуальные различия, отчетливо выраженные в покое с закрытыми глазами.
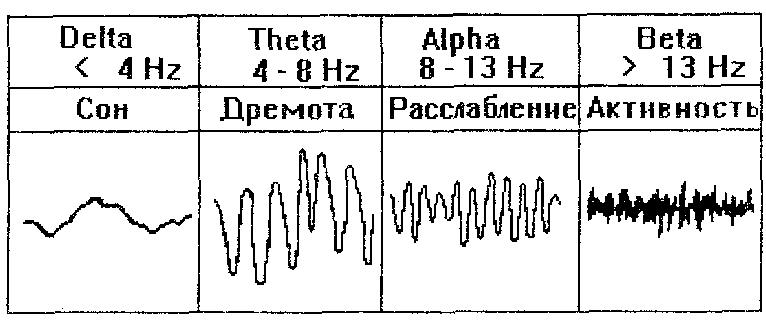
Рис. 18. Соотношение амплитудно-частотных характеристик мозговых волн и уровня бодрствования. Цит. по Дж.Хессету, 1981.
Как в покое с закрытыми, так и открытыми глазами мощность альфа-частот в ЭЭГ задних и центральных областей коры, как правило, значительно превосходит мощность всех других частотных диапазонов (Рис. 19). Различия менее выражены в ЭЭГ передних областей, в которых в покое с открытыми глазами преобладают медленные (лоб) и бета (висок) колебания.
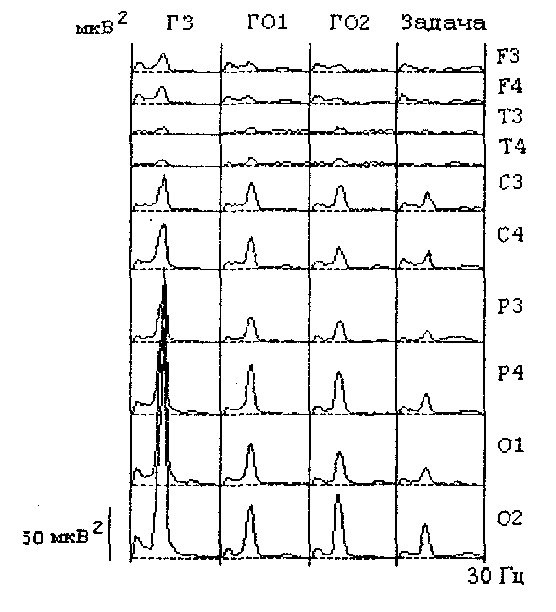
Рис. 19. Спектры мощности ЭЭГ, зарегистрированной у обследуемого в покое с закрытыми (ГЗ), открытыми (ГО1, ГО2) глазами и при решении задачи.
Средняя частота альфа-волн в ЭЭГ спокойного бодрствования повышается в направлении лоб-затылок (Табл. 6), а в покое с закрытыми глазами при сохранении отмеченной тенденции наблюдается увеличение доли более низких альфа-частот.
В состоянии спокойного бодрствования у всех практически здоровых лиц отчетливо проявляются описанные выше закономерности пространственно-временной организации биопотенциалов мозга. При этом между спокойным бодрствованием с альфа-активностью и с десинхронизированной ЭЭГ (в том числе, между состояниями покоя с открытыми и закрытыми глазами) имеются существенные различия. Они состоят, в том, что в покое с закрытыми глазами градиент КК в пределах каждого полушария нелинеен: имеет место существенное снижением их величины с увеличением расстояния в задних отделах, тогда как биопотенциалы фронтальных и центральных областей демонстрируют высокую положительную корреляцию (до 0,8 и выше). КК ЭЭГ наиболее удаленных лобно-затылочных отведений достигают величин -0,6. В покое с открытыми глазами градиент КК практически линеен и даже электрограммы лобно-затылочных отведений характеризируются синфазными отношениями.
Таблица 6.
Распределение мощности альфа-частот в ЭЭГ адаптированных обследуемых в состояниях покоя с открытыми и закрытыми глазами
Частоты (Гц) |
|
Области |
регистрации ЭЭГ |
|
|
|
|
Лоб |
Темя |
Затылок |
Лоб Темя Затылок |
||
|
Глаза открыты |
|
Глаза закрыты |
|||
8 |
11.5 |
3.6 |
3.6 |
7.0 |
4.3 |
2.8 |
9 |
15.4 |
5.5 |
5.5 |
15.4 |
7.7 |
8.2 |
10 |
36.1 |
50.5 |
19.2 |
43.8 |
23.1 |
26.9 |
11 |
23.2 |
36.2 |
30.6 |
19.2 |
42.1 |
30.6 |
12 |
8.5 |
15.4 |
25.0 |
11.6 |
20.7 |
27.8 |
13 |
5.3 |
8.7 |
16.1 |
- |
2.1 |
3.7 |
Фазовые спектры ЭЭГ (Рис. 20), зарегистрированной в покое как с открытыми, так и с закрытыми глазами, отчетливо делятся на две группы. Колебания в симметричных и теменно-затылочных отведениях имеют незначительные фазовые сдвиги на частотах, относящихся к одному ритму ЭЭГ. Колебания в лобно-теменных и лобно-затылочных парах отведений, напротив, имеют существенные фазовые сдвиги даже на близких частотах. Существенных различий между состояниями спокойного бодрствования с открытыми и закрытыми глазами не обнаруживается.
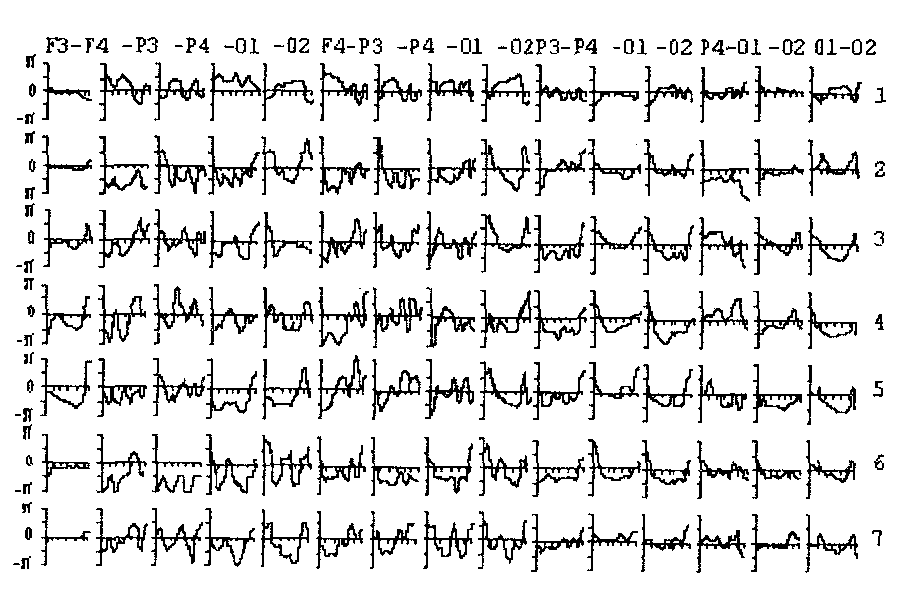
Рис. 20. Фазовые спектры ЭЭГ, зарегистрированной в покое с открытыми глазами у 7 человек.
Аналогичные группы выделяются и в спектрах когерентности (Рис.21). Когерентость колебаний симметричных и теменно-затылочных пар отведений в среднем выше 0.4-0.5 и достигает на низких и альфа-частотах значений порядка 0.7-0.9. Когерентность биопотенциалов лобно-теменных и лобно-затылочных отведений, как правило, существенно ниже. В целом когерентность выше в области низких и альфа-частот и ниже - в области частот бета-диапазона. При этом независимо от расстояния между электродами когерентность колебаний ЭЭГ в пределах правого полушария выше, чем левого.
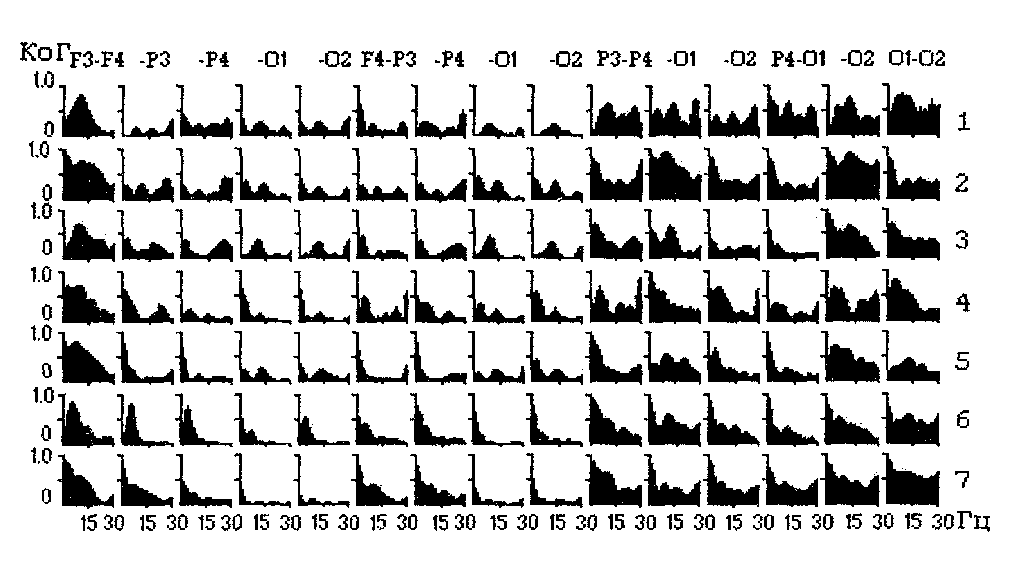
Рис. 21. Спектры когерентности ЭЭГ 7 человек в состоянии спокойного бодрствования с закрытыми глазами.
Внутриполушарная когерентность выше у женщин. Спектральных различий, связанных с ведущей рукой, не обнаружено, однако, леворукие имеют в среднем более высокий уровень когерентности справа на частотах 20-35 Гц. На этом основании делается вывод о том, что, независимо от ведущей руки, у женщин в целом более активировано левое полушарие, а у мужчин - правое. У леворуких, независимо от пола, более активировано правое полушарие, а у праворуких - левое. Половые различия в уровне когерентности могут определяться как большими размерами черепа у мужчин, так и различиями во взаимном расположении неокортикальных полей.
2.4.8.2. ЭЭГ-характеристики активного бодрствования 2.4.8.2.1. Изменение спектральных характеристик ЭЭГ
Активное бодрствование человека по своим внешним, поведенческим проявлениям иногда столь же бедно, как и спокойное. Зачастую единственным его индикатором является состояние ЦНС, в то время как вегетативные показатели остаются на уровне, характерном для состояния покоя. Последнее имеет место, в частности, в условиях интеллектуальной деятельности, которая может различаться по содержанию и уровню операционной напряженности.
В целом, изменения, наблюдаемые при переходе от состояния спокойного к активному бодрствованию, отражают активацию структур ЦНС. Их относительная независимость от побудивших их причин послужила основанием рассматривать активационные процессы, сопровождающие формирование активного бодрствования, как сугубо неспецифические, хотя, в действительности, уровень активации тех или иных структур определяется степенью их участия в реализуемой деятельности.
В настоящее время не представляется возможным сделать однозначного вывода о характере изменений медленных ЭЭГ-частот при переходе от состояния спокойного к активному бодрствованию. Показано, в частности, что в условиях зрительной стимуляции, сенсомоторной деятельности, при решении человеком арифметических, вербальных и наглядно-образных задач, а также при реализации других видов интеллектуальной деятельности мощность дельта-частот возрастает. Предполагается, что повышение их мощности в спектре ЭЭГ может свидетельствовать о усилении высокоселективных тормозных процессов в соответствующих структурах, устраняющих эндогенные и экзогенные факторы, не имеющие прямого отношения к реализуемой деятельности. Однако более детальные исследования показывают, что повышение суммарной мощности дельта-частот отмечается лишь в части случаев, более выражено в передних областях коры и отведениях правого полушария, и несколько менее - левого (Рис. 22).
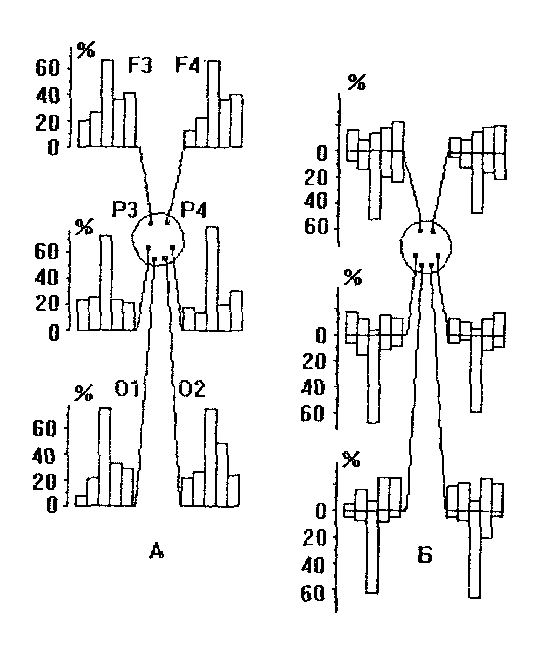
Рис. 22. Изменение мощности ЭЭГ-частот при переходе от состояния спокойного
бодрствования к активному. А - общие изменения, Б - в зависимости от их направления.
Регистрируемые в бодрствовании дельта-колебания следует отличать от дельта-ритма сна. Первые объединяют в так называемый Class 1, а второй - Class 2. Показано, в частности, что они различаются по уровню когерентности, которая во сне выше.
Дельта-активность, регистрируемая в бодрствовании, представлена единичными низкоамплитудными несинхронизированными колебаниями, преимущественно выраженными в отведениях правого полушария и лобных отделов. Глобальное усиление синхронизированных дельта-волн в ЭЭГ в бодрствовании связано с существенным снижением уровня активации мозга, а наличие в его электрической активности высокоамплитудного дельта-ритма трактуется в большинстве случаев как прямое указание на существование патологии стволового происхождения.
Аналогичные в целом изменения наблюдаются в области тета-частот. Зрительная засветка, слежение за движущимися объектами на экране радара или монитора, интеллектуальная деятельность (выполнение корректурной пробы, решение арифметических задач, чтение текста и т.д.), как правило, сопровождаются увеличением мощности тета-частот в спектре ЭЭГ. Поскольку наиболее отчетливое усиление тета-активности имеет место в электрограммах, регистрируемых от передних областей, сформулировано предположение о том, что тета-ритм связан с развитием состояния субъективного погружения в работу и интеллектуальным напряжением. Ряд авторов рассматривают его как показатель нервно-эмоционального напряжения, обозначая его ритмом напряжения.
T.Ishihara и N.Yoshii (1972) были описаны частотные (6-7 Гц) и амплитудные (30-60 мкВ) характеристики тета-волн, регистрируемых от средней линии в передних областях коры. Они получили название "Fm-theta". Критериями для идентификации Fm-theta являются ритмическая синусоидальная форма, сравнительно высокая амплитуда, продолжительность более 1 с и частота в интервале 4.5-7.5 Гц. Было показано, что Fm-theta имеет место только у части обследуемых, которые являются более экстравертированными, менее тревожными и нейротизированными личностями.
Увеличение числа низкоамплитудных тета-волн в ЭЭ1 отдельных областей связывают с развитием в них процессов "избирательного торможения", обеспечивающего селективное вытормаживание побочных иррелевантных связей и ассоциаций, связанных с произвольным вниманием, а также осуществлением автоматизированных процессов переработки информации Значительное усиление тета-частот свидетельствует о развитии и в коре тормозного состояния и отрицательно коррелирует со скоростью протекания мыслительных процессов.
Преимущественное усиление тета-активности в ЭЭГ передних областей коры позволило высказать предположение о том, что своим происхождение она обязана подкорковым структурам, в частности, гиппокампу. Известно, что при переходе от состояния покоя к активному бодрствованию имеет место экзальтация гиппокампального тета-ритма, который представлен регулярными высокоамплитудными волнами частотой 4-6 в 1 с. Последние могут электротонически регистрироваться в коре. Последнее объясняет отсутствие сколько-нибудь существенной корреляции регистрируемого в коре тета-ритма с особенностями реализуемой деятельности, в том числе, фиксацией или актуализацией следов памяти в ЦНС, механизмами квантования извлекаемых из памяти энграмм и т.д. В наиболее известных руководствах по электрофизиологии и клинической энцефалографии доминирование в ЭЭГ бодрствования периодического тета-ритма рассматривается как указание на наличие мозговой патологии различной (как правило, некорковой) природы.
Уже H.Berger показал, что в условиях активации амплитуда альфа-волн ЭЭГ снижается. Впоследствие десинхронизация ЭЭГ при переходе от спокойного к активному бодрствованию отмечалась практически всеми исследователями. Снижение амплитуды и мощности альфа-частот наблюдалось при предъявлении человеку как вербальных, так и невербальных нагрузок, в ответ на зрительную засветку, при выполнении пространственных задач, корректурных проб, зрительном слежении и т.д. Количественно наблюдаемые изменения наиболее выражены в тех областях коры, в которых в покое альфа-активность доминирует, т.е., теменных и затылочных. При автоматизации деятельности отмечается рост амплитуды альфа-колебаний до уровня, характерного для состояния покоя.
Поскольку десинхронизация альфа-ритма имеет место при предъявлении самых разных нагрузок, делается вывод о том, что она носит неспецифический характер. Однако это справедливо лишь отчасти. В изменении характеристик альфа-активности зачастую наблюдаются достаточно четко выраженные региональные различия, связанные с особенностями выполняемой деятельности. Так при выполнении вербальных заданий депрессия альфа-частот более выражена в отведениях левого полушария и передних отделов, а при активном использовании пространственных и квазипространственных видов синтеза - в правом полушарии и теменно-затылочных отделах. При этом отмечается отчетливая зависимость наблюдаемых изменений от субъективной сложности
деятельности: при ее увеличении имеет место более глубокая десинхронизация, причем преимущественно в отведениях левого полушария. Для легких заданий межполушарные различия не обнаружены.
Некоторые авторы выделяют, по крайней мере, 2 типа альфа-ритма: генерализованный, регистрируемый по всей коре, и локальный, свойственный лишь отдельным областям. Полагают, что первый играет роль общего регулятора, в то время как второй имеет отношение к локальным процессам, связанным с переработкой информации. На это указывает, в частности, то, что в условиях концентрации внимания депрессия альфа-ритма имеет место преимущественно в теменных и затылочных областях левого полушария, при выполнении моторных заданий она отмечается в центральных областях, а при чтении - в затылочных. Выраженность этих изменений зависит от индивидуальных особенностей ЭЭГ. Более или менее выраженное снижение мощности альфа-частот при различных нагрузках имеет место лишь в ЭЭГ лиц с выраженным альфа-ритмом (Рис. 23). У лиц с менее выраженным альфа-ритмом его мощность при предъявлении нагрузки в отдельных случаях может не изменяться, а при выполнении простой, хорошо известной деятельности, не требующей сколько-нибудь существенных усилий - даже возрастать. У лиц "без альфа-ритма" (т.е., с десинхронизированным типом ЭЭГ) в активном бодрствовании мощность альфа-частот либо не изменяется по сравнению с покоем, либо даже несколько возрастает. Последнее также имеет место в привычных условиях деятельности, при выполнении субъективно простых заданий и автоматизированных навыков.
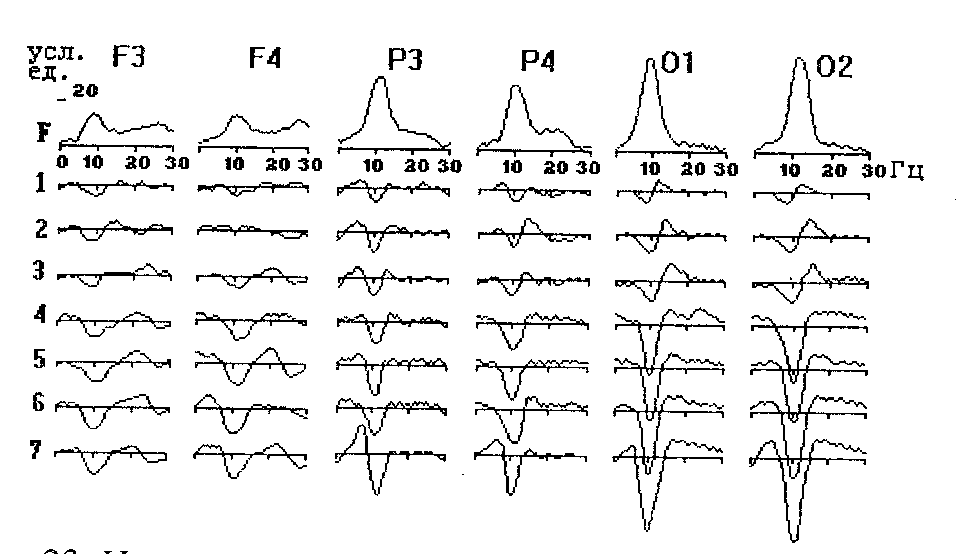
Рис. 23. Изменения спектральной мощности ЭЭГ-частот при решении обследуемым с выраженным альфа-ритмом мыслительных задач.
F-спектральная мощность ЭЭГ-частот в покое, 1 - 7 - ее изменения при решении задач повышающейся сложности.
При переходе от спокойного к активному бодрствованию в целом ряде случаев отмечаются изменения частотных характеристик альфа-ритма. Как правило, они связаны с повышением его частоты, которое имеет место, в частности, в условиях решения математических, пространственных и других типов задач. Более существенные изменения отмечаются у лиц с преобладанием низкочастотной альфа-активностьи и в условиях субъективно более сложной деятельности. Смещение моды в область боле высоких значений связано с более выраженным подавлением низких альфа-частот. При этом гистограммы распределения мощности в диапазоне альфа-частот могут приобретать би- и даже тримодальный характер, что рассматривается как свидетельство параллельного функционирования нескольких альфа-генераторов, различающихся по своим частотным характеристикам.
Как правило, при повышении уровня активации мозга в ЭЭГ отмечается рост мощности бета-частот. Это показано при развитии ориентировочной реакции, в условиях решения различных задач, при чтении текстов и т.д. Вместе с тем, иногда приводятся сведения о том, что в аналогичных условиях мощность бета-частот может не только не возрастать, но даже снижаться. Это показано, в частности, в прецентральпыч областях при выполнении движений и в париетальных при чтении текстов. В связи с этим бета-десинхронизация иногда рассматривается как свидетельство повышения уровня активации мозга. В значительной степени указанные противоречия определяются областью частот ритма бета-1, в которой наряду с альфа- и может иметь место бета-десинхронизация. Мощность бета-2-частот при этом, как правило, увеличивается, причем это более выражено в ЭЭГ задних отделов неокортекса.
В последние годы внимание привлекает диапазон гамма-частот (В.Н.Кирой, О.Г.Чораян, 2000), которые иногда рассматриваются как высокие бета-частоты. Появление осцилляторной активности с частотой 40 Гц и более показано, в частности, в ЭЭГ сенсомоторных областей коры при подготовке к выполнению тонких движений, в условиях сенсорной стимуляции, при воспроизведении энграмм из памяти, ассоциативном обучении и т.д. Учитывая наличие наряду со спонтанными вызванных гамма-осцилляций, топографию их возникновения и ряд других сведений, полагают, что они непосредственно связаны с информационными процессами, протекающими в нейронных структурах мозга.