

§ 3.5 Результаты экспериментов и их обсуждение
Для регистрации осмотического сжатия эритроцитов пробы крови – были получены временные ходы концентрации глюкозы и показателя гематокрита в пробе крови, подготовленной по описанию пробы №1 в § 3.3 «Используемые материалы». Измерения указанных характеристик проводились параллельно в течение 3-х часов после взятия пробы крови из организма. Методика проведения этих экспериментов описана в § 3.4 «Методики проведения экспериментальных работ» в пунктах 1, 2.
Типичный результат первых опытов (рассмотрено более 10 проб крови от различных доноров), полученных ранее сотрудниками нашей группы [20], представлен на рис. 16. На нем показан временной ход концентрации глюкозы (верхняя кривая) и временной ход показателя гематокрита (нижняя кривая) пробы крови.
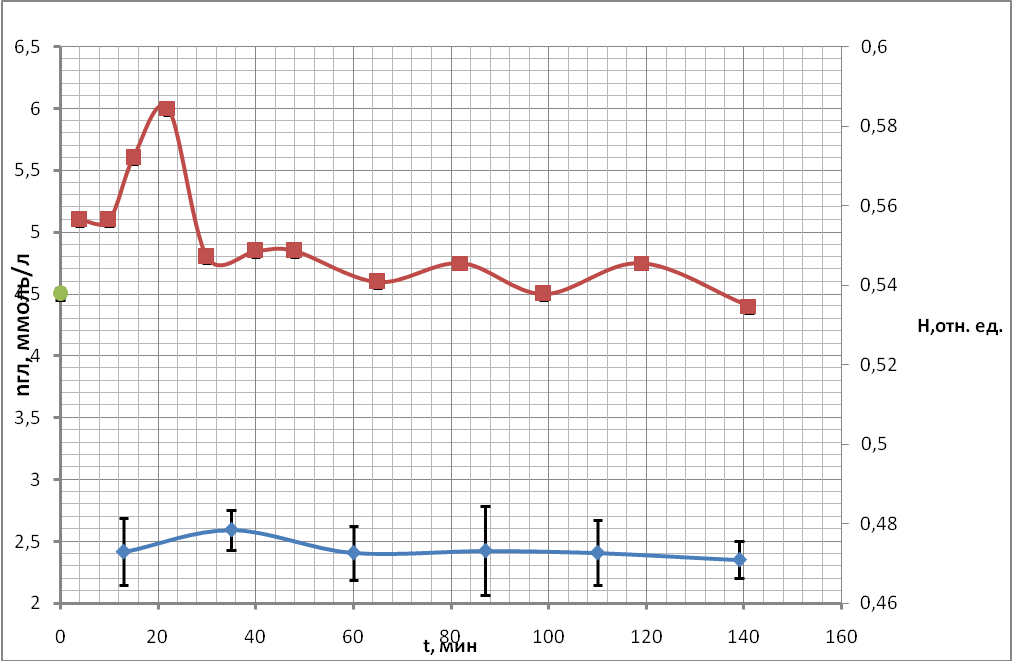
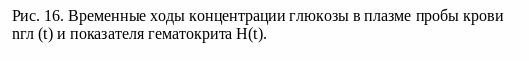
Из графика видно, что начальная экспериментальная точка во всех опытах измерения концентрации глюкозы, полученная в пробе крови, оказывалась выше, чем в пробе крови, взятой непосредственно из пальца (она показана на оси ординат, где представлены значения концентрации глюкозы). Первая экспериментальная точка из пробы была получена через 3-4 минуты после измерения концентрации глюкозы в плазме крови, полученной из пальца. Погрешность измерения концентрации глюкозы была посчитана по формуле (5) и ее значение не выходит за размеры точки на графике.
Временной ход показателя гематокрита, как видно из рис. 16, в пределах погрешности меняется незначительно. Следовательно, по этой характеристике нельзя судить о первоначальном изменении объема эритроцитов.
Теоретический анализ показывает, что первоначальное осмотическое сжатие эритроцитов имеет место из-за распада пептидов в плазме пробы крови, осуществляемого пептидазами. Однако в представленном эксперименте это сжатие не наблюдалось. Было сделано предположение о том, что осмотические процессы настолько быстры, что мы не успеваем их увидеть за время, прошедшее от взятия крови из вены до первого измерения (не менее 2-х минут).
Для того чтобы убедиться в правильности этого предположения, была проведена отдельная серия опытов. Были получены временные ходы концентрации глюкозы в пробах крови, подготовленных по описанию проб №1, №2 (с 50 мкл антипептидаз) в § 3.3 «Используемые материалы». Антипептидазы ингибируют ферментативный распад пептидов на отдельные аминокислоты, тем самым увеличивая время, за которое происходит этот распад.
Типичный результат показан на рис. 17 приведены результаты такого опыта. Временной ход показателя гематокрита не приводится из-за его не информативности, так как время одного измерения больше характерного времени протекания быстрых осмотических процессов, описанных выше.

На рис. 17 представлены временные ходы концентрации глюкозы в плазме пробы крови: 1 – без добавления антипептидаз, 2 – с добавлением антипептидаз. Видно, что кривая 2 имеет «провал» в значениях концентрации глюкозы в первый час наблюдения. Ход кривой 1 не отличался от хода кривых в экспериментах, сделанных ранее.
Такое уменьшение концентрации глюкозы в пробе крови с применением антипептидаз свидетельствует о наличии сжатия эритроцитов. Таким образом, наше предположение о том, что ферментативный распад пептидов в плазме крови вызывает осмотическое сжатие эритроцитов и, что эти процессы происходят очень быстро (за время меньшее 5 мин), верно. Такое первоначальное осмотическое сжатие может дать начало неравновесным процессам, впоследствии приводящим к установлению нового уровня энергетического метаболизма.
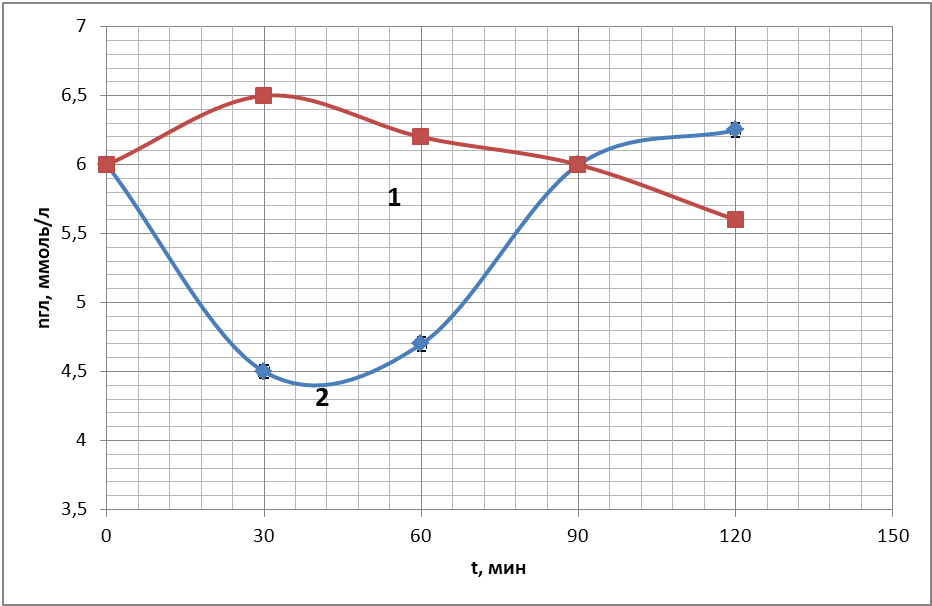
В опытах, ставящих своей целью регистрацию колебательных процессов, были получены временные ходы концентрации глюкозы в пробах крови, подготовленных по описанию проб №1, №2 (с 50 мкл антипептидаз), №3 (250 мкл антипептидаз), №4 (с цитратом натрия) в § 3.3 «Используемые материалы». Время наблюдения колебаний продолжалось в течение 1 часа. Чтобы получить осмотические колебания, предсказанные теоретическим анализом, получали экспериментальные точки через максимально короткий промежуток времени для всех 4-х пробирок параллельно (раз в 2 минуты). Временные ходы концентрации глюкозы для 4 проб показаны на рис.18 (а, б, в, г) в течение одного часа. Во всех 4-х пробах крови наблюдались осмотические колебания.
а)
б)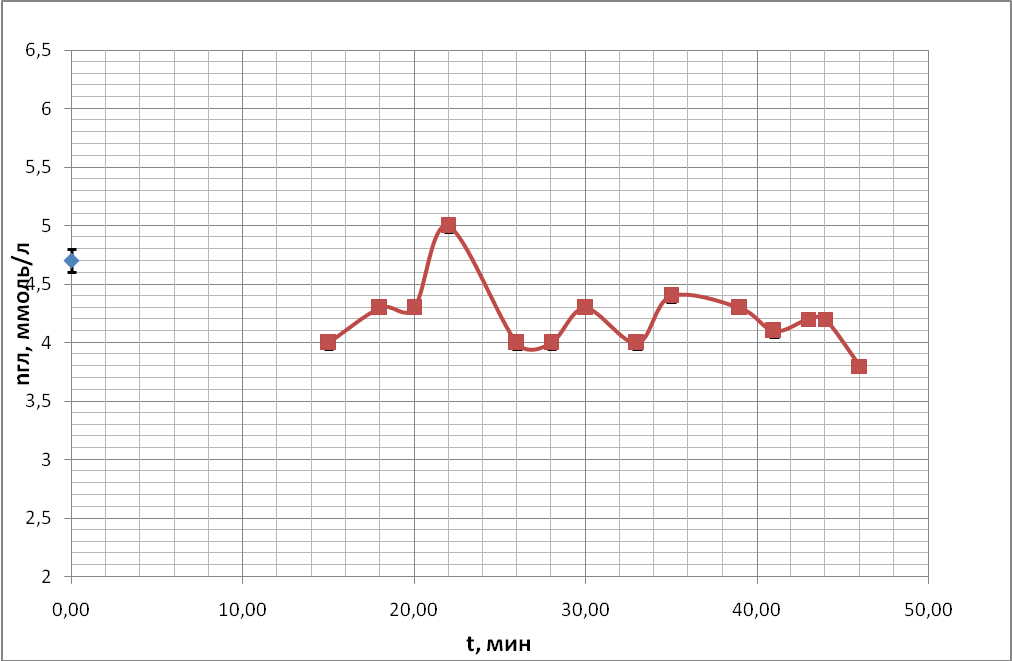
в)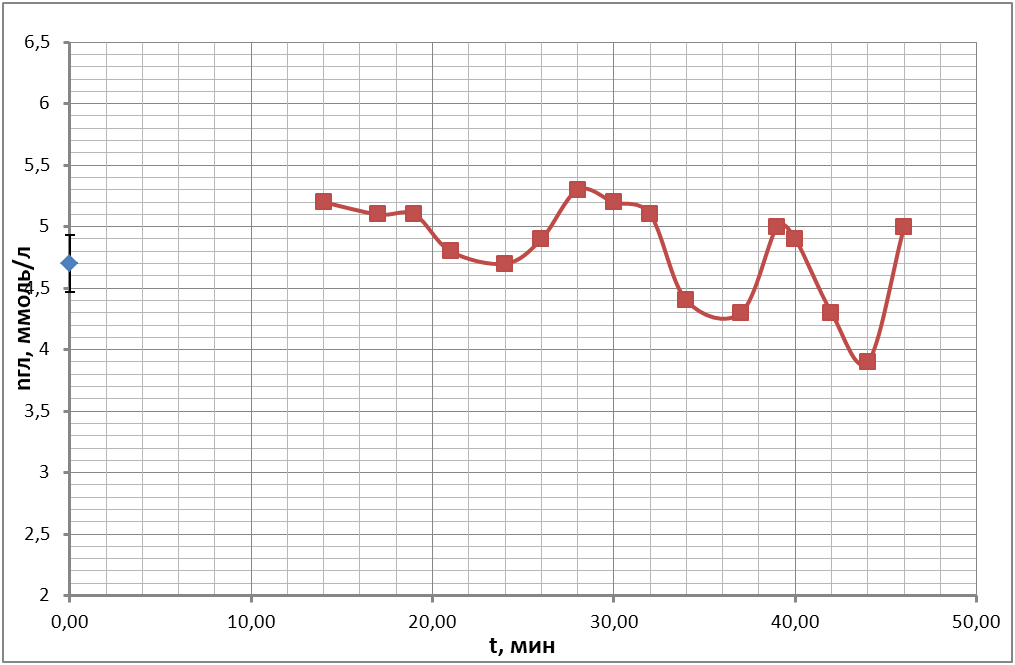
г)
Таким образом, из этих экспериментов видно, что в пробах крови происходят быстрые колебательные процессы, связанные с рассогласованием процессов энергетического метаболизма, как это предсказано теоретическим анализом.
Для того, чтобы определить оптимальное время после взятия крови из организма для проведения измерения неспецифической проницаемости эритроцитарных мембран для кислорода, была проведена следующая серия экспериментов. Были получены временные ходы концентрации глюкозы в пробах крови, подготовленных по описанию проб №1, №2 (с 250 мкл антипептидаз), №3 (500 мкл антипептидаз) в § 3.3 «Используемые материалы». Полученные кривые представлены на рис. 19 (а, б, в).
а)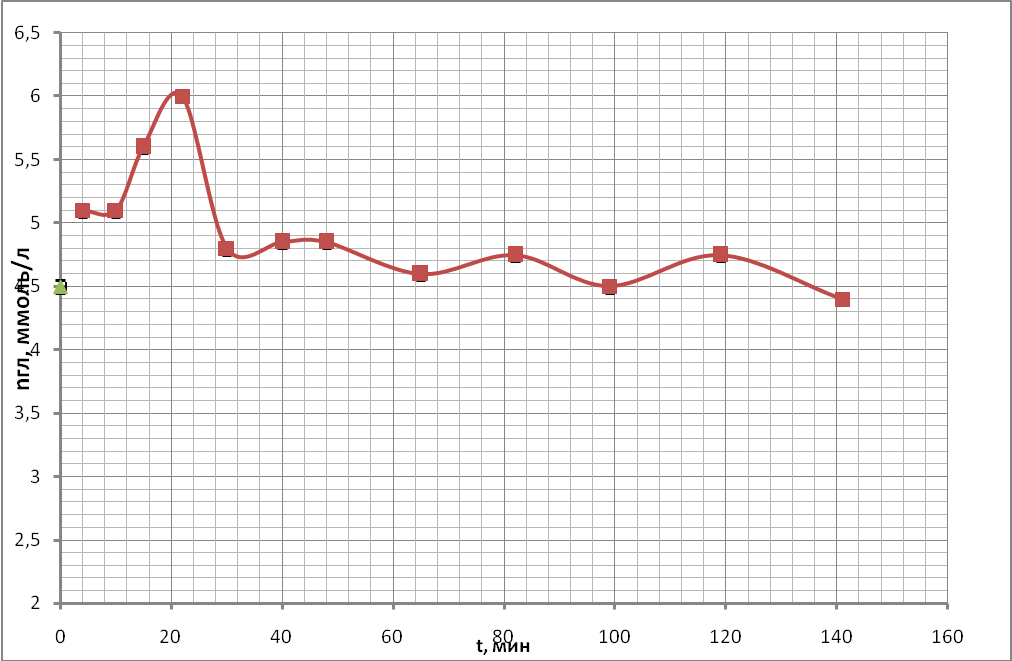
б)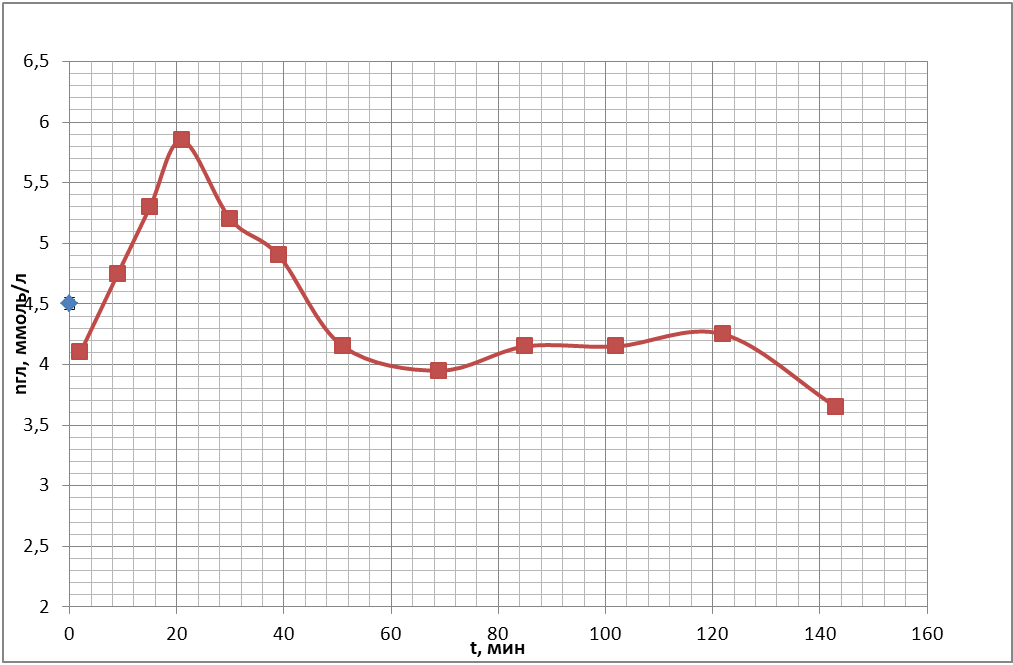
в)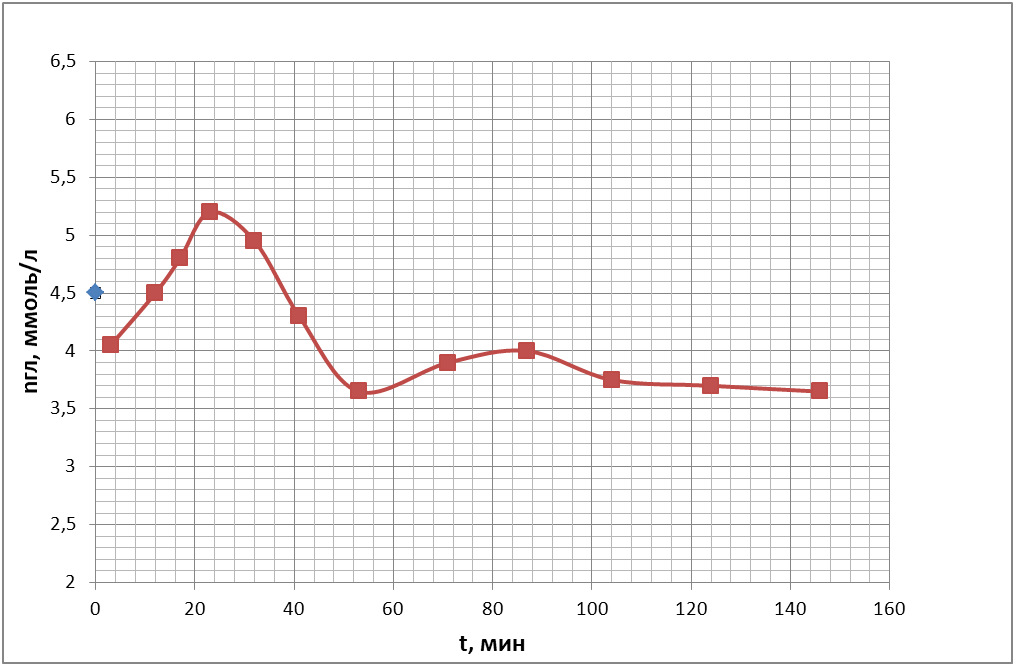

Из рисунков видно, что через час после взятия крови из организма происходит относительная стабилизация концентрации глюкозы в плазме проб крови, когда колебания этого параметра становятся много меньше колебаний, происходящих в первый час наблюдений. Таким образом, проницаемость в следующей серии опытов необходимо измерять через час после взятия крови из организма.
Следующая серия экспериментов ставила своей целью регистрацию факта совершившегося перехода метаболизма эритроцитов из состояния с высокой концентрацией АТФ в состояние с низкой его концентрацией по выявленным отличиям параметров различных проб крови. Теоретический анализ показывает, что этот переход должен совершиться быстрее в пробах, в которых нет антипептидаз, по сравнению с пробами, в которых они есть. Были получены временные ходы концентрации глюкозы и показателя кислотности рН в пробах крови, подготовленных по описанию проб №1, №2 (с 500 мкл антипептидаз), №3 (1000 мкл антипептидаз), №4 (с этиловым спиртом) в § 3.3 «Используемые материалы». В данной серии использовался спирт из-за того, что он лучше проходит в эритроциты, чем цитрат натрия. Наблюдения временных ходов концентрации глюкозы и рН для каждой пробы проводились параллельно.
Дополнительно через час после начала наблюдений и по окончании эксперимента (через 3 часа после взятия крови из организма) измерялась величина неспецифической проницаемости мембран для кислорода с помощью прибора КИНОКС каждой пробы.
Временные ходы концентрации глюкозы приведены на рис. 20 (а, б, в, г).
а)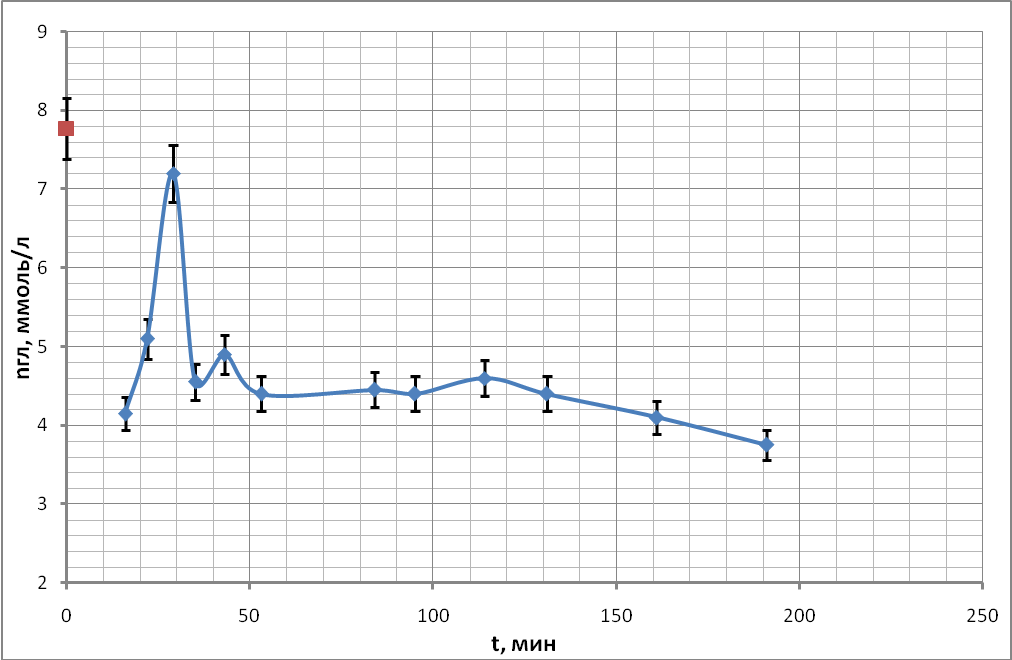
б)
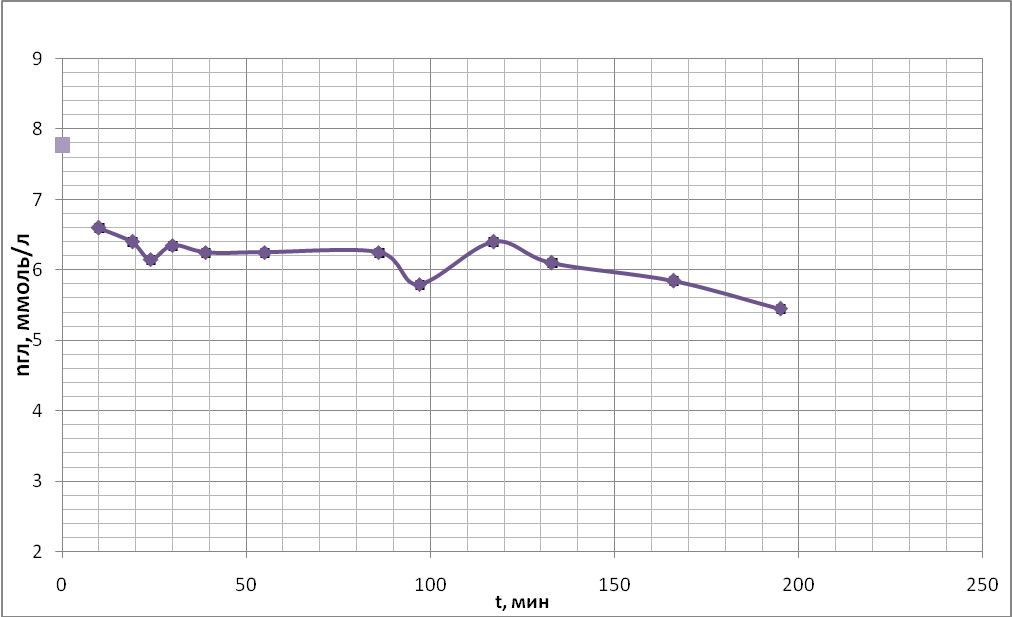
в)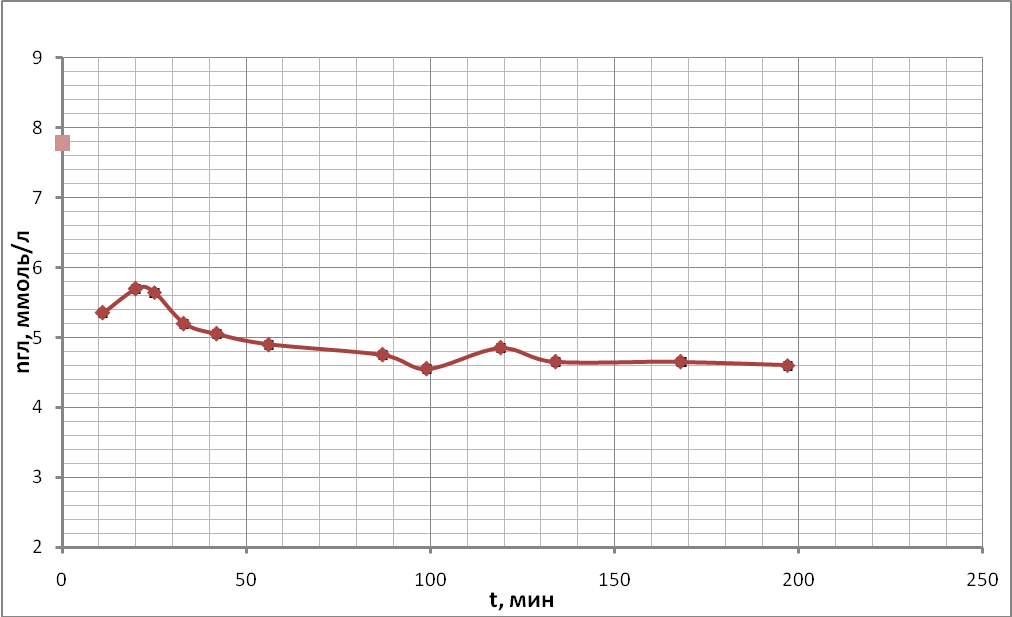
г)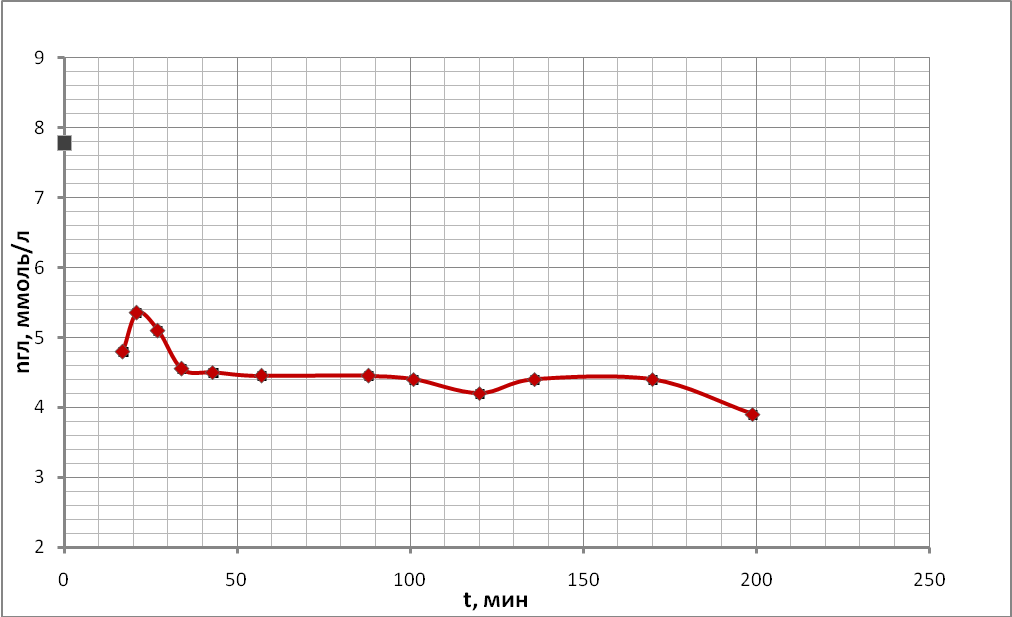

Характер хода кривых концентрации глюкозы в данной серии экспериментов существенным образом не отличался от ходов кривых, полученных в предыдущих опытах, но есть отличия в мелких деталях.
На рис. 21 представлены временные ходы показателей кислотности рН для всех проб.
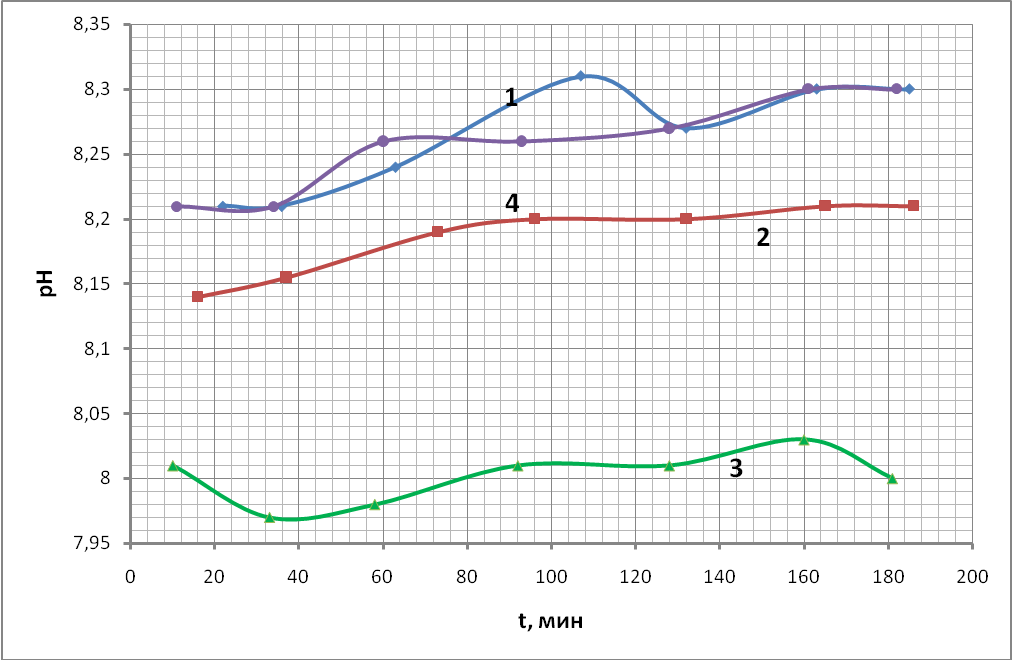

Наиболее значимыми являются начальные точки, полученные через 10-20 минут после взятия крови из организма. За это время произошло подкисление плазмы за счет выхода ионов Н+ из эритроцитов (кислые продукты гликолиза, накапливаемые в цитоплазме, выходят в плазму крови). С другой стороны, происходит подщелачивание плазмы крови за счет распада бикарбонатов и выхода образующегося СО2 в атмосферу, поскольку специальных мер для задержки его мы не предпринимали. Перемешивание крови в пробах производилось с одинаковой интенсивностью. Видно, что начальные точки значений рН для пробы крови без добавления антипептидаз и с добавлением спирта практически совпадают. Начальные точки для проб с добавлением антипептидаз существенно ниже по рН, чем начальные точки, полученные для остальных двух проб (1 и 4). Этот факт свидетельствует о том, что переход уже совершился в 1-й и 4-й пробе крови. Такой переход во второй и третьей пробе не произошел, так как состояние метаболизма эритроцитов в этих пробах наиболее близко к in vivo (за счет замедления неравновесных осмотических процессов). Это соответствует предположению, сделанному в теоретическом анализе, о том, что рН нового состояния метаболизма эритроцитов выше, чем в состоянии in vivo.
Еще до измерения
проницаемости было замечено, что при
перемешивании крови для предотвращения
оседания эритроцитов, кровь в 1-й и 4-й
пробе быстрее насыщалась кислородом,
чем во 2-й и 3-й пробе. Об этом мы судили
по цвету крови в пробирках: в 1-й и 4-й –
алая, во 2-й и 3-й – темная. На фотографии,
сделанной в день эксперимента, это
хорошо было видно (см.фото1).


Последующее определение проницаемости мембран подтвердило, что величина проницаемости мембран эритроцитов в пробах 1, 4 значительно выше, нежели чем во второй и третьей.
На рис. 22 представлен временной ход величины степени насыщения крови кислородом для 3-х проб крови (2,3,4). Номера кривых соответствуют номерам проб крови.
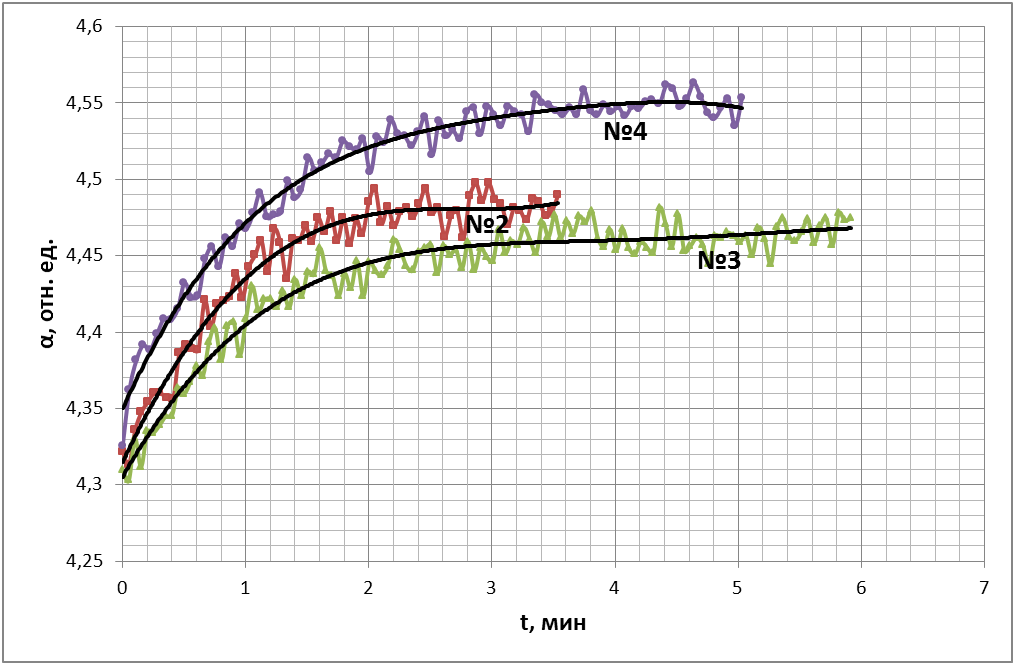

Как известно из работ Фока М.В. [17, 18] по мере насыщения крови кислородом на заключительном этапе процесса оксигенации проницаемость мембран эритроцитов резко уменьшается. Сравнение степени насыщения проб крови было затруднено тем обстоятельством, что 1 и 4 пробы были уже в значительной мере насыщены кислородом. В то время как кровь во 2-й и 3-й пробе была практически не оксигенирована. Поэтому сравнение проницаемости проводилось при одной и той же степени насыщения крови кислородом, что и представлено на рис. 22. Временной ход степени насыщения первой пробы кислородом не представлен на данном графике, так как эта проба крови находилась в насыщении кислородом, и уровень сигнала находился на более высоком уровне, что препятствовало сравнению. Значимы начальные участки кривых, чем больше угол наклона кривой к горизонтальной оси, тем выше проницаемость мембран для кислорода.
Результат сравнения говорит о том, что проницаемость мембран эритроцитов 1-й и 4-й пробы выше, нежели чем для 2-й и 3-й. Совокупность результатов в этой серии экспериментов позволяет нам утверждать, что переход состояния метаболизма эритроцитов в пробах 1-й и 4-й уже произошел, и мы наблюдали следствия этого перехода. Состояния метаболизма эритроцитов пробы 2-й и 3-й практически соответствует состоянию in vivo. Интенсивность энерговыделения при утилизации глюкозы эритроцитами в 1-й и 4-й пробе ниже и, следовательно, подкисления крови существенно ниже, чем в пробах 2 и 3. Значит, в 1-й и 4-й пробе метаболизму эритроцитов соответствует состояние с низкой концентрацией АТФ, а метаболизму в пробах 2 и 3 – состояние с высокой концентрацией АТФ, как и было предсказано теоретическим анализом.