

Материнская часть плаценты образуется за счет основной отпадающей оболочки (части) миометрия
( слизистой оболочки матки). Параллельно происходят изменения в хорионе. Эти изменения обеспечивают выполнение 2-х задач: снабжение растущего зародыша веществами, необходимыми для его развития, предохранение матки от чрезмерного проникновения хориона.
Комплекс этих изменений приводит к превращению слизистой оболочки матки в децидуальную ткань, наличие которой является главным критерием для деления плацентарных животных на группу адецидуатов, слизистая матки которых не изменяется, и децидуатов, эндометрий которых подвержен децидуальным изменениям.
В децидуальную ткань превращается соединительная ткань стромы слизистой оболочки матки. В соединительной ткани в большом количестве образуются децидуальные клетки. В их цитоплазме появляются многочисленные зерна гликогена и липиды. Значительно увеличивается гидратация слизистой оболочки, становятся менее различимы ретикулярные и коллагеновые волокна, значительно увеличивается просвет сосудов. Эти изменения приводят к тому, что слизистая оболочка матки становится толстой, сочной и рыхлой.
Физиологическое значение децидуальной реакции заключается в том, что для плода она обеспечивает питание и защиту. Трофическое значение ее проявляется, главным образом, в ранней гистотрофной фазе питания плодного яйца. Зигота перед имплантацией питается, в основном, продуктами слизистой и, прежде всего, секретом децидуальных желез. Далее, главным источником трофного питания являются продукты цитолиза децидуальных клеток.
Защитная функция децидуальной ткани проявляется в ее исключительной фагоцитарной активности. Благодаря этой способности она связывает и разрушает токсины, бактерии, проникающие из организма матери к плоду.
Для матери защитная роль децидуальной системы состоит в торможении ею внедрения трофобласта. Это осуществляется благодаря наличию в Децидуальной ткани протеолитических ферментов, которые во второй половине беременности сохраняются только в более глубоких слоях слизистой.
К защитным факторам децидуальной ткани относится наличие в ней в значительных количествах фибринобластических (способствующих свертыванию крови) п. фибринолитических (усиливающих процесс расплавления тромбов) факторов. На более^оздних этапах беременности эта ткань замещается фибрингиалиноподоб- ной массой.
Фибриноид образуется в месте непосредственного контакта синцитотрофобласта и децидуальных клеток.
Плацента выполняет ряд функций: трофическую, дыхательную, выделительную, барьерную, эндокринную.
1. Трофическая: из крови матери в кровь плода поступают различные питательные вещества (аминокислоты, глюкоза, нуклеотиды, витамины), необходимые для .нормального развития зародыша.
2. Дыхательная: кровь плода богата углекислотой, которая легко и быстро диффундирует через плаценту. Кислород диффундирует не так легко и для обеспечения плода достаточным количеством кислорода напряжение его в тканях матери должно быть значительно выше, чем по другую сторону плацентарного барьера. Установлению этого градиента способствует также отличие гемоглобина плода от гемоглобина взрослого организма по сродству к кислороду. При одном и том же напряжении кислорода гемоглобин плода способен связывать больше кислорода, чем гемоглобин матери.
3. Выделительная функция плаценты заключается в выведении продуктов распада (мочевина, аммиак и др.) из крови плода в кровь матери.
Трофическая, дыхательная и выделительная функции плаценты обеспечиваются с помощью организма матери. Поэтому беременная женщина должна соблюдать нормальный режим, не курить, не принимать алкоголь, который беспрепятственно проходит через плаценту и разрушает лервную,- систему плода. Будущая мать должна быть на воздухе, следить за работой своего выделительного аппарата.
4. Барьерная функция. Плацента является барьером, через который не проходят из крови матери в кровь плода многие вещества, которые могут нарушить нормальное течение эмбриогенеза. Ни один микроб, кроме бледной спирохеты, вызывающей сифилис, не проникает через плаценту. Плацента защшвдет организм матери от антигенов плода, препятствует развитию иммунобиологического конфликта.
5. Эндокринная: плацента является временной эндокринной железой, которая обеспечивает нормальное течение беременности. Один из первых гормонов, который был выделен из плаценты — это гонадотропный гормон. Максимум его секреции достигается к 8—9 неделе, а затем содержание его снижается. Вырабатывают его клетки Лангганса. Планцета вырабатывает АКТГ (при беременности улучшается состояние больных Адисоновой болезнью, заболеваниями соединительной ткани — коллагенозами), тиреот- ропный гормон, меланОфорный, лютеотропный, эстрогены, прогестерон, андрогены, кортикоиды. Вьфабатываемый плацентой релаксан вызывает расслабление лонного сочленения при беременности. В плаценте вырабатывается гистамин, под его влиянием расширяются капилляры, а гладкая мускулатура — сокращается. Гистамины разрушаются гистаминазами, которые вырабатываются плацентой в большом количестве. Когда плацента стареет, количество гистаминазы уменьшается и гистамины приводят мышцы матки к родовой деятельности. Образующийся в плаценте ацетилхолин оказывает то же действие, что и гистамин.
Таким образом, плацента обеспечивает нормальное развитие
зародыша. После рождения плода материнская и дочерняя части плаценты отторгаются из матки. Это получило название рождение последа.
ТЕОРЕТИЧЕСКИЕ ОСНОВЫ ПОНИМАНИЯ ЭМБРИОГЕНЕЗА
В предыдущих разделах . были описаны основные этапы эмбриогенеза. Как известно, филогенез дает историю вида, онтогенез 45- историю индивида. В свою очередь онтогенез, описывающий историю индивида от момента зарождения до наступления физиологической смерти, может быть подразделен на эмбриональный и постэмбриональный период. Эмбриология как наука изучает эмбриональный период онтогенеза. У плацентарных, к которым относится и человек, эмбриональный период захватывает время от момента оплодотворения до рождения, т. е. до момента изгнания плода из матки.
Рассматривая многочисленные работы, выполненные по эмбриологии в течение нескольких столетий, можно юс подразделить на описательные или феноменологические и на экспериментальные, стремящиеся вскрыть причины и механизмы, обусловливающие сложные и не до донца выясненные процессы эмбриогенеза.
Описание эмбриогенеза позволяет выделить в нем стадии, процессы, элементы и элементарные акты. Под стадией следует понимать этап развития зародыша, коренным образом отличающийся от предыдущего в силу появления новых свойств и структур. Исходя из этого определения можно выделить следующие стадии эмбриогенеза: зигота, бластула, гаструла, нейрула, тканевое и органное строение.
.Переход от одной стадии к следующей обеспечивается с помощью определенных эмбриональных процессов.
Так, появлению стадии зиготы, предшествует процесс прогенеза, сущностью которого является образование и созревание половых клеток, их перенос навстречу друг другу с последующим слиянием. Оплодотворение является процессом, приводящим к появлению зиготы. Биологическая сущность оплодотворения состоит в том, что две половые клетки с гаплоидным набором хромосом, не способные раздельно к прогрессивному развитию и обреченные на быструю гибель, при встрече сливаются и образуют диплоидную одноклеточную стадию организма — стадию зиготы с обширной и реализуемой программой прогрессивного развития.
Процесс, с помощью которого зигота переходит в следующую стадию развития — бластулу, получил название дробления. Дробление совершается, используя известный механизм митотическо- го деления, но с рядом важных особенностей. Во-первых, при дроблении интерфаза резко укорочена. Она представлена, по сути, периодом, в котором протекает репликация. Во-вторых, при дроб-
момента оплодотворения до рождения на свет (у живородящих организмов) или до вылупливания из яйцевых оболочек (у яйцекладущих животных)». (А. Г. Кнорре, 1914—1982).
Предметом изучения эмбриологии является также прогенез — предзародышевый период (формирование половых клеток) и по- слезародышевый- (постэмбриональный) или послеродовой (постна- тальный) периоды индивидуального развития. У млекопитающих: некоторые системы органов (например, половая, эндокринная) приобретают дефинитивные, т. е. окончательные, свойственные взрослому состоянию, структуры и функции спустя тот или иной промежуток времени после рождения.
Эмбриология исследует как общие закономерности индивидуального развития, проявляющиеся в развитии всех'многоклеточных животных организмов, так и частные особенности процессов индивидуального развития, характерные для представителей отдельных типов, классов или видов животных.
В связи с этим различают общую эмбриологию, рассматривающую наиболее общие вопросы и наиболее широкие закономерности индивидуального развития, и частную эмбриологию отдельных групп животных.
Значение эмбриологии заключается прежде всего в формировании общих биологических представлений на- материалистической основе. Велико значение эмбриологии как теоретической основы педиатрии, акушерства и гинекологии, тератологии.
МЕТОДЫ ИССЛЕДОВАНИЯ
Для изучения эмбрионального развития животных пользуются различными методами.
Простейший и наиболее древний из них — наблюдение и описание развития зародыша у тех или иных животных или человека. На основе этого метода возникла описательная эмбриология.
Значительно позже в эмбриологических исследованиях стал применяться сравнительный метод, создавший сравнительную эмбриологию. Ее возникновение было обусловлено накоплением фактов о развитии различных видов животных. Применение сравнительного метода открыло возможности для понимания общих закономерностей развития. С помощью сравнительного метода был открыт закон зародышевого сходства К. М. Бэром (1792—1876).
Описательная и сравнительная эмбриология стали фундаментом для развития эволюционной эмбриологии, создателями которой явились А. О. Ковалевский (1840—1901), И. И. Мечников (1845— 1916). Развитие эволюционной эмбриологии привело к открытию биогенетического закона Э. Геккелем (1834—1919), Ф. Мюллером (1821—1897), А. Н, Северцевым (1861—1936).
С развитием и усложнением экспериментальной техники воз
никло новое направление исследований — экспериментальная эмбриология. Основателями ее явились В. Ру (1850—1924), Г. Дриш (1867—1941) и Г. Шпеман (1869—1941). В основе экспериментальной эмбриологии лежит изучение последствий удаления или трансплантация различных частей зародыша, а также воздействия на зародыш или его части разных экзогенных факторов.
Современная эмбриология пользуется всеми видами световой, электронной и сканирующей микроскопии, методами прижизненного наблюдения эмбриогенеза при помощи меток, наносимых на зародыш с помощью изотопов или красителей, методами гистохимии, биохимии, радиологии, иммунологии, а также статистики.
ТЕОРИИ ЭМБРИОЛОГИИ
. Как и любая другая наука, эмбриология на каждом этапе развития была вооружена собственной теорией.
С именем Гиппократа (460—377 год до н. э.) и Аристотеля (384—322 год до н. э.) связывают не только определенные знания в области развития организмов, но и создание первых научных эмбриологических представлений.
Согласно теории Гиппократа, плод образуется через смешение мужского и женского семени. Он считал, что все члены организма 1возникают независимо друг от друга. Ни один из них не возникает раньше или позже другого.
Эта идея вновь возродилась и стала господствовать в течение XVII—XVIII вв. Она получила название теории преформизма. Согласно этой теории каждый зародыш с самого начала является уже вполне сформированным, имеющим все части организма, которому остается только расти. Наиболее крайние и наивные преформисты представляли себе, что каждый организм, в том числе и человеческий, заключает в себе громадное количество поколений зародышей, вложенных друг в друга в теле прародителей со времени сотворения мира.
Между представителями двух направлений преформистов велись горячие споры. Анималькулисты или сперматики считали, что зародыш преформирован в сперматозоиде. Овисты думали, что зародыши в миниатюрном виде находятся не в мужских половых клетках, а в яйце.
Сторонниками префоризма были многие выдающиеся ученые XVII и XVIII века: А. Левенгук (1632—1723), Я. Сваммердам (1637—1680), М. Мальпиги (1628—1694), А. Галлер (1708—1777), Ш. Бонне (1720—1793).
Аристотель впервые сформировал теорию, эпигенеза. Согласно Аристотелю, части зародыша не заложены в невидимо малых размерах в «семенах вещей» и не возникают все одновременно, а растут и развиваются в определенной последовательности. Это
лении имеет место в интерфазе только синтез и-РНК, остальные виды РНК и синтез белка еще не происходят. Отсюда, клетка не способна к росту и поэтому при каждом новом этапе дробления дочерние клетки-бластомеры получают размеры вдвое меньше,, чем материнские.
Если вспомнить положение биогенетического закона и приложить его к дроблению, то этот процесс онтогенеза как бы повторяет в ускоренном виде переход от одноклеточных к многоклеточным формам-, который имел место в филогенезе. '
Следующий процесс, эмбриогенеза — это гаструляция, с помощью которой зародыш из стадии бластулы переходит в стадию гаструлы. В основе этого процесса лежит перемещение клеточных масс, приводящее вначале к появлению двух, а затем и трех зародышевых листков: эктодермы, энтодермы и мезодермы: Причины и механизмы этого перемещения клеток выяснены недостаточно.
. Продолжающееся перемещение клеток тела зародыша — процесс нейруляции, приводит к образованию органов спинного комплекса: нервной трубки, хорды, сомитов, тем самым приводит зародыш на стадию нейрулы. На этой стадий развития зародышевые зачатки заняли «исходные рубежи» и готовы к следующему: процессу — гистогенезу, т. е. к образованию тканей. Параллельно- с^образованием тканей их взаимодействие приводит к появлению новых структур — возникают органы. Процесс образования органов, получил название - органогенеза. Очень часто оба эти процесса объединяют под общим термином морфогенеза.
Процессы морфогенеза продолжаются на протяжении всей жизни человека или животного. В настоящем разделе мы рассмотрим эти процессы в эмбриональном периоде. .
Все процессы эмбриогенеза можно условно разделить на элементы. К ним относятся: размножение клеток, рост клеток, образование некоторыми из, них межклеточного промежуточного вещества, перемещение клеток, детерминация и дифференцировка клеток, взаимодействие и гибель клеток.
Размножение клеток как элемент эмбриогенеза представлен дроблением, митозом и мейозом. Основная масса клеток возникает в результате митотического деления. Роль амитоза, как самостоятельного вида деления клеток, в. условиях нормального развития организма в настоящее время пересматривается. Мейоз свойственен только половым клеткам в период их созревания. У большинства видов животных, в том числе и у человека; он протекает в постэмбрйональном периоде.
Размножение клеток и последующий их рост, начиная со стадии гаструлы, генетически запрограммированные процессы, регулируемые действием биологически активных веществ. Рост клеток объясняют преобладанием процессов ассимиляции над
процессами диссимиляции. Уравнение интенсивности этйХ^процес- «сов приводит к остановке роста клеток.
Одну из загадок эмбриогенеза представляют способы и механизмы перемещения клеток. Можно выделить перемещение клеток пластом, как это имеет место при гаструляции и нейруляции; перемещение группами или отдельными клетками, как это имеет место при миграции первичных половых клеток, нейробластов в -состав ганглиев, миобластов из миотомов в места локализации -будущих скелетных мышц. Механизм перемещения отдельных клеток объясняют * с позиций амебоидного и ундулирующего движения. Направление движения определяется хемотаксисом, а •остановка'—механизмом контактного торможения,•• в основе которого лежит прекращение движения при встрече однотипных клеток. ,
Прежде чем перейти к рассмотрению механизмов детерминации :и дифференцировки, дадим определение понятию «элементарный акт». В основе всех явлений жизни, как особой формы движения матери, а, Следовательно, процессов, протекающих в клетках, тканях и органах на этапах эмбриогенеза, лежат элементарные -акты морфогенеза.
Под элементарным актом мы понимаем цикл молекулярно-фер- ментативных. процессов, приводящих к образованию макромолекул или сборке надмолекулярных субклеточных структур.
К элементарным актам морфогенеза следует отнести процессы репликации, приводящие к., синтезу молекул ДНК, процессы транскрипции—ксинтезу всех.видов молекул РНК (и-РНК, р-РНК, т-РНК, рег.-РНК), процессы трансляции, приводящие к синтезу молекул белка, а также' процессы самосборки, приводящие к образованию различных мембран и органелл.
При эмбриогенезе появление и становление элементарных актов морфогенеза происходит в определенной последовательности. В зиготе возникшей из изолецитальной яйцеклетки, еще не представлены макромолекулярные синтезы. В период дробления у зародышей этого типа появляются процессы репликации и начинается синтез и-РНК. На стадии бластулы к этим элементарным актам присоединяется синтез т-РНК, на стадии гаструлы — р-РНК наконец, на стадии нейрулы в клетках зародыша представлены все виды макромолекулярных'синтезов и сборки надмолекулярных структур. Тем самым, на стадии нейрулы получили развитие все элементарные акты, обеспечивающие дальнейшее эмбриональное развитие.
Стадия нейрулы является стартовой площадкой для; дальнейших процессов гистогенеза, объединяемых общим термином «морфогенез». Под морфогенезом следует понимать цепь последовательно развивающихся процессов, приводящих к становлению определенных структур и функций.
Исследуя начальные, промежуточные и конечные этапы морфогенеза, нетрудно заметить, что различные по структуре и функциям структуры возникают часто из одинаковых зачатков и отличия между ними нарастают постепенно. Так, нервная ткань и развивающийся из нее спинной и головной мозг возникают из общего с кожей зачатка — эктодермы. Жидкая кровь, костная ткань и жировая ткань у зародыша имеют общего родоначальника в виде мезенхимы. Вместе с тем, клетки крови резко отличаются как от клеток костной, так и жировой ткани по своим размерам, форме, функции, биохимизму, что указывает, что активность генов, определяющих все эти отличия, различна у клеток этих тканей.
Процесс, который при онтогенезе приводит к развитию специализированных по структуре и функции клеток, получил название дифференцировки. Как объяснить, что одинаковые по своему происхождению, структуре и функции клетки мезенхимы выбирают различные направления развития, т. е. дифференцируются по-разному? Для объяснения этого очень важного положения введено понятие о детерминации.
Под детерминацией следует понимать стойкое и передающееся по наследству изменение состояния регуляторных структур клетки, определяющее направление дифференцировки.
Итак, под действием определенных факторов в клетке происходит детерминация, т. е. клетки получают программу развития. Затем наступает дйфференцировка, с помощью которой реализуется программа детерминации.
Понимание механизмов развития, что получило название «механики развития», создавалось работниками экспериментаторов-эмбриологов на протяжении более 100 лет и существенно продвинулось благодаря успехам молекулярной биологии, молекулярной генетики и электронной микроскопии.
Как известно, из зиготы путем дробления возникают бластомеры, из которых в свою очередь развиваются различные клетки зародыша. Решим, во-первых, вопрос, какова роль ядра клеток в процессе дифференцировки клеток. С этой целью рассмотрим результаты опытов Бриггса и Кинга.
Излюбленным объектом для решения вопросов механики развития являются зародыши амфибий, в частности, головастики. Это объясняется доступностью объекта, неприхотливостью, высокой скоростью эмбрионального развития.
Если удалить ядро из зиготы головастика с помощью специальной иглы, т. е. энуклеировать зиготу, то последняя совершит несколько дроблений, но возникшие энуклеированные бластомеры вскоре погибнут и дальнейшее развитие прекратится. Вывод ясен — ядро необходимо для процессов эмбриогенеза.
Эксперимент несколько изменяется: в энуклеированные зигбты будущих головастиков трансплантируются ядра, выделенные из
бластомеров или из клеток зародыша, находящихся на стадии бластулы, ранней гаструлы, поздней гаструлы и нейрулы. Затем ведется наблюдение за судьбой зародыша. В подавляющем проценте случаев энуклеированные зиготы, которым были трансплантированы ядра из клеток бластулы и ранней гаструлы проходят все этапы развития и превращаются в нормальных головастиков. Эти результаты позволяют сделать выводы, что ядра клеток зародыша, находящегося на этих стадиях, по своим свойствам не отличаются от ядер зиготы, т. е. они эквипотентны, а поскольку они обеспечивают различные дифференцировки в процессе эмбриогенеза, то ядра клеток бластулы и ранней гаструлы подобно ядрам зиготы полипотентны.
При трансплантации ядер клеток зародышей, находившихся на стадиях поздней гаструлы и нейрулы, развитие головастиков прекращается из-за гибели на стадии ранней гаструлы. Следовательно, при эмбриогенезе/ ядра клеток зародышей уже на стадии поздней гаструлы претерпевают изменения и теряют способность обеспечивать развитие зиготы до взрослой формы. На стадии поздней гаструлы ядра клеток головастиков уже не экви- и не полипотентны. На основании этих опытов можно сделать вывод, что в процессе эмбриогенеза изменяются свойства ядер клегок: уменьшается их способность обеспечивать развитие клеток в различных направлениях, т. е. они специализируются или иначе дифференцируются.
В отличие от Бриггса и Кинга, Гердон в: более поздних работах обнаружил, что в 1% случаев, т. е. очень , редко, ядра клеток эпителия кишечника головастика при трансплантации в эну- клеированную зиготу могут вызывать развитие нормального зародыша. Эти результаты требовали объяснения. В настоящее время можно дать, по-видимому, два объяснения результатам Гердона. Первое: среди эпителиоцитов кишечника есть вдетки, которые могут подвергнуться дедифференцировке, стать снова полипотент- ными и обеспечить развитие зародыша. Второе: в процессе развития не все клетки дифференцируются, очень небольшая часть из них сохраняет экви- и полипотентность, составляя камбий различных тканей. Это можно объяснить небольшим процентом успешных трансплантаций у Гердона. Тем самым опыты Гердона не отменили основных выводов, сделанных Бриггсом и Кингом, но поставили вопрос о возможности дедифференцировки ядер в процессе эмбрионального развития.
Общий вывод, который можно сделать на основании всех Зтих экспериментов, состоит в том, что изменения в ядрах клеток имеют большое значение в механизме эмбриогенеза.
Возникает вопрос: какова роль цитоплазмы при этом? Для выяснения этого вопроса обратимся к результатам некоторых экспериментов. Хорошо известно, что образование всех видов РНК>
исключая митохоидриальные, протекает в ядрах клеток. Этот процесс транскрипции может быть легко обнаружен современными методами авторадиографии и биохимии. Как было показано выше,, становление синтеза различного вида молекул РНК происходит постепенно, по мере прохождения по стадиям развития. Этот процесс протекает только в ядре клеток.
Для решения вопроса в какой степени синтез молекул РНК зависит от зрелости цитоплазмы были поставлены следующие опыты. В энуклеированную зиготу трансплантировали ядра клеток,, выделенные у зародыша головастиков, находящихся на стадии бластулы, гаструлы и нейрулы. В этих ядрах, как известно, уже имеет место синтез различных комплексов молекул РНК. Однако, при взаимодействии этих ядер клеток с цитоплазмой зиготы они временно прекращали процессы транскрипции. Следовательно, реализации транскрипций на этапах эмбриогенеза б клетках зародыша требует определенной зрелости цитоплазмы, способной ингибиро- вать эти процессы.
Все сказанное выше позволяет сделать вывод по этому разделу: не только ядро определяет развитие элементарных актов в цитоплазме зародыша, но и цитоплазма в свою очередь регулирует ход элементарных актов в ядре клеток, т. е. взаимодействием ядра и цитоплазмы можно объяснить сложные процессы развития клеток, их дифференцировку на этапах эмбриогенеза.
Молекулярно-генетические механизмы дифференцировки клеток в последнее время подвергаются интенсивному изучению. У прокариот объяснение развития дифференциальной активности генов можно получить с позиций гипотезы Жакоба и Моно. Сложнее дело обстоит у эукариот. У бактерий, имеющих одну кольцевую хромосому, репрессия и депрессия генов получает объяснение с позиций взаимодействия оперона с геном-регулятором. У эукариотов в ядре находится несколько хромосом, в которых располагается громадная по объему генетическая информация. Существуют механизмы взаимодействия между отдельными порциями генетической информации, локализованными в различных хромосомах.
Приблизиться к пониманию хотя бы в обшюс чертах к механизмам взаимодействия между различными отделами генома клетки в процессе дифференцировок клеток эукариот при эмбриогенезе, помогает схема-гипотеза, высказанная Бриттеном^ Дэвидсоном.
В соответствии с понятиями Жакоба и Моно, Бриттен и Дэвидсон подразделяют гены на регуляторные и продюссерные (структурные). В свою очередь, регуляторные гены подразделяют на сензорные, интегрирующие и рецепторные. Сензорные гены взаимодействуют с регуляторными факторами-сигналами (гормоны,, биологически активные вещества). Сензорным генам подчинены гены-интеграторы, которые под воздействием сензорных генов определяют выработку активаторной РНК. Последняя может дейст
вовать на рецепторные гены, расположенные в различных «ролф- сомах и тем самым генерировать и усиливать сигнал. Рецепторные гены под действием активированной РНК включают гены- продюссеры, которые подобно структурным генам определяют синтез различных видов РНК, а, следовательно, и синтез определенных видов белка, которыми на конечном этапе и определяется ход и направление дифференцировки клеток.
Характер этих взаимодействий молекулярно-генетических механизмов клеток при их дифференцировке представлен на схеме 1.
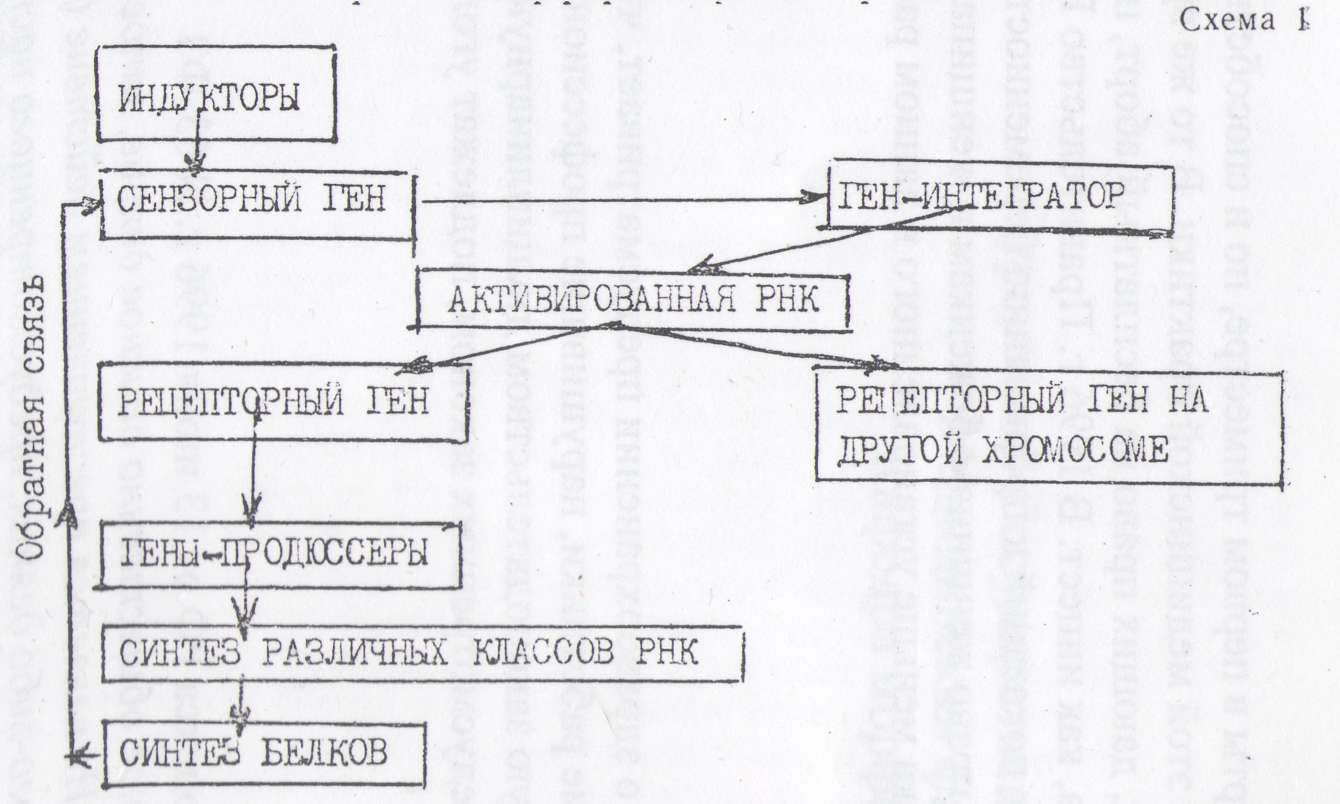
Некоторые из вновь синтезированных белков, или продукты,, возникающие под их действием, обеспечивают обратную связь,, воздействуя на сензорные гены.
Процесс развития протекает постепенно по схеме причинно- следственных взаимоотношений, следствие причины 1 может выступать в роли причины 2 и определять появление следствия 2 и т. д. Этот процесс развертывается во времени. Время, в свою очередь, определяется последовательностью генетически детерминированных процессов, протекающих в какой-то структуре.
Процесс развития зародышей не автономный, а зависит от действия ряда причин. Неоплодотворенные половые клетки у высших, организмов не способны обеспечить развитие зародышей. Толчком или причиной, обуславливающей развитие, является оплодотворение, которым запускается весь эмбриогенез и, в первую очередь,, дробление.
Развитие элементарных молекулярных актов на этапах раннего эмбриогенеза объясняется взаимодействием генетических структур ядра и органелл цитоплазмы. На стадии нейрулы основные
эмбриональные зачатки уже детерминированы, чем и подготовлен следующий этап в развитии зародыша—этап гистогенеза, а затем и органогенеза.
В развитии этих этапов важнейшая роль принадлежит взаимодействию клеток. Характер этих взаимодействий для высших животных был вскрыт в опытах Шлепана и его учеников. При этом было показано, что удаление зачатка хорды у зародышей амфибий блокирует образование из эктодермы нервной пластинки и превращение последней в нервную трубку. Пересадка хордального тяжа на вентральную сторону зародыша приводила к развитию нервной трубки на передней стороне тела головастика. Если зародышу головастика на стадии гаструлы трансплантировать зачаток хорды, взятый из другого зародыша, то отмечалось развитие двух нервных трубок на дорзальной и вентральной сторонах тела. На основании этих опытов был сделан вывод, что хорда вызывает развитие нервной трубки.
Этот процесс получил название эмбриональной индукции, а метаболиты хорды, ответственные за процесс, получили название индукторов.
Вторым классическим примером взаимодействий в процессах эмбрионального органогенеза может быть индукция развития хрусталика под действием глазного бокала. Индуктор является метаболитом и переносится из клеток индуцирующей в клетки индуцируемой структуры. Разделение этих структур с помощью сплошных, непроницаемых перегородок блокирует эффект индуктора, разделение с помощью решетчатых перегородок не препятствует развитию эффекта. Индуктор, достигнув клеток-мишеней, выступает в роли фактора, действующего на генетические структуры— сензорные гены и вызывает стойкое и наследуемое изменение активности генов — развитие эпигеномной наследственности. Таков, по-видимому, механизм детерминации, определяющий в дальнейшем характер последующих дифференцировок.
Коррекция процессов дифференцировки и перемещение клеток, приводящих к развитию тканевых и органных структур, получают объяснение с позиций принципа проверки обстановки клетками, предложенного Боннером. Согласно этому принципу, дифференци- ровка клеток идет ступенчато и клетки проверяют окружающую обстановку, сверяя ее с генетической программой своего развития. Если такая сверка совпадает с программной, то наступает следующий запрограммированный этап развития. Реализация принципа проверки обстановки допустима в том случае, если считать, что принцип дополняется принципом сигнализации. Согласно этому принципу каждый этап дифференцировки сопровождается выработкой клетками веществ сигналов, которые являются индукторами для последующих дифференцировок в клетках, проверяющих обстановку.
Мы попытались представить положения этой чрезвычайно слож- ' ной и бурно развивающейся отрасли знания о теоретических основах процессов эмбриогенеза. Каждый день приносит новые сведения, дополняющие наши знания, ставящие новые проблемы перед учеными.
Как было показано выше, теоретические проблемы эмбриогенеза в разное время получали объяснение с позиций преформизма или эпигенеза. Основатель преформизма Бонне считал, что в женской яйцеклетке находятся зародыши в готовом виде, в яйцеклетках этих зародышей находятся еще. меньшие по размерам зародыши следующего поколения. По его подсчетам в яичнике Евы содержалось 27 000 000 зародышей всех последующих поколений людей. Абсурдность подобных построений сохраняло, однако, свое значение в течение длительных этапов и эти отголоски преформизма пытаются найти свое убежище в концепциях неопреформизма.
С позиций неопреформизма в яйцеклетке заложен не готовый зародыш, а тот молекулярный материал, из которого будет идти развитие будущего организма. Неопреформизм продолжает отрицать возникновение новых структур в процессе развития.
Положение теории эпигенеза в первичном ее представлении также требует коррекции. Структуры цитоплазмы яйцеклетки не гомогенны и не полипотентны. Из определенных зон яйцеклетки, как правило, возникают определенные структуры зародыша. Следовательно, цитоплазма яйцеклетки в какой-то степени преформиро- вана в виде развития структур из подготовленного в процессе га- метогенеза материала. Иначе не имел бы смысл сложный подготовленный этап созревания женской половой клетки.
Обе длительное время конкурирующие теории могут быть ис- поЛьзованы при создании синтетической теории эмбриогенеза. Такой теорией должна стать генетическая теория развития.
Половые клетки не имеют готовых структур будущего зародыша, но они содержат набор генов — генетическую программу для данного вида животных, включая особенности для каждого индивидуума. С этих позиций структуры половых клеток и зиготы не гомогенны и не экви- и полипотентны. Вместе с тем, в процесса эмбриогенеза возникают структуры от молекулярного до органного уровней, которых нет в половых клетках. Это подтверждает некоторые положения эпигенеза. Следовательно, сложно-организованными половыми клетками передаются по наследству не готовые структуры, а инструкции по их изготовлению. Эти инструкции не энтелехия, а программа развития путем специфических синтезов, закодированных молекулами ДНК.
МЕТОДИЧЕСКАЯ РАЗРАБОТКА К ПРАКТИЧЕСКИМ ЗАНЯТИЯМ ПО КУРСУ ЭМБРИОЛОГИИ
Задание к практическому занятию № 1
I. ТЕМА: Основы сравнительной эмбриологии. Сущность стадий эмбриогенеза.
II. ЦЕЛЬ ЗАНЯТИЯ: Изучить историю эмбриологии,^половые клетки, основные стадии эмбриогенеза.
III. НЕОБХОДИМЫЙ ИСХОДНЫЙ УРОВЕНЬ ЗНАНИЙ:
1. Понятие об эмбриогенезе, онтогенезе, филогенезе.
2. Строение половых клеток, функциональные и генетические свойства.
3. Классификация яйцеклеток.
4. Основные этапы эмбрионального развития, присущие всем видам животных.
5. Оплодотворение, его биологическая сущность.
6. Дробление, типы дробления зародыша у различных позвоночных.
7. Бластулы, виды бластул.
8. Гаструла и виды гаструляции.
IV. ПРАКТИЧЕСКИЕ ЗНАНИЯ И УМЕНИЯ, КОТОРЫЕ ДОЛЖНЫ БЫТЬ ПОЛУЧЕНЫ СТУДЕНТАМИ:
1. Знать определение эмбриологии, краткий очерк развития эмбриологии.
2. Знать строение половых клеток и уметь их дифференцировать по количеству и распределению желтка.
3. Знать процессы и стадии эмбриогенеза (оплодотворение — зигота, дробление— бластула, гаструляция — гаструла).
4. Знать зародышевые листки, уметь дифференцировать их на препаратах.
V. ВОПРОСЫ, РАССМАТРИВАЕМЫЕ НА ЗАНЯТИИ:
1. Предмет и задачи эмбриологии.
2. История эмбриологии и роль отечественных ученых в развитии эмбриологии.
3. Половые клетки. Строение и функции. Различные типы яйцеклеток.
4. Этапы эмбриогенеза. Оплодотворение, зигота, дробление.
5. Бластула. Виды бластул и их строение.
6. Гаструляция. Основные способы гаструляции. Гаструляция у птиц. Образование первичной полоски.
VI. ЗАДАНИЕ ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ СТУДЕНТОВ: Исследовать под микроскопом, зарисовать и обозначить структуры следующих препаратов:
Препарат № 1. FISSIO IN TOTO INEQUALIS
Дробление полное, неравномерное Окр. пикрофуксин
1. Microblastomerus (микробластомер)
2. Macroblastomerus (макробластомер) Препарат № 2. BLASTULA
Бластула амфибии
1. Blastoderma (стенка бластулы)
2. Крыша
3. Дно
4. Zona narginalis (краевая зона)
5. Blastocelia (бластоцель) Препарат № 3 LINEA PRIMITIVA
Первичная полоска Окр.: гематоксилин
1. Linea primitiva
2. Ectoderma
3. Mesoderma
4. Endoderma
VII. ДЕМОНСТРАЦИОННЫЕ ПРЕПАРАТЫ:
1. Яйцеклетка млекопитающего. Срез яичника млекопитающего. Окр. азаном.
2. Оплодотворение яйцеклетки аскариды. Окр. железным гематоксилином.
3. Полное равномерное дробление зародыша аскариды. Окр. железным гематоксилином.
4. Бластула морского ежа. Окр. гематоксилином.
VIII. ЭЛЕКТРОННЫЕ МИКРОФОТОГРАФИИ:
1. Сперматозоид. Атлас, рис. 510, 512.
2. Яйцеклетка из фолликула яичника. Атлас, рис. 520—522.
IX. КОНТРОЛЬНЫЕ ВОПРОСЫ:
1. Отличие половых клеток от соматических.
2. Биологическая сущность процесса оплодотворения.
3. Чем отличается дробление от деления?
4. Типы бластул в зависимости от типа яйцеклетки.
5. Типы гаструляций у различных типов животных.
X. ЛИТЕРАТУРА:
Обязательная — Гистология под ред. В. Г. Елисеева, 1983 г.г с. 72—98.
Дополнительная — Общая эмбриология, Б. П. Токин, 1977„ с. 84—136; Современная эмбриология, Ч. Бодемер.
Задание к практическому занятию № 2
I. ТЕМА: Дифференцировка" зародышевых листков. Образование провизорных органов.
II. ЦЕЛЬ ЗАНЯТИЯ: Изучить производные зародышевых листков. Знать причины возникновения и усложнения внезародышевых -органов и их функциональное назначение.
III. НЕОБХОДИМЫЙ ИСХОДНЫЙ УРОВЕНЬ ЗНАНИЙ:
1. Определение понятия «гаструляция».
2. Основные способы протекания гаструляции.
3. Зародышевые листки и их значение.
4. Осевой комплекс зачатков у зародыша и его формирование.
5. В незародышевые (провизорные) органы, виды и их функциональное значение.
6. Типы плацент у млекопитающих. '
IV. ВОПРОСЫ, РАССМАТРИВАЕМЫЕ НА ЗАНЯТИИ:
1. Дифференцировка зародышевых листков и их производные.
2. Образование осевого комплекса зачатков органов и их дальнейшая дифференцировка.
3. Образование туловищной складки. Формирование внезародышевых органов (желточный мешок, амнион, серозная оболочка, аллантоис) у птиц.
4. Определение основополагающих направлений в эмбриогенезе: а) стадии; б) процессы; в) элементы морфогенеза; г) элементарные акты.
5. Детерминация и дифференцировка процессов развития.
6. Внутренние и внешние факторы, определяющие детерминацию и дифференцировку (опыт Бигса и Кинга, Гердона и Шпе- :мана).
7. Гипотеза Боннера. Программа развития и сверки ситуации.
8. Основные положения синтетической теории эмбриогенеза.
V. ЗАДАНИЕ ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ СТУДЕНТОВ: Исследовать под микроскопом, зарисовать и обозначить структуры следующих препаратов:
Препарат № 1. Сомиты, хорда, нервная трубка
Окр.: гематоксилин
1. Ectoderma
2. Tuba neuralis
3. Chorda
4. Somitae
5. Nephotom
6. Mesoderma laterale somaticum
7. Mesoderma laterale splanchnicum
8. Coeloma
9. Endoderma
Препарат № 2. Образование туловищных и амниотических
складок
Окр.: гематоксилин
1. Plisa corporis
2. Plisa amniotica
3. Ectoderma
4. Tuba neuralis
5. Chorda
6. Aorta dorsalis
7. Myotomi
8. Coeloma
9. Mesenchyma
10. Endoderma
VI. ДЕМОНСТРАЦИОННЫЕ ПРЕПАРАТЫ:
1. Гаструляция по типу инвагинации и эпиболии. Гаструла* амфибий. Окр.: пикрофуксином.
2. Зародышевый диск курицы. Плоскостной, тотальный препарат. Окр.: кармином.
3. Желточный мешок. Зародыш рыбы с желточным мешком, (поперечный срез). Окр.: пикрофуксином.
4. Смыкание туловищной и амниотической складок. Окр.: гематоксилином.
VII. КОНТРОЛЬНЫЕ ВОПРОСЫ:
1. Что такое «гаструляция»? В чем ее сущность и какие фазы- в ней выделяют?
2. Назовите основные способы гаструляции и приведите примеры.
3. Что является источником хордомезодермального комплекса у ланцетника?
4. Что понимается под комплексом зачатков осевых органов у амфибий, птиц и из чего они образуются?
5. Что такое сомит и на какие части он дифференцируется? В чем сущность дифференцировки?
6. Чем отличаются провизорные органы от деффинитивных к когда они исчезают?
7. У каких представителей позвоночных впервые образуется желточный мешок и в чем заключается его основная функция?
8. Какой из осевых органов является источником развития аллантоиса и какие зародышевые листки принимают участие в его образовании?
9. Перечислите внезародышевые органы у позвоночных, укажите, какие внезародышевые органы формируются только у млекопитающих и какие функции они выполняют.
10. Перечислите зародышевые листки, входящие в состав амниона у птиц, и назовите функции данного внезародышевого органа.
VIII ЛИТЕРАТУРА: Гистология под ред. В. Г. Елисеева,. 1983 г., с. 92—100.
I. ТЕМА: Основы эмбриологии человека. Плацента.
II. ЦЕЛЬ: Изучить основные этапы развития зародыша человека. Развитие, строение и функции плаценты.
Задание к практическому занятию № 3
III. НЕОБХОДИМЫЙ ИСХОДНЫЙ УРОВЕНЬ ЗНАНИЙ:
1. Половые клетки млекопитающих.
2. Этапы эмбрионального развития у позвоночных.
3. Провизорные органы у позвоночных.
4. Роль плаценты в развитии зародыша.
5. Типы плацент.
IV. ВОПРОСЫ, РАССМАТРИВАЕМЫЕ НА ЗАНЯТИЯХ:
1. Половые клетки человека и процесс оплодотворения.
2. Дробление, образование бластулы.
3. Гаструляция и образование провизорных органов.
4. Плацента, ее образование, функции и строение.
V. ЗАДАНИЯ СТУДЕНТАМ ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ: Исследовать под микроскопом, зарисовать препараты и сделать к ним обозначения:
Препарат № 1. Плацента
PLACENTA I. PARS FOETALIS
1. Lamina amnii
2. Epithelium amnii
3. Stratum proprium amnii
4. Lamina chorii
5. Stratum fibrinoideum
6. Villus secundarius
7. Stroma villi
II. PARS UTERINA
8. Decidua basalis
9. Cellula decidualis
VI. КОНТРОЛЬНЫЕ ВОПРОСЫ:
1. Чем дробление у человека отличается от дробления у ланцетника?
2. В чем отличие первичных ворсинок трофобласта от вторичных ворсинок?
3. Почему у человека вначале развиваются внезародышевые части, а затем происходит гаструляция?
VII. ЛИТЕРАТУРА: Гистология под редакцией В. Г. Елисеева» с. 91—97, стр. 596—611.
ОРИЕНТИРОВОЧНАЯ КАРТОЧКА ДЛЯ САМОСТОЯТЕЛЬНОГО ИЗУЧЕНИЯ СТУДЕНТАМ ЙГ^ УЧЕБНОЙ ЛИТЕРАТУРЫ ПРИ ПОДГОТОВКЕ К ПРАКТИЧЕСКИМ ЗАНЯТИЯМ ПО ТЕМЕ «ЭМБРИОЛОГИЯ»
с с |
Содержание и последовательность учебных действий |
Указания к учебным действиям |
Литература |
||
1. |
Определение эмб- |
Обратить внимание на роль |
Уч. Гистологии |
||
риологии. История |
отечественных ученых в истории |
ред. В. Г. Елисе |
|||
развития эмбриоло |
развития эмбриологии (К. Ф. |
ева, 1972, с. 71—72. |
|||
гии. Методы эмб |
Вольф, К. М. Бэр, И. И. Меч |
Б. П. Токин «Общая |
|||
риологии. |
ников, А. О. Ковалевский). Ме |
эмбриология», |
|||
|
тоды эмбриологии: описатель |
1977, с. 9—23. |
|||
|
ный, сравнительный, эволюци |
|
|||
|
онный, экспериментальный. |
|
|||
2. |
Половые клетки. |
Отличие половых клеток от со |
Уч. Гистологии |
||
Мужские половые |
матических. Особенности строе |
с. 72—74, 74—77 |
|||
клетки — сперматозо |
ния сперматозоидов. Классифи |
|
|||
иды. Женские поло |
кация яйцеклеток по количест |
|
|||
вые клетки — яйце |
ву и распределению желтка |
|
|||
клетки. |
(изолецитальная, телолециталь- |
|
|||
|
ная,— умеренно и резко, вторич |
|
|||
|
но изолецитальные). |
|
|||
3. |
Стадии и процессы |
Обратить внимание на отли |
Уч. Гистологии |
||
эмбрионального раз |
чие процесса от стадий и сущ |
с. 77—91. |
|||
вития: |
ность каждого процесса. Опло |
Б. П. Токин «Общая |
|||
Оплодотворение — |
дотворение — зигота, дробле |
эмбриология», |
|||
зигота |
ние — бластула, гаструляция — |
с. 447—481. |
|||
Дробление — бла |
гаструла. Дифференцировка — |
|
|||
стула |
гистогенез и детерминация. Ор |
|
|||
Гаструляция — |
ганогенез. Объяснить зависи |
|
|||
гаструла |
мость последующих стадий от |
|
|||
Нейрулядия — ней- |
предыдущих в свете биогенети |
|
|||
рула |
ческого закона Мюллера, Гек- |
|
|||
Гистогенез — ткани |
келя, Северцева. |
|
|||
Органогенез — орган |
|
|
|||
4. |
Образование внеза- |
1. Уметь схематически изобра |
|
||
родышевых (прови |
зить процесс формирования трех |
|
|||
зорных) органов у |
зародышевых листков (эктодер |
|
|||
птиц: желточный ме |
мы, мезодермы, энтодермы) и |
|
|||
шок, амнион, сероза, |
осевых органов на примере раз |
|
|||
алантсис. Функцио |
вития птиц. |
|
|||
нальное значение |
|
|
|||
внезародышевых ор |
|
|
|||
ганов. Развитие, |
|
|
|||
строение и функции |
|
|
|||
плаценты. |
|
|
|||
|
2. Обратить внимание на об |
Уч. Гистологии |
|||
|
разование туловищной и амнио- |
с. 91—93. |
|||
|
тической складок и их значе |
Б. П. Токин «Общая |
|||
|
ние для последующего развития |
эмбриология», |
|||
|
3. Этапы развития плаценты. |
с. 152—155. |
|||
|
Строение и функции плаценты |
|
|||
|
в системе плод — плацента — |
|
|||
|
мать. |
|
|||
ВОПРОСЫ к СЕМИНАРСКОМУ ЗАНЯТИЮ И К ЭКЗАМЕНУ ПО РАЗДЕЛУ «ЭМБРИОЛОГИЯ»:
1. Определение понятия «эмбриология». Задачи предмета.
2. Краткий обзор истории эмбриологии.
3. Основные положения теории преформизма и эпигенеза.
4. Сущность закона зародышевого сходства К. Бэра.
5. Основные положения биогенетического закона (Геккель — Мюллер—Северцев).
6. Современная эмбриология. Методы и задачи.
7. Характеристика половых клеток. Их значение в процессе развития.
8. Строение сперматозоида.
9. Женская половая клетка. Особенности ее строения.
10. Роль кортикальной зоны яйцеклетки для осуществления: дальнейшего развития.
11. Типы яйцеклеток в зависимости от количества и характера расположения запасного трофического материала.
12. Изолецитальная яйцеклетка (ланцетник).
13. Телолецитальные яйцеклетки (умеренно и резко — амфибии, рептилии и птицы).
14. Особенности яйцеклетки человека в связи с условием развития зародыша.
15. Эмбриональный период. Его определение и характеристика.
16. Этапы эмбриогенеза.
17. Биологическая сущность оплодотворения.
18. Зигота — стадия одноклеточного организма.
19. Виды дробления.
20. Дробление как разновидность клеточного деления, его биологические особенности. Связь типа дробления с типом яйцеклетки.
21. Бластула.
22. Типы бластул. Связь типа бластулы с характером дробления зиготы и типом яйцеклетки.
23. Бластула ланцетника, амфибий, птиц.
24. Процесс гаструляции.
25. Характеристика всех типов гаструляции.
26. Особености гаструляции ланцетника.
27. Отличительные особенности гаструляции птиц.
28. Нейруляция. Характеристика и значение этого процесса.
29. Образование туловищной и амниотической складки у птиц. Их значение и производные.
30. Провизорные органы птиц. Источники образования.
31. Строение и функции серозной оболочки, амниона, аллантоиса, желточного мешка.
32. Ранние стадии развития зародыша человека.
■ 33. Провизорные органы зародыша человека.
34. Типы планцет у разных классов позвоночных.
35. Развитие, строение и функции планцеты у человека.
36. Определение понятий эмбриогенеза: стадии, процессы, элементы, элементарные акты морфогенеза.
37. Детерминация и дифференцировка процессов развития.
38. Внутренние и внешние факторы, определяющие дифферен- цировку. (Опыты Бриггса, Кинга, Гердона, Шпемана, Ру).
39. Понятие о критических периодах. (П. Г. Светлов).
40. Нарушение процессов детерминации и дифференцировки как причина аномалий и уродства.
41. Основные периоды развития зародыша человека.
42. Регуляция работы генов. Схема Дэвидсона и Бриттена.
43. Репрессия генов.
44. Дерепрессия генов.
45. Современная генетическая теория развития зародышей.
ЗАДАЧИ ПО ЭМБРИОЛОГИИ
Задача 1. На электоронограмме представлены мужские и женские половые клетки. Как по составу органелл можно отличить яйцеклетку от сперматозоида?
Задача 2. Половая клетка окружена двумя оболочками: блестящей и лучистым венцом. Назовите эту клетку. Какие клетки принимают участие в образовании этих оболочек?
Задача 3. На электронных микрофотографиях представлены поперечные срезы сперматозоидов. На одном хорошо прослеживаются осевые нити, окруженные митохондриями, на другом видна только центриоль. Назовите, какие отделы клетки представлены на фотографиях.
Задача 4. При оплодотворении в яйцеклетку попадает сперматозоид, несущий «У» хромосому. Какой пол будущего зародыша?
Задача 5. На рисунке изображены яйцевые клетки ланцетника, лягушки, курицы и человека. К какому типу яйцевых клеток по количеству и распределению желтка, принадлежит каждая из них?
Задача 6. У ланцетника на стадии двух бластомеров в эксперименте уничтожен один бластомер. Что произойдет с оставшимся?
Задача 7. На препарате виден зародыш, состоящий из четного числа бластомеров, имеющий одинаковую величину. Определите, какой тип дробления характерен для этого зародыша.
Задача 8. На стенде выставлены препараты зародышей на стадии развития дискобластулы, амфибластулы и стерробластулы. К каким классам представителей хордовых относятся эти зародыши?
Задача 9. Яйцеклетка содержит мало желтка и распределен он равномерно. Определите тип яйцеклетки, характер дробления, вид бластулы будущего зародыша. Каким представителям животного мира свойственны такие яйцеклетки?
Задача 10. Даны два зародыша одного вида животных. Один на стадии двух бластомеров, другой на стадии морулы. Какой зародыш больше по массе?
Задача 11. В результате дробления у одного зародыша образуются микро- и макробластомеры. Для каких яйцеклеток характерно наличие таких бластомеров?
Задача 12. У зародыша дробление отмечено только на анимальном полюсе. Как называется такой вид дробления и тип яйцеклетки?
Задача 13. Желтка в яйцеклетке много, концентрируется на вегетативном полюсе. Как называется такая яйцеклетка и для какого класса животного мира она характерна?
Задача 14. На препарате бластула с многослойной бластодермой. Бластоцель располагается эксцентрично. Определить тип яйцеклетки, характер дробления и какому классу животных это> свойственно?
Задача 15. В рёзультате дробления зародыша возникла цело- бластула. Определите тип яйцеклетки и характер дробления.
Задача 16. Дробление зиготы происходит полностью, но неравномерно. Определите тип яйцеклетки и вид бластулы.
Задача 17. Дробление зиготы дискоидальное. Определите тип яйцеклетки и класс животных, для которых характерно такое дробление.
Задача 18. На препарате бластула с однослойной бластодермой,, бластоцель в центре. Определить, какой тип дробления обеспечивает образование данной бластулы.
Задача 19. В эксперименте на стадии бластулы введено вещество, блокирующее перемещение клеток. Развитие какой стадии: эмбриогенеза будет блокировано?
Задача 20. На рисунке изображена гаструла ланцетника, амфибии, птиц. Какой тип гаструляции свойственен каждому из перечисленных представителей?
Задача 21. В эксперименте у зародыша на стадии гаструлы блокирован процесс перемещения клеток через первичную полоску. Развитие какого зародышевого листка будет нарушено?
Задача 22. В эксперименте у зародыша птицы на стадии гаструлы блокирован процесс перемещения клеток через головной узелок. Развитие какого осевого органа будет нарушено?
Задача 23. Перед исследователем лежат препараты серийных поперечных срезов зародышей цыпленка через 12 часов инкубирования их в термостате. На одном из препаратов зародышевые листки связаны между собой плотным скоплением клеток. Какой стадии развития соответствует зародыш и на каком уровне произведен срез?
Задача 24. В условном эксперименте микроманипулятором разрушили миотом. Нарушение развития какой ткани вызовет это воздействие?
Задача 25. В условном эксперименте блокировано перемещение клеточного материала через первичную полоску и головной узелок. Какое нарушение в развитии зародыша вызовет это воздействие?
Задача 26. В условном эксперименте микроманипулятором разрушили дерматом. Нарушение развития какой ткани вызовет это воздействие?
Задача 27. Экспериментальным путем у зародыша поврежден нефротом. Нарушения в каких системах развития произойдут?
Задача 28. В эксперименте на амфибиях на стадии гаструлы •с помощью сплошной пластинки изолировали хордальный вырост ■от эктодермы. Какие нарушения в развитии зародыша вызовет это воздействие?
Задача 29. В эксперименте на головастике лягушки на стадии ранней гаструлы хордальный вырост пересадили под эктодерму на вентральную часть зародыша. Какие изменения произойдут в результате этого воздействия?
Задача 30. В эксперименте над зародышами амфибий на стадии поздней гаструлы сделана пересадка области дорзальной губы бластопора одного зародыша на вентральную поверхность другого зародыша. К каким последствиям это приведет? Идентичны ли по результатам подобные эксперименты на стадии ранней гаструлы?
Задача 31. У зародыша помимо других провизорных органов, развивается трофобласт. К какому классу животных относится этот зародыш?
Задача 32. У зародышей развиты все провизорные органы: желточный мешок, амнион, серозная оболочка и аллантоис. К каждому классу животных следует отнести эти зародыши?
Задача 33. В эксперименте у зародыша цыпленка нарушен процесс срастания амниотических складок. Образование каких провизорных органов будет нарушено?
Задача 34. У зародышей, относящихся к типу позвоночных, из всех провизорных органов развивается только желточный мешок. К какому классу животных относятся зародыши и какой образ .жизни ведут их взрослые особи?
Задача 35. Исходя из главной функции желточного мешка — депонирование питательных веществ и образование первичных эритробластов, у каких рыб — мелководных или глубоководных — можно ожидать ускорение развития кровяных островков?
Задача 36. При развитии цыпленка образуется амниотическая складка. Какими зародышевыми листками она представлена и какие оболочки она образует?
Задача 37. В эксперименте у зародыша цыпленка повреждена внезародышевая латеральная мезодерма. Образование каких внезародышевых органов будет нарушено?
Задача 38. При развитии зародыша цыпленка поврежден орган, выполняющий функции газообмена и выделения метаболитов. Как называется этот орган? Какие листки его образуют?
Задача 39. При развитии зародыша птицы повреждена внезародышевая эктодерма. В составе каких внезародышевых оболочек она входит и какие функции будут нарушены?
Задача 40. У женщины в результате воспалительного процесса произошла облитерация (закрытие просвета) обоих яйцеводов. Возможно ли оплодотворение в этих условиях?
Задача 41. В процессе сперматогенеза у человека нарушено формирование акросомы (симптом «округлой головки сперматозоида»). Какая функция сперматозоида будет изменена? Возможно ли оплодотворение в этом случае?
Задача 42. Яйцеклетка оплодотворена двумя сперматозоидами. Каков хромосомный набор зародыша? Возможно ли его нормальное развитие?
Задача 43. При цитологическом исследовании околоплодных вод обнаружены клетки, содержащие половой хроматин. Каков пол ребенка?
ЗАДАЧИ ПО ЭМБРИОЛОГИИ ЧЕЛОВЕКА
Задача 44. На гистологическом препарате яйцеклетка человека, в цитоплазме которой имеется небольшое количество желточных включений, распределенных равномерно. Какой тип яйцеклетки?
Задача 45. У человека сперматозоиды направленно движутся в сторону яйцеклетки и оболочки половых клеток вступают в специфические контакты. Как называется это направленное движение?
Задача 46. Яйцеклетка человека оплодотворена сперматозоидом, содержащим У-хромосому. Каков будет пол ребенка?
Задача 47. Проникновение одного сперматозоида в яйцеклетку человека предотвращает возможность проникновения остальных сперматозоидов. Назовите вид оплодотворения и что препятствует проникновению в одну яйцеклетку более одного сперматозоида?
Задача 48. В результате 2-го дробления у человека образуется три различных по величине бластомера. Определите тип дробления зиготы.
Задача 49. Зародыш человека состоит из 8 бластомеров. Определите приблизительный срок беременности и место нахождения зародыша.
Задача 50. При дроблении зиготы образовались светлые и темные бластомеры. Какие бластомеры являются источником развития эмбриобласта?
Задача 51. При дроблении зиготы образовались светлые и темные бластомеры. Какие бластомеры являются источником развития трофобласта?
Задача 52. При дроблении зиготы образовались темные и -светлые бластомеры. Светлые бластомеры дробятся и обрастают одним слоем темные. Какая плодная оболочка образуется из ♦светлых бластомеров?
Задача 53. В зародыше человека образуется полость и происходит дифференцировка бластомеров. На какой стадии развития находится зародыш? Где это происходит? Какие образования являются результатом дифференцировки?
Задача 54. Для развития человека характерно развитие трофобласта, который на второй неделе дифференцируется на два слоя. Как называются эти слои и какими гистологическими структурами они образованы?
Задача 55. Зарегистрировано начало имплантации зародыша человека. На какой стадии находится зародыш? Каково число бластомеров и возраст зародыша?
Задача 56. Имплантации и дальнейшему развитию эмбриона человека сопутствует трансформация стромы эндометрия. Каков •ее результат?
Задача 57. На препарате разрез 14-дневного зародыша человека, где в полости плода видны два пузырька. Как называются эти пузырьки? Какие зародышевые листки образуют контактирующие стенки?
Задача 58. На определенном этапе развития у человека между сосудистой системой матери и плода устанавливается особая функциональная связь. Какой орган опосредует эту связь и с какой недели беременности?
Задача 59. Представлены препараты плодной и материнской части планцеты. Какие структурные образования входят в состав плодной части плаценты?
Задача 60. Представлены препараты плодной и материнской части плаценты. Какие структурные образования входят в состав материнской части плаценты?
Задача 61. На препарате материнская часть планцеты, в слизистой оболочке которой расположены крупные клетки со светлой цитоплазмой и округлым ядром. Как называются эти клетки и какую функцию они выполняют?
Задача 62. «Человек родился в рубашке». О какой «рубашке» говорится в пословице и каков источник развития эпителия, ее изнутри выстилающего?
Задача 63. Клеточный материал эмбриобласта зародыша человека становится двуслойным. Какой механизм образования слоев, стадия эмбриогенеза, возраст зародыша?
положение дополнялось виталистическим взглядом о том, что ма- тёриал материнского зачатка приобретает форму будущего организма под влиянием нематериальных сил — «энтелехий», заложенных в семени отца.
Виталистический взгляд Аристотеля в XVII веке был возрожден У. Гарвеем (1578—1657), предложившим и термин «эпигенез». У. Гарвей выдвинул принцип «все из яйца». Причем у животных, у которых ему не удалось обнаружить яиц, он допускал их зарождение из «разлагающихся веществ».
Однако, в обеих этих интерпретациях теория эпигенеза успеха не имела, и во второй половине XVII и в первой половине XVIII века господствовала теория преформизма.
Теория преформизма вновь возродилась во второй половине XVIII века в связи с распространением учения о всемирном тяго-. тении и влиянии его на мир химических и биологических процессов.
Обоснование эпигенезу дал К. Ф. Вольф (1733—1794). Он утверждал, что лишь в результате поступления извне питательных соков в зачатке образуются клетки и сосуды, из которых затем формируются органы будущего зародыша. К. Ф. Вольф ссылался на наличие «существенной силы», которая обуславливает наступление и продвижение питательных веществ, необходимых для образования частей зародыша.
Борьба теорий преформизма и эпигенеза стала особенно ожесточенной в конце XIX века в связи с развитием экспериментальной эмбриологии и, так называемой механики развития, претендовавших на каузальное объяснение онтогенеза.
Однако, в те времена речь шла уже не о наличии в зачатковых клетках готового миниатюрного зародыша, а о химических и морфологических структурах, предопределяющих развитие организма. Эти положения дополнялись виталистическим взглядом о том, что материал материнского зачатка приобретает форму будущего организма под влиянием нематериальных сил — «энтелехий», заложенных в семени отца.
Полемика между сторонниками эпигенеза и преформизма велась также вокруг проблемы соотношения внутренних и внешних факторов наследственности в эволюции. Представители эпигенеза пытались обосновать свои взгляды с ссылкой на явления наследования приобретенных признаков, на возможность влияния внешней среды на онто- и филогенез, т. е. опирались на различные ламарк- систские теории. Однако, многочисленные попытки подтвердить эти представления экспериментально оказались безуспешными. Вместе с тем успехи молекулярной биологии и генетики во второй половине XX столетия доказали наличие в зачатковых клетках предетермированных химических и биологических структур (молекул ДНК), которые определяют воспроизведение основных видов и индивидуальных особенностей развивающегося организма.
СОВРЕМЕННАЯ ЭМБРИОЛОГИЯ
Ставит своей задачей выявление движущих сил эмбрионального формообразования, вскрытие генетических и цитологических основ, клеточной, тканевой и органной дифференцировки.
В 20—40 годы XX века большую роль в развитии эмбриологии сыграли работы Г. Шпемана и его школы по изучению влияния одних частей зародыша на другие; были выведены понятия «индуктор», «организатор».
Д. П. Филатов (1876—1943) и другие советские исследователи развили учение Г. Шпемана и внесли в него существенные поправки.
Д. П. Филатов связал экспериментальную эмбриологию с эволюционным учением и сформулировал понятие о формообразовательном аппарате, т. е. тех частей зародыша, взаимодействие которых приводит к осуществлению определенных этапов развития.
Советские ученые сделали существенный вклад и в понимание закономерностей индивидуального развития. Н. И. Кольцов впервые произвел попытку синтеза эмбриологии и генетики. -П. Г. Светлов предложил оригинальный вариант теории «критических периодов» в развитии организма. Б. П. Токин начал исследование соматического эмбриогенеза, т. е. изучение развития организмов из соматических клеток.
В настоящее время процесс развития понимается как реализация тех возможностей, которые заложены в «наследственном коде» половых клеток. Хотя для осуществления этих процессов развития необходимо наличие определенных условий среды, они йеизменно реализуются путем самодифференцировки на основе тех видоспецифических закономерностей, которые были исторически «выработаны и закреплены на протяжении эволюции органического мира.
ПОЛОВЫЕ КЛЕТКИ ИЛИ ГАМЕТЫ
Сперматозоиды позвоночных имеют жгутиковую форму и обладают способностью к активному движению. В них различают головку, шейку, тело и хвост.
Головка сперматозоида содержит плотное ядро, окруженное тонким слоем цитоплазмы. В ядре содержится гаплоидный набор хромосом, содержащих ДНК, РНК» протамины, гистоны. Сверху головка сперматозоида покрыта чехликом, в котором располагается плотная гранула — акросома. Чехлик и акросома являются производными пластинчатого комплекса Гольджи. Акросома содержит фермент гиалуронидазу, способную растворять оболочки, покрывающие яйцеклетки.
Задача 64. В наружном листке зародышевого щитка определяется координированное перемещение клеточных масс в каудальном направлении? Какая структура при этом образуется, на такой неделе развития? Как называется эта стадия эмбриогенеза?
Задача 65. У зародыша человека началась закладка осевых органов. Каков возраст зародыша?
Задача 66. У эмбриона человека определяется 7 пар сомитов. Каков возраст эмбриона?
Задача 67. В зародыше человека регистрируется процесс обособления его тела от провизорных органов. Образование какой структуры приводит к этому и каков возраст зародыша?
Задача 68. При развитии человека образуется желточный мешок, который не содержит желтка. Какую функцию выполняет этот орган?
Задача 69. На серийных поперечных срезах зародыша человека обнаруживается замыкание нервной трубки на всем протяжении, исключая невропор. Какой неделе развития соответствует данное состояние?
Задача 70. Инъекция мочи беременной женщины неполовозрелым мышам вызывает у последних бурное созревание фолликулов яичника (ранний клинический тест на выявление беременности). Какое вещество, содержащееся в моче, определяет данный гонадотропный эффект и где оно синтезируется?
Задача 71. Дифференцировка эпителия в культуре возможна лишь в присутствии мезенхимы. Пересадка спинной губы бластопора стимулирует развитие нервной трубки в прилежащей эктодерме. Какой механизм развития демонстрирует эти примеры?
Задача 72. Ребенок первых месяцев жизни обладает толерантностью к ряду инфекционных болезней. Чем определяется иммунитет новорожденного?
Дополнительная литература
1. Токин Б. П. Общая эмбриология М., Высшая школа. 1977 г.
2. Бодемер. Современная эмбриология. М., «Мир». 1971 г.
3. Кнорре А. Г. Эмбриональный гистогенез. Л. «Медицина», 1971 г.
4. Кнорре А. Г. Краткий очерк эмбриологии человека. Л., «Медицина»,. 1963 г.
5. Пэттэн Б. М. Эмбриология человека. М., «Медицина», 1959 г.
6. Волкова О. В., Тарабрин С. Б. Эмбриогенез человека и его регуляция. М., Изд-во 2 МОЛГМИ, 1973 г.
7. Волкова О. В., Пекарский М. И. Эмбриогенез и возрастная гистология внутренних органов человека. М., «Медицина». 1976 г.
8. Фалин Л. И. Эмбриология человека. Атлас. М., «Медицина», 1976 г.
9. Субботин М. Я., Донских Н. В. Гистофизиология и гистопатология внезародышевых органов человека и млекопитающих. Новосибирск, 1975 г.
10. Субботин М. Я. Лекции по эмбриологии. Новосибирск, 1977 г.
11. Гармашева Н. Л., Константинова Н. Н. Введение в перинатальную медицину. М., «Медицина», 1978 г.
12. Мучиев Г. С., Фроленко О. Г. Охрана здоровья плода и новорожденного в СССР. М., «Медицина», 1979 г.
13. Браун Д., Диксон Г. Антенатальная охрана плода. М., «Медицина», 1982 г.
14. Хэм А., Кормак Д. Гистология. Изд. «Мир», 1983, т. 5.
15. Брусиловский А. И. Развитие, строение и функции плаценты человека. Симферополь, 1986 г.
Шейка сперматозоида содержит проксимальную центриоль, играющую важную роль в делении оплодотворенного яйца, и дистальную, от которой отходит осевая нить, проходящая через тело и хвост. Тело сперматозоида содержит митохондрии, которые по спирали окружают осевую нить, а также вторую половину дистальной центриоли. В теле находятся запасы гликогена.
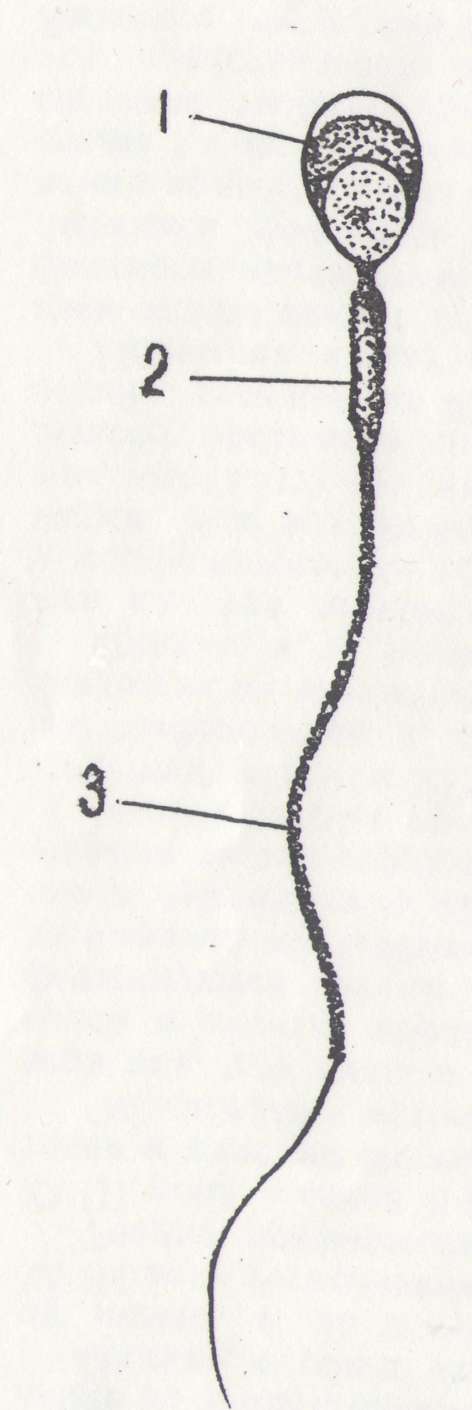
Рис. 1. Строение сперматозоида .
1—головка; 2 — шейка; 3 — хвост; 4 — акросома, 5 — ядро; .6 дистальная центриоля; 7 — проксимальная центриоля; 8 — митохондрии; 9 дипломакро-фибриллы центральные и периферические
Хвост сперматозоида содержит осевую нить, которая окружена небольшим количеством цитоплазмы, содержащей фермент адено зинтрифосфатазу. Скорость движения сперматозоида 2—3 мм в минуту (рис. 1) .
Женская половая клетка — яйцеклетка — имеет округлую форму и не обладает способностью к самостоятельному движению. Яйцеклетка содержит крупное ядро с гаплоидным набором хромосом, ядрышко и ооплазму, в которой располагается питательный материал в виде желтка. Величина яйцеклетки зависит от количества отложенного в ооплазме желтка.
Размеры яйцеклеток млекопитающих имеют диаметр 50—150 микрометров. Яйцеклетка содержит слаборазвитую цитоплазматическую сеть, митохондрии, пластинчатый комплекс Гольджи. В зрелой яйцеклетке отсутствует центросома. По периферии располагаются кортикальные гранулы, содержащие кислые мукополи- сахариды. Яйцеклетка покрыта первичной оболочкой, которая образуется путем затвердения наружного слоя цитоплазмы в ходе оогенеза, и вторичной оболочки, образованной фолликулярными клетками и называемой блестящей оболочкой. Ее наружная часть, состоящая из кислых мукополисахаридов, секретируется фолликулярными клетками. Внутренняя зона, состоящая из нейтральных мукополисахаридов, образуется ооцитом. Фолликулярные клетки выполняют трофическую функцию, доставляя в яйцеклетку питательные вещества.
ТИПЫ ЯЙЦЕКЛЕТОК
Классификация яйцеклеток основана на количестве и распределении желтка, содержащегося в ооплазме. Желток — совокупность включений, состоящих из питательных веществ. Яйцеклетки с небольшим количеством желтка, распределенном равномерно по всей цитоплазме, называются изолецитальными. Они характерны для ланцетника и морских ежей (рис. 2а).
Яйцеклетки со средним количеством желтка; распределенном на нижнем вегетативном полюсе яйца, называются умеренно тело- лецитальными. Цитоплазма концентрируется на верхнем анималь- ном полюсе. Такие яйцеклетки характерны для амфибий (рис. 26).
Яйцеклетка, содержащая большое количество желтка, заполняющего почти всю яйцеклетку, называется резко телолецитальной. Примером являются яйцеклетки акуловых, костистых рыб, пресмыкающихся, птиц и яйцекладущих млекопитающих (ехидна, утконос).
У млекопитающих и человека в связи с внутриутробным развитием и питанием зародыша за счет материнского организма, яйцеклетка в процессе эволюции стала изолецитальной, возникнув из телолецитальной. Такие яйцеклетки, называются вторично х изолецитальными.