

МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РОССИЙСКОЙ
ФЕДЕРАЦИИ
ФГОУ ВПО БЕЛГОРОДСКАЯ ГОСУДАРСТВЕННАЯ
СЕЛЬСКОХОЗЯЙСТВЕННАЯ АКАДЕМИЯ
ШМАЙЛОВ В.В.
КОНСПЕКТЫ ЛЕКЦИЙ ПО ГЕНЕТИКЕ
СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ
Белгород 2006
УДК 575 (075.8)
В.В.Шмайлов. Конспекты лекций по генетике сельскохозяйственных животных. Методическое пособие, 2006. - 106 с. / Изд.-во БГСХА.
В пособии в краткой форме изложены основные положения общей генетики, генетики сельскохозяйственных животных и ветеринарной генетики. Оно включает 15 лекций раскрывающих историю возникновения этой науки, закономерности наследования признаков при половом размножении, цитологические и молекулярные основы наследственности, теоретические основы мутационной изменчивости и генетики популяций. Рассмотрены также вопросы иммуногенетики, генетики иммунитета, наследственные аномалии животных, болезни с наследственной обусловленностью и возможность селекции на устойчивость к ним.
Пособие предназначено для студентов факультетов технологии животноводства и ветеринарной медицины.
Рецензенты:
© Белгородская государственная сельскохозяйственная академия
Лекция 1.
ПРЕДМЕТ, МЕТОДЫ И ИСТОРИЯ РАЗВИТИЯ ГЕНЕТИКИ.
ЗНАЧЕНИЕ ГЕНЕТИКИ ДЛЯ ПРАКТИКИ.
Генетика, как биологическая наука. Связь генетики с другими науками.
Биология – это наука о живых организмах. По мере своего развития биология накопила очень много информации. Всю эту массу научной информации не в состоянии осмыслить и проанализировать один исследователь. Поэтому возникла необходимость в дифференциации этой науки. Так из биологии выделились ботаника (наука о растительных организмах), зоология (наука о животных организмах), микробиология и другие науки. В том числе из биологии выделилась и генетика.
Генетика – наука о наследственности и изменчивости живых организмов. Свое название получила от греческого слова genesis (происхождение). Датой рождения генетики считается 1900 год, когда независимо друг от друга три ученых Г. Де-Фриз, К. Корренс и Э. Чермак повторно открыли законы, установленные Г. Менделем в 1865 году. В настоящее время генетика занимает центральное место в биологии.
Наследственность – свойство живых организмов обеспечивать материальную и функциональную преемственность между поколениями, а также обуславливать специфический характер индивидуального развития организмов. Каждый вид животных и растений сохраняет в ряду поколений характерные для него черты: курица выводит цыплят, овца рождает ягнят, рожь воспроизводит рожь и т.д., причем каждый вид животных и растений, куда бы его не перевозили и в какие бы условия его не помещали, если он сохранит способность размножаться, воспроизведет свои особенности. Некоторые виды могут оставаться в течение миллионов лет относительно неизменными. Например, современный опоссум мало чем отличается от опоссума раннего мелового периода.
Наряду с явлением наследственности в предмет исследования генетики входит изучение изменчивости. Изменчивость – это различие между особями одного и того же вида, между предками и потомками по ряду признаков и свойств. Если мы внимательно проанализируем стадо коров черно-пестрой породы, то при общем сходстве животных этой породы мы обнаружим различия между ними по массе, форме и расположению пятен, форме рогов, развитию вымени, темпераменту и другим признакам. Среди животных нет двух организмов, полностью похожих друг на друга, за исключением однояйцовых близнецов.
Как и любая наука, генетика не может развиваться самостоятельно, вне связи с другими науками. Она постоянно заимствует знания и достижения других наук. В первую очередь необходимо отметить тесную связь генетики с эволюционным учением Ч. Дарвина, неотъемлемой частью которого она является. Основными критериями эволюции являются: измен-
3
чивость, наследственность и естественный отбор. Генетика также изучает эти явления и помогает понять и объяснить с научной точки зрения многие вопросы эволюции.
Значительное влияние на развитие генетики оказала цитология – наука о строении клетки. Без глубоких знаний цитологии невозможно понять материальную преемственность между поколениями. Как установлено, в клетке за наследственную информацию отвечают хромосомы. Именно эти органоиды клетки в большей степени интересуют генетиков.
Генетика также связана с биохимией, так как без знания химической природы гена невозможно представить процессы передачи наследственной информации и целенаправленного вмешательства в эти процессы. Общий раздел генетики и биохимии – это раздел нуклеиновые кислоты.
Использование в качестве объекта исследований вирусов и бактерий обусловило тесную связь генетику с микробиологией и вирусологией. В частности развитие генетической инженерии это и есть удачное соединение знаний и достижений этих наук.
Генетика в своих исследованиях широко использует математические методы, в первую очередь теорию вероятности и вариационную статистику. Впервые статистический метод удачно применил для выяснения закономерностей наследования признаков Г. Мендель. Особенно широко математические методы исследований применяют в настоящее время для изучения наследования хозяйственно – полезных признаков у животных, что привело к возникновению биометрии.
Теоретические знания, накопленные в процессе развития генетики, находят практическое применение. Эти знания используют селекционеры при создании новых сортов растений и пород животных. Таким образом, генетика связана с селекцией, разведением животных и племенным делом.
Методы генетики. Для познания закономерностей наследования признаков и их изменчивости генетика использует ряд методов. Основным методом является гибридологический. При этом методе для выявления закономерностей наследования того или иного признака проводится скрещивание особей, различающихся по этому признаку, и изучается полученное потомство в первом и последующих поколениях. Гибридологический метод впервые в своих исследованиях удачно применил Г. Мендель.
Генеалогический метод является одним из вариантов гибридологического. Наследование признака при этом изучают путем анализа передачи его потомству в целых семьях или родственных группах животных, для чего составляют родословные на несколько поколений предков, отдельных особей и целых семей. Генеалогический метод имеет большое значение при изучении наследственности человека и медленно плодящихся животных, к которым обычный гибридологический метод или не применим, или требует продолжительного времени для получения результатов опыта.
Цитологический метод применяют при изучении наследственности на уровне клетки и хромосом. Установлено, что многие дефекты и нарушения
в организме связаны с изменениями в числе и структуре хромосом. Поэтому при диагностике некоторых наследственных заболеваний человека и животных широко используется цитологический метод.
Биохимический метод используется в генетике для более глубокого анализа нарушений в обмене веществ и их строении. Этот метод используется при манипуляциях на уровне ДНК в генной инженерии.
Популяционно-статический метод применяется при обработке результатов скрещиваний, изучении изменчивости признаков и связи между ними. При использовании этого метода анализу подвергаются большие по численности массивы растительных или животных организмов. Этот метод является основным в биометрии.
Феногенетический метод применяется для установления степени влияния генов и факторов внешней среды на развитие признаков организма. При использовании этого метода изучаются особи с разной наследственностью или находящиеся в разных условиях среды.
Кроме названных методов, в генетике используются и другие методы: иммунологический, близнецовый, онтогенетический.
История развития генетики. Русские ученые-генетики. Мыслители и ученые задумывались над вопросами передачи наследственных признаков от родителей к детям со времен глубокой древности. Но в те далекие времена представления о наследственности и изменчивости были очень неточными и во многих случаях ошибочными. Вот как объяснял наследование признаков у человека древнегреческий ученый Эмпедокл: «Образование зародыша подчиняется во время беременности воображению женщин: часто они воспламеняются любовью к статуям или картинам и имеют детей, похожих на эти предметы».
Многочисленные исследования по гибридизации растений, проводившиеся в 18 и 19–х веках, постепенно вскрыли отдельные закономерности в наследовании признаков. Известный шведский ученый Карл Линней, создатель системы растительного и животного мира, занимался гибридизацией растений. Линней выдвинул теорию о наследовании материнских и отцовских признаков, пологая, что у растений и животных внутренние части и органы наследуются от матери, наружные – от отца.
В 1760-70 годах ботаник Кельрейтер в результате опытов по гибридизации табака установил, что гибриды имели признаки, промежуточные между признаками обоих родителей. Это свидетельствовало о передаче родительских признаков как через пыльцу, так и через семяпочки. Кельрейтер первым установил явление, связанное с более мощным развитием гибридов первого поколения (явление гетерозиса). Однако Кельрейтеру и ученым, работающим по гибридизации растений после него, не удалось раскрыть природу механизма наследственности. Это объясняется тем, что в то время еще не были известны цитологические основы наследственности.
Большой вклад в развитие генетики внесли Томас Найт, Огюстен Сарже, Шарль Ноден и другие.
Интересовали проблемы наследования признаков и Ч. Дарвина. Он сформировал свои взгляды на эту проблему в «гипотезе пангенезиса». Согласно этой гипотезе от каждой части тела отделяются особые частицы-геммулы. Эти частицы кровью переносятся к половым клеткам. В дальнейшем при развитии нового организма из каждой частицы формируется тот орган, к которому она принадлежала в родительском организме. В этой гипотезе правильным является факт передачи признаков через половые клетки, но в то же время ошибочно предположение о связи частей тела с половыми клетками посредством особых частиц – «геммул».
Известный немецкий ботаник Карл Негели предложил умозрительную гипотезу идиоплазмы. Основными положениями ее являются существование особой субстанции в клетке – идиоплазмы, которая играет роль носительницы наследственности, признание полной равноценности всех клеток организма в явлениях наследственности и допущение возможности наследования приобретенных свойств.
Значение умозрительных гипотез наследственности состояло прежде всего в том, что они поставили ряд вопросов, которые позже стали предметом экспериментальных исследований. Эти гипотезы внесли в науку несколько новых представлений, прежде всего, допущение существования особых носителей наследственных свойств – генов, которые кодируют информацию о признаках организма.
Впервые закономерности наследования признаков в полном объеме были открыты в 1865 году Г. Менделем, который на основании опытов по скрещиванию различных сортов гороха установил единообразие гибридов первого поколения, расщепление признаков в соотношении 3 : 1 во втором поколении и независимость наследования различных признаков. Эти открытия дали толчок к дальнейшим работам по проверке описанных закономерностей на других видах растительных и животных организмов. В результате была подтверждена их универсальность, и они приобрели статус законов.
В 1910 году Томас Морган с учениками, использовав качестве объекта исследований муху дрозофилу и опираясь на накопленные к тому времени данные цитологии, создали подтвержденную в дальнейшем цитологически хромосомную теорию наследственности. Согласно этой теории, гены локализуются в хромосомах в строго определенной для каждого из них линейной последовательности и на определенном расстоянии друг от друга.
С начала 40 годов начались интенсивные исследования явлений наследственности и изменчивости на молекулярном уровне. В 1944 году американский ученый О. Эвери с сотрудниками показал, что ведущая роль в сохранении и передаче наследственной информации принадлежит ДНК. Это открытие послужило началом развития молекулярной генетики.
Дж. Уотсон и Ф. Крик в 1953 году расшифровали структуру молекулы ДНК. После этого стало ясно, каким способом кодируется наследственная информация о составе и структуре организмов. В дальнейшем, благодаря
научным работам Ниренберга и Очоа, был расшифрован генетический код. В 1969 году в США Корана с сотрудниками вне организма химическим путем синтезировал участок молекулы ДНК или простейший ген. Эта и другие работы легли в основу генной инженерии, которая бурно развивается в настоящее время.
Большой вклад в развитие мировой науки внесла наша отечественная генетика. Ученые нашей страны открыли ряд важнейших закономерностей наследственности и изменчивости.
Ю. А. Филипченко является создателем первой в России кафедры генетики в Петербургском университете. Им написано более десятка книг и брошюр по вопросам генетики.
Н. И. Вавилов провел большую по объему экспериментальную работу. Он организовал и осуществил более 10 экспедиций в малодоступные районы зарубежных стран по изучению центров происхождения культурных растений. Им написано 8 книг, создан Всесоюзный институт растений (ВИР) с широчайшей сетью отделений и опытных станций. Н. И. Вавилов был организатором и первым руководителем ВАСХНИЛ и института генетики АН СССР. Начав с экспериментальной работы в области генетики пшеницы и иммунитета растений. Н. И. Вавилов вскоре перешел к широкому изучению и обобщению собранных материалов по всем культурным растениям, что позволило ему открыть закон гомологичных рядов наследственной изменчивости. Н. И. Вавилов был талантливым организатором. В созданный им институт он приглашал крупнейших генетиков из зарубежных стран. Так, в этом институте работали ученые из США – К. Бриджес и Г. Меллер, болгарский ученый Д. Костов и другие.
Н. К. Кольцов – основоположник экспериментальной биологии. Он был блестящим организатором науки, сплотившим вокруг себя большое количество учеников, многие из которых впоследствии стали крупными учеными (А. С. Серебровский, С. С. Четвериков, Б. Л. Астауров и другие).
Г. А. Надсон совместно с Г. С. Филипповым в 1925 году провели исследования по изучению действия рентгеновских лучей на дрожжевые грибки. В их работе доказана возможность экспериментального получения мутантов под действием ионизирующих излучений. Эти работы оказали влияние на развитие и возникновение нового направления в генетике – радиационной генетики.
Г. Д. Карпаченко известен по работам в области отдаленной гибридизации. Используя явление полиплоидии, ему впервые удалось получить межвидовые гибриды растений, которые обычным путем не скрещиваются. Эти теоретические разработки сегодня с успехом используют селекционеры в своей работе.
Большой вклад в развитие отечественной генетике внесли также М. Е. Лобашев, Н. П. Дубинин, Н. В. Цицин, В. В. Сахаров и другие.
Значение генетики для практики. Генетика сегодня занимает ведущее место в современной биологии. Фундаментальные открытия этой науки реализуются в селекции растений и разведении животных. За последние
годы созданы гибриды ячменя и пшеницы, ячменя и ржи, выведены новые сорта пшеницы, способные давать около 100 центнеров зерна с гектара, высокомасличные сорта подсолнечника с содержанием жира в семенах до 55%. Выведены фитофтороустойчивые и ракоустойчивые сорта картофеля, полиплоидные сорта сахарной свеклы и плодовых деревьев. В животноводстве широко используется явление гетерозиса (более мощное развитие гибридов первого поколения по сравнению со своими родителями). Практически на всех птицефабриках нашей страны производят мясо птицы за счет бройлеров, а для получения яиц используют гибридную птицу. Применяется это явление также в свиноводстве и мясном скотоводстве.
С помощью иммуногенетических методов сегодня проводится уточнение происхождения животных при их продаже.
Разработанные методы пересадки оплодотворенных яйцеклеток и эмбрионов нашли применение при размножении высокопродуктивных животных.
Методы генетической инженерии широко применяются в биотехнологии (отрасли по производству нужных человеку веществ с помощью живых организмов). Методами генетической инженерии созданы промышленные штаммы микроорганизмов, продуцирующие инсулин (гормон щитовидной железы), интерферон, соматотропин и другие биологически активные вещества. В медицине и ветеринарии нашли применение моноклональные антитела, полученные методами гибридомной технологии.
Генетические методы находят применение в медицине для ранней диагностики некоторых наследственных заболеваний, защиты организма человека от негативного действия различных факторов и веществ.
Лекция 2
ЦИТОЛОГИЧЕСКИЕ ОСНОВЫ НАСЛЕДСТВЕННОСТИ
Строение клетки. После того как был сконструирован микроскоп учёные установили, что все организмы растений и животных состоят из мельчайших частиц- клеток. Клетка является элементарной единицей строение всех живых организмов. Данные микроскопических исследований показывают, что каждая клетка содержит много органоидов которые выполняют разнообразные функции. Схема строения животной клетки показана на рисунке 1.
Снаружи клетка покрыта оболочкой или мембраной. Клеточная оболочка состоит из трёх слоёв: белки, липиды, белки, а у растительных клеток есть ещё слой из клетчатки. В оболочке клеток имеются поры, через которые осуществляется связь клетки с окружающей средой. Установлено, что клеточные мембраны обладают дифференцированной проницательностью, одни вещества пропускают, другие - нет. Клеточные мембраны придают клетке определённую форму, выполняют защитную, питательную и выделительную функции. Наиболее характерным процессом для мембран является активный транспорт веществ. Этот транспорт осуществляется с помощью белков, которой переносят различные вещества с одной стороны мембраны на другую. Под оболочкой клетки находиться гелеобразное вещество – цитоплазма, в которой располагаются органоиды: ядро, эндоплазматическая сеть, рибосомы, митохондрии, комплекс Гольджи, лизосомы, клеточный центр, а в клетках растений – также и пластиды.
В большинстве случаев ядро имеет шаровидную или овальную форму и располагается в центре клетки. Оно служит важным регулирующим центром клетки, так как в нём расположены хромосомы, определяющие признаки данного организма, и управляет многими внутриклеточными процессами. В ядре находятся сферические тельца, называемые ядрышками. Они исчезают, когда клетка готовится к делению. Предполагают, что ядрышки участвуют в синтезе рибонуклеиновых кислот.
Внутреннюю часть клетки заполняет гелеобразное вещество, которое называется цитоплазмой. Цитоплазма представляет собой сложный лабиринт из мембран, образующих эндоплазматическую сеть. Существуют два типа эндоплазматической сети: гранулярная и агранулярная. На поверхности гранулярной сети находится много рибонуклеопротеидных частиц, называемых рибосомами. Эндоплазматическая сеть увеличивает обменную поверхность клетки. По её канальцам происходит транспорт веществ внутри клетки.
Рибосомы – мелкие частицы, находящиеся в клетке как в свободном состоянии, так и прикрепленные к наружной поверхности каналов эндоплазматической сети. Они являются чистыми рибонуклеопротеидами, так как состоят только из РНК и белка. В рибосомах бактерий содержится 60-64% РНК, в рибосомах млекопитающих – 40-45%.
Каждая рибосома состоит из двух сферических субъединиц, неравных по величине и химическому составу. В рибосомах происходит биосинтез белка. Количество рибосом в клетке непостоянно и зависит от интенсивности синтеза белка.
Все живые клетки содержат митохондрии. Это небольшие тельца размером 0,2 – 5 мкм, имеющие сферическую или палочковидную форму. В клетке их может быть от несколько штук до тысячи и более. Митохондрии выполняют энергетическую функцию, поэтому они сосредоточены в той части клетки где обмен веществ наиболее интенсивен. В них синтезируется аденозинтрифосфорная кислота (АТФ), необходимая для энергетических затрат клетки.
Митохондрии имеют внутреннюю внешнюю мембраны. Внутренняя мембрана образует складки, называемые кристами. В химическом отношении митохондрии включают в себя липопротеидный комплекс. В их составе содержится также много дыхательных ферментов: оксидаза, цитохромоксидаза и др.
В составе почти всех клеток имеется комплекс Гольджи. Он представляет собой сеть канальцев, выстланных мембранами. Обычно он расположен около ядра и окружает центриоли. Комплекс Гольджи служит местом временного хранения веществ, вырабатываемых в гранулярной эндоплазматической сети.
Лизосомы – группа внутриклеточных органелл, встречающихся в клетках животных. Они представляют собой ограниченные мембранами тельца, которые содержат разнообразные ферменты, способные гидролизовать макромолекулярные компоненты клетки.
Для цитоплазмы растительных клеток характерно присутствие пластид, которые осуществляют фотосинтез, синтез крахмала и пигментов. По окраске и выполняемой функции пластиды разделяются на три группы: лейкопласты, хлоропласты и хромопласты. Пластиды размножаются путём прямого деления. Для генетиков пластиды представляют объект тщательного изучения, так как они содержат ДНК и принимают участие в передаче наследственной информации.
Строение и типы хромосом. Главными органоидами клетки, локализованными в ядре и отвечающими за хранение и передачу наследственной информации, являются хромосомы. Своё название эти органоиды получили от греческого слова хром, что в переводе означает цвет. Это указывает на интенсивное поглощение хромосомами красителей. Хромосомы можно видеть под микроскопом только в период деления клетки. Наиболее удачное время для наблюдения за хромосомами это метафаза. У большинства организмов хромосомы имеют продолговатую форму и длину от 1 до 30 мкм.
При микроскопическом анализе хромосом видны различия в их форме и величине. Каждая хромосома имеет своё индивидуальное строение. Вместе с тем можно заметить, что хромосомы имеют общие морфологические признаки. Схема строения хромосомы показана на рисунке.
Хромосома имеет продолговатые участки – плечи или теломеры, которые разделены центромерой. Некоторые хромосомы имеют вторичную перетяжку, которая отделяет от хромосомы небольшой участок, называемый спутником. По положению центромеры хромосомы разделяют на четыре типа: метацентрики, субметацентрики, акроцентрики и телоцентрики.
К метацентрикам относят хромосомы, у которых центромера расположена по середине. Хромосомы, у которых одно плечо значительно длиннее другого, называют субметацентриками. К акроцентрическому типу относят хромосомы, у которых одно плечо длинное, а другое представлено небольшим зачатком. Телоцентрические хромосомы имеют только одно плечо. Точно определить тип хромосомы можно по величине плечевого индекса, который вычисляют путём деления длины длинного плеча на короткое.
Длинное плечо
Плечевой индекс ( П.И.) = ----------------------
Короткое плечо
К метацентрикам относят хромосомы с величиной плечевого индекса
1 - 1,9, к субметацентрикам – 2 – 4,9, к акроцентрикам – 5 и более. Для телоцентрических хромосом плечевой индекс не вычисляют, так как они имеют только одно плечо. Расположение центромеры и величина плечевого индекса служат одним из критериев классификации и идентификации хромосом.
Химический анализ хромосом показал, что в их состав входят белки сложного состава типа гистонов и протаминов и ДНК. Причём ДНК в составе хромосом находится в спирализованном состоянии в виде хроматид.
При окрашивании хромосом установлено, что они окрашиваются по всей длине не одинаково. Наблюдаются светлые и тёмные участки. Тёмно-окрашенные участки хромосом были названы гетерохроматиновыми, а светлоокрашенные – эухроматиновыми. Предполагают, что тёмноокрашенные участки – это неактивные участки где ДНК плотно спирализована.
В настоящее время разработано много методов дифференцированного окрашивания хромосом: G, C, Q, NOR и др. При дифференцированном окрашивании каждая хромосома приобретает свой специфический рисунок – чередование светлых и тёмных полос, отражающих различную функциональную активность отдельных участков хромосом. С помощью дифференцированного окрашивания можно не только идентифицировать хромосомы, но и обнаруживать различные нарушения в их строении.
В соматических клетках всех организмов содержится двойной или диплоидный набор хромосом (2n). Половые же клетки имеют одинарный или гаплоидный набор хромосом (n). Одинарный набор хромосом называется геномом.
Набор хромосом соматической клетки, свойственный данному виду организмов, называют кариотипом. Причём кариотип характеризуется не только числом хромосом, но и их формой, наличием полос при дифферен-
цированном окрашивании и другими признаками.
Кариотипы человека и других организмов
Человек 46 Кролик 44
Крупный рогатый скот 60 Осёл 62
Лошадь 64 Кошка 36
Свинья 38 Куры 78 (77)
Овца 54 Гуси 82 (81)
Собака 78 Дрозофила 8
Среди всех хромосом различают пары аутосом, одинаковых для мужских и женских особей, и одну пару половых хромосом, различающихся у мужских и женских организмов. Половые хромосомы женских особей млекопитающих обозначают буквами ХХ, а мужских ХY.
Деление клеток (митоз, мейоз). В основе роста и дифференцировки органов и тканей животных лежит размножение клеток путём их деления. Основным типом деления соматических клеток является митоз. Для митоза характерно строгое распределение генетической информации в дочерне клетки.
Схема митоза выглядит следующим образом:
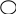 2n
- материнская клетка
2n
- материнская клетка

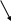


2n 2n – две дочерние клетки
Промежуток от одного клеточного деления к другому называется клеточным циклом. Клеточный цикл состоит из интерфазы и собственно митоза. В период интерфазы, а она по продолжительности во много раз длиннее митоза, клетка активно выполняет жизненные функции, типичные для неё. В интерфазе выделяют три периода: предсинтетический (G1), синтетический (S) и постсинтетический (G2). В предсинтетическом периоде в клетке происходит активный синтез белка и других веществ, необходимых для образования клеточных структур и последующего деления. В S – периоде синтезируется ДНК и происходит формирование второй хроматиды. Таким образом, в митоз клетка вступает с удвоенным числом хромосом. В постсинтетическом периоде активность жизненных процессов в клетке снижается, клетка готовится к делению.
Вслед за интерфазой начинается деление клетки – митоз. Большинство учёных митоз разделяют на четыре фазы: профаза, метафаза, анафаза и телофаза.
Профаза. Эта фаза характеризуется постепенным уплотнением и спирализацией хромосом, в результате чего они становятся различимы под микроскопом, образуя нитевидные структуры. Видно, что каждая хромосома состоит из двух копий, расположенных рядом друг с другом и соеди-
нённых центромерой. Эти копии, пока они не разошлись, называются сестринскими хроматидами. Другим характерным событием профазы являются постепенное исчезновение ядрышка и разрушение оболочки ядра. Центриоли к концу профазы обычно расходятся к полюсам клетки. Под микроскопом в эту фазу видна сетчатая структура ядра.
Метафаза. У большинства организмов в этой фазе ядерная оболочка уже исчезла и хромосомы в сформированном виде находятся в цитоплазме. Центромеры хромосом прикрепляются нитями веретена деления к центриолям клетки. В эту фазу хромосомы собираются в плоскости, расположенной в области экватора клетки. Эта фаза митоза наиболее удобна для наблюдения и изучения хромосом.
Анафаза. Обычно это короткая стадия митоза. В эту фазу каждая центромера делится пополам. В результате сокращения нитей веретена дочерние хроматиды расходятся к полюсам клетки.
Телофаза. В этой фазе два набора хромосом группируется у противоположных полюсов клетки. Здесь они начинают раскручиваться и удлиняться, приобретая форму интерфазных хромосом. Вокруг каждого набора хромосом образуется ядерная мембрана и вновь возникают ядрышки. К концу телофазы делится цитоплазма и образуются две дочерние клетки с диплоидным набором хромосом.
Продолжительность митоза зависит от типа тканей, физиологического состояния организма и внешних факторов. Например, установлено, что во время покоя и сна животных митотическая активность различных тканей значительно выше, чем в период бодрствования.
Длительность митоза может составлять от нескольких минут до часа и более. При изучении митоза в культуре клеток человека установлено, что в среднем продолжительность фаз митоза следующая: профаза длится 60% времени, метафаза – 5%, анафаза – 5% и телофаза 30%.
Таким образом, в результате митоза из одной материнской клетки возникают две дочерние, содержащие такой же набор хромосом, как у исходной клетки. Основное биологическое значение митоза состоит в точном распределении хромосом между дочерними клетками; тем самым сохраняется преемственность хромосомного набора в ряде клеточных поколений и полноценность генетической информации каждой клетки, что необходимо для осуществления общих и специфических функций живого организма.
Мейоз – это два последовательных деления ядра, которые приводят к образованию половых клеток. Во время мейоза каждая клетка делится дважды, в то время как хромосомы удваиваются лишь один раз, в результате чего число хромосом в половых клетках оказываются вдвое меньше их числа в исходной клетке. Схема мейоза выглядит следующим образом.
2n - соматическая клетка
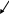
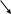 -
редукционное деление
-
редукционное деление
n n
 - эквационное деление
- эквационное деление
n n n n - половые клетки
Первое деление мейоза, которое приводит к уменьшению числа хромосом в два раза, называется редукционным, второе деление, в результате чего число хромосом не изменяется, - эквационным. Предшествующая мейозу интерфаза полностью аналогична митотической интерфазе. В ней происходит синтез ДНК и удвоение хромосом.
Редукционное деление начинается с профазы 1, которая подразделяется на пять стадий: лептонемы, зигонемы, пахинемы, диплонемы и диакинеза. В стадии лептонемы хромосомы представляют тонкие нити. Они еще деспирализованы и в 2-5 раз длиннее метафазных. Под микроскопом можно видеть, что они состоят из двух хроматид, соединенных центромерой.
На стадии зигонемы гомологичные хромосомы коньюгируют, т. е. соединяются друг с другом наподобие застёжки « молния ». Такое соединение гомологичных хромосом называется синапсом. Это важное генетическое событие, поскольку оно даёт возможность обмена участками между гомологичными хромосомами. На этой стадии под микроскопом можно видеть, что каждая хромосома состоит из двух нитей, а в комплексе образуется бивалент их четырёх хроматид. Далее на стадии пахинемы происходит утолщение хромосом, так что становятся хорошо различимы сестринские хроматиды.
На стадии диплонемы две гомологичные хромосомы почти расходятся, однако сестринские хроматиды остаются соединёнными общей центромерой. Кроме того, у гомологичных хромосом остаются одна или несколько зон контакта, которые называются хиазмами. Каждая хроматида может образовывать хиазмы с любой из хроматид гомологичной хромосомы, так что хиазмами могут быть связаны две, три или все четыре хроматиды бивалента. Число хиазм в биваленте может быть различным, но обычно не более двух - трех. Наличие хиазм свидетельствует о том, что между хроматидами происходит кроссинговер (т.е. обмен участками).
Диакенез характеризуется максимальным утолщением и спирализацией хромосом, принимающих форму коротких толстых палочек. У большинства организмов на этой стадии хиазмы перемещаются в направлении от центромер к концам хромосом и исчезают. После завершения диакенеза ядерная мембрана и ядрыщки растворяются.
После завершения профазы 1 наступает метафаза 1. В эту фазу биваленты располагают в плоскости экватора центромерами к противоположным полюсам. В анафазе 1 начинается расхождение гомологичных хромосом к противоположным полюсам, которое носит случайный характер. В телофазе 1 хромосомы достигают полюсов клетки. Вокруг них формируется ядерная оболочка и начинается деление цитоплазмы. Таким образом, в результате редукционного деления из одной клетки с диплоидным набором хромосом образуют две клетки с гаплоидным числом хромосом.
Между первой и второй стадиями мейоза имеется непродолжительный период покоя- интеркенез, во время которого не происходит синтез
ДНК и удвоения хромосом.
Эквационное деление происходит по типу митоза. Профаза 2 часто проходит очень быстро. В метафазе 2 хромосомы прикрепляются центромерами к нитям веретена и располагаются в плоскости экватора. К началу анафазы 2 каждая центромера делится и сестринские хроматиды таким образом становятся хромосомами, расходящимися затем к противоположным полюсам. Телофаза 2 завершается образованием ядерной оболочки вокруг каждого из двух гаплоидных ядер.
Таким образом, в результате двух мейотических делений из одной клетки с диплоидным набором хромосом образуются четыре клетки с гаплоидным набором. Случайный характер распределения хромосом и обмен их участками в результате кроссинговера позволяет создать новые комбинации наследственного материала в половых клетках. Кроме этого, мейоз обеспечивает поддержание постоянства числа хромосом в смежных поколених организмов.
Образование половых клеток и оплодотворение. У живых организмов размножение происходит в основном половым путём. Начало новому организму дают половые клетки – гаметы. Мужские половые клетки – сперматозоиды образуются в семенниках, а женские яйцеклетки – в яичниках. Половые клетки образуются из соматических в результате сложных процессов в мейозе. Процесс образования мужских половых клеток называется сперматогенезом, а женские – оогенезом.
Соматическая клетка, из которой образуются женские половые клетки, называется сперматоцитом первого порядка. В результате редукционного деления из него формируются два сперматоцита второго порядка с гаплоидным числом хромосом. Далее происходит эквационное деление, в результате которого каждый сперматоцит первого порядка делится с образованием двух клеток. Таким образом, в результате двух делений образуются четыре сперматида, которые в процессе формирования превращаются в полноценные сперматозоиды.
Сперматозоиды состоят из головки, шейки и хвоста. Головка содержит ядро и очень небольшое количество цитоплазмы. Сперматозоиды способны передвигаться в половых путях самки. Эти клетки образуются в придатках семенников непрерывно в течении всей жизни животного, начиная с момента полового созревания.
Яйцеклетка образуется из ооцитов первого порядка в процессе мейоза. При первом деления из ооцита образуются две гаплоидные клетки – ооцит второго порядка и полярное тельце. Причём эти клетки не равноценны. Ооцит второго порядка намного крупнее, так как эта клетка содержит почти всю цитоплазму материнской клетки. Полярное тельце включает хромосомы и очень небольшое количество цитоплазмы. Далее ооцит второго порядка делится, образуя крупную клетку, - оотид и полярное тельце. Полярное тельце полученное при первом делении также делится. Оотид в процессе формирования и созревания превращается в полноценную яйцеклетку, а полярные тельца в дальнейшем развитии не участвуют.
Таким образом, в результате двух последовательных делений из ооцита первого порядка образуется одна полноценная яйцеклетка с гаплоидным набором хромосом и три полярных тельца. Образование яйцеклеток у самок животных происходит при их половом созревании и протекает циклично.
Половые клетки участвуют в процессе оплодотворения. Оплодотворение – это слияние мужских и женских половых клеток, в результате чего восстанавливается диплоидный набор хромосом и начинается развитие нового организма.
Процесс оплодотворения видоспецифичный или избирательный. Это значит, что в норме яйцеклетка оплодотворяется сперматозоидом своего вида. Это закреплено процессом эволюции и не допускает смешения видов в природе. Кроме этого, оплодотворение носит случайный характер, т. е. яйцеклетка может быть оплодотворена любым из попавших в половые пути самки сперматозоидом.
Как исключение, в природе иногда наблюдается развитие организмов без оплодотворения. Это явление получило название партеногенеза. При партеногенезе получают потомство, полностью похожим на родительский организм. Различают две формы партеногенеза – андрогенез и гиногенез. При андрогенезе получают особей только мужского пола, при гиногенезе – женского. Используя явление партеногенеза Астаурову удалось решить проблему регуляции пола у тутового шелкопряда. В природе партеногенез встречается у низших форм (ракообразные, перепончатокрылые и др.), а из высших это явление обнаружено у птиц (индейки).
Лекция 3
Закономерности наследования признаков при половом размножении
Вопрос наследования признаков у различных видов растительных и животных организмов интересовал ученых и практиков с давних времен. В многочисленных работах гибридизаторы 18-го и первой половины 19-го веков наметили некоторые закономерности наследования признаков. Но ни эксперименты, ни чисто умозрительные теории наследственности (Дарвин, Негели, Вейсман и др.) не дали исчерпывающего объяснения и обоснования установленных фактов. Приоритет открытия законов наследования признаков принадлежит Г.Менделю. Результаты своих опытов Г.Мендель опубликовал в 1865 году, и впоследствии они стали известны в научном мире.
Открытие законов наследования признаков стало возможным благодаря использованию Г.Менделем метода гибридологического анализа. Сущность этого метода заключается в следующем:
- для скрещивания подбираются особи, различающиеся между собой по одной, двум или более парам альтернативных (взаимоисключающих) признаков, например, красная и белая окраска цветков и др.
- учет и анализ наследования признаков проводится на протяжении нескольких поколений.
- при скрещивании проводится точный количественный учет проявления у потомков изучаемых признаков.
При гибридологическом анализе для записи различных схем скрещиваний используются условные обозначения: родителей обозначают буквой Р (от латинского слова parentеs - родители), женский пол знаком -♀ , мужской - ♂ , скрещивание - х, потомство буквой F (от латинского fillii- дети ).
Согласно закону о чистоте гамет в зиготе и соматических клетках организма имеется по два аллеля от каждой пары, один из которых получен от отца, другой - от матери. В отличие от них половые клетки - гаметы содержат только по одному гену из каждой аллельной пары.
В генетических схемах скрещиваний аллельные гены принято обозначать одной и той же буквой. Доминантные гены обозначаются заглавными буквами латинского алфавита (А,В,С), а их рецессивные аллели - строчными (а, в, с).
Наследование признаков при моногибридном скрещивании. Моногибридным называется скрещивание организмов, отличающихся одной парой альтернативных признаков. Г.Мендель изучал у гороха семь пар четко выраженных альтернативных признаков (окраска цветков и семян, форма семян, величина растений и др.). Сначала Мендель скрещивал два растения, отличающихся по изучаемому признаку, высевал полученные в результате скрещивания семена и изучал внешний вид гибридов первого поколения. При этом он обнаружил, что гибридные растения сходны между собой и напоминают одну из родительских форм. Если, например, у одного родителя цветки были красными, а у другого белые, то все гибридные растения первого поколения были красными. Признак, проявившийся у гибридов первого поколения был назван доминантным (преобладающим), а исчезнувший признак – рецессивным. Сходные результаты были получены и по другим признакам.
Продемонстрируем это правило на результатах скрещивания крупного рогатого скота черной и красной масти.
-
Ген
Признак
А
а
черная масть
красная масть
Р ♀ АА х ♂ аа
черная красный


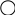 Гаметы: А
а
Гаметы: А
а
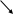
F1 Аа
черная
На основании скрещивания и анализа гибридов первого поколения Г.Менделем был сформулирован первый закон (правило) - единообразия гибридов первого поколения. Этот закон в настоящее время читается так: " При скрещивании двух гомозиготных особей, отличающихся по одной паре альтернативных признаков, первое поколение будет единообразно по данному признаку".
Следующим этапом было скрещивание двух гибридов первого поколения между собой. Из полученных семян Мендель выращивал растения второго поколения и проводил учет проявившихся признаков. Оказалось, что какая бы пара признаков не изучалась, 3/4 растений второго поколения имели доминантный признак и 1/4 – рецессивный. Такое соотношение между доминантным и рецессивным признаками ( 3 : 1 ) было впоследствии подтверждено другими исследователями. Схема скрещивания животных черной масти, полученных в первом поколении, приводится ниже.
Р F1 ♀ Аа х ♂ Аа
черная черный
Гаметы: А а А а
Для записи потомства второго поколения удобно пользоваться решеткой Пеннета в которой гаметы одного родителя размещают по вертикали, а другого – по горизонтали.
F2
-
 ♂
♂
♀
р А
q a
р А
АA
черная
Aa
черная
q a
Aa
черная
Aa
красная
В основе этих расщеплений лежат известные теперь биологические закономерности: нахождения генов в хромосомах, редукционного деления, приводящего к гаплоидному набору хромосом в гамете, парность
хромосом в зиготе, и, наконец, случайный характер оплодотворения.
Появление рецессивных признаков у растений второго поколения ясно показывает, что эти признаки не исчезают и не смешиваются с доминантными у гибридов. Наследственные факторы или гены имеют дискретный характер и передаются из поколения в поколение в "чистом" виде.
Полученные во втором поколении результаты Мендель сформулировал в виде правила расщепления, которое читается так: «При скрещивании гибридов первого поколения между содой в потомстве F2 происходит расщепление в соотношении 3:1».
Расщепление гибридов первого поколения в соотношении 3:1 будет соблюдаться при следующих условиях; равновероятное образование гамет двух типов гибридными организмами, равновероятная встреча гамет обоих родителей при оплодотворении, одинаковая жизнеспособность особей различных генотипов во втором поколении, большое количество учтенного потомства. Если же какое-либо условие не соблюдается, соотношение организмов во втором поколении будет иным.
Анализ потомства F2 показал, что при скрещивании животных черной масти между собой одна часть их не дает расщепления (СС), а другие две части (Сс) дают. Для того что6ы различать эти организмы, были введены термины гомозиготность и гетерозиготность.
Гомозиготными называют организмы, имеющие одинаковые наследственные факторы или гены (АА,аа) и не дающие расщепления при скрещивании. Гетерозиготные - это организмы с разными наследственными факторами ( А а ) и дющие расщепление при скрещивании.
В 1909 году датским ученым В. Иоганнсеном были введены термины генотип и фенотип. Генотип - совокупность наследственных задатков (генов) организма. Фенотипом называют совокупность всех признаков и свойств организма, доступных наблюдению и анализу. Фенотип формируется под влиянием генотипа и условий среды.
Таким образом, во втором поколении расщепление по фенотипу 3 : 1 и по генотипу - 1 : 2 : 1.
Взаимодействие аллельных генов. Аллельными называются гены одной пары ( А и а ), локализованные в одном и том же локусе гомологичных хромосом. В опытах, проведенных Г. Менделем, мы описали явление доминирования, при котором один из генов подавлял действие другого гена. Но такое взаимодействие генов бывает не всегда. В дальнейших опытах учеными были выявлены случаи неполного доминирования, промежуточного наследования, кодоминирования и сверхдоминирования. При промежуточном наследовании потомство первого поколения сохраняет единообразие, но оно не похоже полностью ни на одного из родителей, как это было при полном доминировании, а обладает признаком промежуточного характера. Например, при скрещивании длинноухих овец ( длина ушной раковины около 10 см) с безухими баранами потомство первого поколения будет короткоухим (длина ушной раковины около 5 см).
Иногда признак занимает не среднее выражение, а уклоняется в сторону одного из родителей. В этом случае говорят о неполном доминировании. Примером неполного доминирования может быть наследование черно-пестрой масти у скота. При скрещивании черно-пестрых коров с быками, имеющими сплошную окраску потомство первого поколения будет иметь пеструю окраску, но интенсивность этой окраски у гибридов будет отличаться от чистопородных черно-пестрых животных.
При сверхдоминировании у гибридов первого поколения проявляется гетерозис. Гетерозисом называется явление превосходства помесей первого поколения. Например, при скрещивании кур породы А, имеющих их живую массу 3,0 кг, с петухами породы В (живая масса 3,3 кг) получили бройлеров, живая масса которых 3,5 к г.
При кодоминировании у гибридной особи в равной мере проявляются оба родительских признака. По типу кодоминирования наследуется большинство антигенов систем групп крови человека и животных и полиморфные типы белков.
Анализирующее скрещивание - это скрещивание организма неизвестного генотипа с рецессивной формой с целью определения его генотипа. Генотип родителя определяют по фенотипу потомства. Если при анализи-
рующем скрещивании получают единообразное потомство, то организм был гомозиготен. В том случае, когда в потомстве наблюдается расщепление, анализируемая особь была гетерозиготной. Продемонстрируем это на примере. Допустим, мы купили черную собаку неизвестного генотипа и решили узнать, чистопородна она или нет. Нам известно, что черная масть у собак (В) доминирует над коричневой (в). Для этого проведем анализирующее скрещивание, т.е. скрестим нашу собаку с животным, имеющим коричневую окраску, и по результатам определим ее генотип.
Родители: ВВ? х вв Вв? х вв
черная коричневая черная коричневая

 Гаметы: В в
В, в в
Гаметы: В в
В, в в
Потомство: Вв Вв вв
 черная черная коричневая
черная черная коричневая
Дигибридное скрещивание. Дигибридное скрещивание – это скрещивание особей, различающихся по двум парам альтернативных признаков. Установив закономерности наследования по отдельным парам признаков, Мендель провел опыты, где одновременно учитывал два признака: окраску семян гороха (желтые и зеленые) и их форму (гладкие или морщинистые ). Мендель скрещивал две чистые линии гороха, одна из которых образовывала желтые и гладкие, а другая - зеленые и морщинистые семена. Из предыдущих опытов было известно, что желтый цвет и гладкая форма семян являются доминантными признаками.
Опыты показали, что в первом поколении все гибридные растения имели желтые гладкие семена. Эти растения Мендель скрещивал и подсчитывал во втором поколении число растений с разными типами семян. Из 556 семян 315 оказалось желтых гладких, 101 - желтое морщинистое, 108 -зеленых гладких и 32 - зеленых морщинистых. На основании полученных данных Мендель определил статистический характер расщепления во втором поколении при дигибридном скрещивании - 9 : 3 : 3 : I .
Рассмотрим схему дигибридного скрещивания на примере животных. Возьмем для анализа два признака, определяющие масть у крупного рогатого скота и наличие рогов. Установлено, что черная окраска (А) доминирует над красной (а), а комолость (К) - над рогатостью (а). При скрещивании гомозиготного черного комолого быка с красной рогатой коровой, все потомство первого поколения будет черным комолым.
Р ♀ ААКК х ♂ аакк
черная комол. красный рогатый
Гаметы: АК ак
F1 АаКк
черная комолая
Далее посмотрим, какое потомство будет при скрещивании между собой животных первого поколения.
Р ♀ АаКк х ♂ АакКк
черная комолая черный комолый
Гаметы: АК, Ак, аК, ак АК, Ак, аК, ак
-
 ♀
♀
А К
А к
а К
а к
А К
ААКК
черн. комол.
ААКк
черн. комол.
АаКК
черн. комол.
АаКк
черн. комол.
А к
ААКк
черн. комол.
ААкк
черн. рогат.
АаКк
черн. комол.
Аакк
черн. рогат.
а К
АаКК
черн. комол.
АаКк
черн. комол.
ааКК
красн. комол.
ааКк
красн. комол.
а к
АаКк
черн. комол.
Аакк
черн. рогат.
ааКк
красн. комол.
аакк
крас. рогат.
Как видно из решетки Пеннета, во котором поколении расщепление по фенотипу : 9 черных комолых, 3 черных рогатых, 3 красных комолых и 1 красное рогатое. Причем расщепление по каждому из двух признаков не зависит друг от друга, то есть 3 : 1. Расщепление по генотипу во втором поколении более сложно. Среди потомства встречаются животные восьми различных генотипов. На основании проведенных опытов Г.Мендель сформулировал третий закон (правило) независимого наследования признаков, которое читается так: «При скрещивании организмов, различающихся по двум или более парам альтернативных признаков, каждый из них наследуется независимо друг от друга» . Следует заметить, что независимое наследование признаков бывает только в том случае, если гены этих признаков располагаются в разных хромосомах.
Полученные Менделем закономерности в последующем были подтверждены данными цитологии. Независимое комбинирование признаков связано с независимым распределением хромосом разных пар в гаметы при редукционном делении и случайным их сочетанием в результате оплодотворения.
При тригибридном скрещивании, в котором анализ проводится по трем парам признаков, первое поколение единообразно и имеет доминирующие признаки родителей. Во втором поколении расщепление носит более сложный характер, так как образуется 8 сортов гамет, которые, сочетаясь друг с другом при оплодотворении, дают 64 комбинации, включающие 8 фенотипов. Расщепление по фенотипу при полном доминировании признаков происходит во втором поколении в отношении: 27: 9 : 9 : 9 : 3 : 3 : 3 : 1.
Чем большим количеством признаков отличаются скрещиваемые особи, тем сложнее расщепление и сильнее возрастает комбинативная изменчивость. Если учесть, что у животных и человека несколько десятков хромо-
сом и в каждой хромосоме более сотни генов, то число возможных фенотипов и генотипов при независимом комбинировании будет иметь очень большие величины. Таким образом, в природе нет двух организмов, похожих друг на друга за исключением однояйцовых близнецов.
Фенотипический радикал. Остановимся несколько написания формул генотипов и фенотипов. При полном доминировании гомозиготные формы по фенотипу неотличимы от гетерозиготных. Так, организм генотипа ААВВ по фенотипу неотличим от АаВв„ ААВв и АаВВ. Сходные фенотипы иногда называют фенотипическим радикалом. Под фенотипическим радикалом понимается та часть генотипа организма, которая определяет его фенотип. Для четырех перечисленных генотипов фенотипический радикал будет А-В-. Подставляя в фенотипический радикал разные аллели на место прочерков, можно получить разные генотипы. Например, фенотипический радикал А-вв имеет генотипы ААвв и Аавв. С помощью фенотипического радикала можно написать расщепление по фенотипу в F2 при дигибридном скрещивании - 9 А-В-, 3 А-вв, 3 ааВ- и 1 аавв.
Наследование признаков при взаимодействии генов. В природе часто встречаются случаи, когда признак определяется не одним, а несколькими парами неаллельных генов. Формирование признака в этом случае зависит от характера их взаимодействия в процессе развитии организма. В настоящее время обнаружено и дано теоретическое объяснение нескольким случаям такого взаимодействия.
Эпистаз. При этом типе взаимодействия ген одной пары подавляет действие других неаллельных генов. Гены, которые подавляют действие других генов называются эпистатическими или генами - супрессорами. Эпистаз бывает' доминантным или рецессивным. При доминантном эпистазе доминантный ген подавляет действие других генов, при рецессивном - рецессивный.
Рассмотрим простейший случай наследования масти у лошадей по типу эпистаза. В данном случае масть определяется парой неаллельных генов. Причем ген С является эпистатическим и подавляет действие другого доминантного гена В. Поэтому все организмы, имеющие этот ген, независимо от других генов будут иметь серую масть.
-
Взаимодействующие
гены
Генотипы
Признак
С, В
ССBB,ССВB, CCBb, CcBb
серая
C, b
CCbb, Ccbb
серая
с, B
ccBB, ccBb
серая
с, b
ccbb
рыжая
Р ССВВ х ссвв
серая рыжая

 Гаметы: С В
св
Гаметы: С В
св
F1 СсВв
серая
Во втором поколении при скрещивании серых гетерозиготных лошадей расщепление будет следующим:
Р СсВв х СсВв
серая серая
Гаметы: СВ, Св, сВ, св СВ, Св, сВ, св
-
♂ ♀
СВ
Св
сВ
св
СВ
ССВВ
серая
ССВв
серая
СсВВ
серая
СсВв
серая
Св
ССВв
серая
ССвв
серая
СсВв
серая
Ссвв
серая
сВ
СсВВ
серая
СсВв
серая
ссВВ
вороная
ссВв
вороная
св
СсВв
серая
Ссвв
серая
ссВв
вороная
ссвв
рыжая
Таким образом, при доминантном эпистазе в случае, когда признак определяется двумя парами генов, расщепление будет: 12 серых, 3 вороных и 1 рыжая.
В некоторых случаях супрессором может быть и рецессивный ген. Так., например, у мышей имеется ген А, определяющий серую окраску шерсти, и рецессивный ген а (черная окраска). Ген другой пары В способствует образованию пигмента, а его рецессивный аллель в подавляет синтез пигмента и мыши становятся альбиносами. Таким образом, гетерозиготные по обоим генам мыши (АаВв) имеют серую окраску. При скрещивании их между собой получится расщепление в соотношении 9 серых, 4 белых и 3 черных.
Комплементарное действие генов. При комплементарности признак определяется взаимодействием нескольких пар неаллельных генов. Каждый из этих генов в отдельности не способен определить развитие признака. Признак проявляется только в присутствии доминантных генов всех пар.
С явлением комплементарности столкнулся еще Мендель. При скрещивании белоцветковых сортов фасоли он обнаружил в потомстве окрашенные формы. Появление их невозможно было объяснить исходя из открытых им законов. И только позже было высказано предположение, что признак окраски цветков сложный. Для появления окраски нужно по крайней мере присутствие двух веществ – предшественника пигмента и фермента. В результате биохимических реакций под действием фермента неактивная форма предшественника пигмента превращается в активную форму, что и определяет окраску.
Синтез предшественника пигмента контролируется доминантным геном А, а синтез фермента определяет доминантный ген В. Рецессивные аллели этих генов блокируют синтез обоих веществ. При скрещивании белоцветковых форм растений, у которых заблокирован синтез предшественника пигмента (ааВВ) или фермента (ААвв), у гибридов первого поколения появляется окраска. Наследование этого признака в первом и втором поколении показана на схеме.
-
Взаимодействующие
гены
Генотипы
Признак
А,В
ааBB,ААВв, АаBВ, АаBв
красные
А,в
ААвв, Аавв
белые
а, B
ааBB, ааBв
белые
а, в
аавв
белые
Р ААвв х ааВВ
белые белые
F1 АаВв
красные
Р АаВв х АаВв
красные красные
Гаметы: АВ, Ав, аВ, ав
F2 9 А-В-, 3 А-вв, 3 ааВ-, 1 аавв
красные белые
Типичное расщепление во втором поколении при комплементарности, когда признак определяется взаимодействием двух пар генов 9 : 7.
Примером комплементарного взаимодействия генов может служить также наследование формы гребня у кур. Известно, что ген розовидного гребня R доминирует над геном простого гребня г. Ген другой пары Р определяет гороховидный гребень, а его рецессивная аллель р – простой. Присутствие в генотипе птицы доминантных генов R и Р приводит к поялению нового признаках - ореховидной формы гребня.
При скрещивании гомозиготных кур, имеющих розовидный гребень, с петухами с гороховидным гребнем у всех потомков первого поколения будет ореховидный гребень (RrPp). Во втором поколении произойдет расщепление
P RrPp x RrPp
ореховидный ореховидный
F2 9 R-P- 3 R-pp 3rrP- 1 rrpp
Фенотип: ореховидный розовидный гороховидный простой
Полимерия. При полимерии, или полимерном наследовании, на один и тот же признак влияет несколько разных генов. Каждый из этих генов усиливает развитие признака. Такие однозначно действующие гены называются аддитивными. Впервые такой тип взаимодействия генов был установлен Нильсоном-Эле при изучении наследования окраски чешуи овса и зерен пшеницы.
Рассмотрим пример наследования окраски прилистников злаков при взаимодействии двух пар полимерных генов. Полимерные гены обозначают одной и той же буквой. Для их отличия используют индексы. В зависимости от количества доминантных генов и проявляется развитие признака.
-
Взаимодействующие гены
Признак
А1А1 А2 А2
А1 А1 А2 а2
А1 А1 а2 а2
А1 а1 а2 а2
а1 а1 а2 а2
темно-зеленая
зеленая
светло-зеленая
желтая
белая
При скрещивании гетерозиготных форм во втором поколении происходит расщепление по фенотипу – 1 : 4 : 6 : 4 : 1.
Полимерный тип взаимодействия генов имеет большое значение при наследовании количественных признаков у сельскохозяйственных животных. К количественным признакам относятся признаки, характеризующие продуктивность животных: величина удоя, живая масса, настриг шерсти у овец, яйценоскость кур и др. Все эти признаки формируются под влиянием многих генов, каждый из которых усиливает развитие признака. Так, например, настриг шерсти у овец зависит от генов, контролирующих длину и тонину шести, густоту шерсти и складчатость кожи и других.
Гены - модификаторы. Гены, не проявляющие собственного действия, но усиливающие или ослабляющие эффект действия других генов, называются генами-модификаторами. Изучение окраски у домашних животных показало, что наряду с крайними формами, обладающими полным развитием пигмента или его отсутствием, наблюдается целый ряд генотипически обусловленных переходных форм. В организме имеется не менее трех пар генов—модификаторов, влияющих на количество красного пигмента в волосе крупного рогатого скота. В результате у гомозиготных по рецессивному гену красной масти животных интенсивность окраски колеблется от вишневой, как у скота красной горбатовской породы, до почти 6елой с желтоватым оттенком у коров ситмментальской породы.
У кроликов породы бабочка имеется ген пятнистой окраски. В зависимости от действия генов-модификаторов пятнистость сильно варьирует. Есть животные почти черной окраски с белыми пятнами на лапках и кончике носа и животные с белой окраской, у которых отмечаются черные пятна только на лапках и вокруг глаз. Кроме этого, существует целый ряд переходных форм с расположением черных пятен по всему телу.
Гены-модификаторы имеют большое селекционное значение. В результате накопления желательных небольших изменений признака, вызванных генами-модификаторами, можно усилить степень его развития, подавить развитие нежелательных признаков и даже изменить степень доминирования того или иного признака.
Лекция 4