

Функции эритроцитов. Механизм транспорта кислорода эритроцитами, аллостерическая регуляция сродства гемоглобина к кислороду. Роль эритроцитов в транспорте углекислого газа.
Функции эритроцитов:
Дыхательная - транспорт кислорода и участие в транспорте углекислого газа.
Адсорбция и транспорт питательных веществ.
Адсорбция и транспорт токсинов.
Регуляция ионного состава плазмы крови.
Формирует реологические характеристики крови/вязкость и т.д./
Гемоглобин присоединяет кислород, с образованием оксигемоглобина Эта реакция обратима:
Нв+О2=НвО2
Направленность реакции зависит от содержания кислорода: если количество кислорода в крови увеличивается, то реакция идет в сторону образования оксигемоглобина, если уменьшается - то в противоположную сторону.
С понижением рН освобождение О2 гемоглобином облегчается. То же происходит при повышении концентрации СО2. Это чрезвычайно важно с физиологической точки зрения, так как в тканях с быстро протекающим обменом веществ, таких, как мышечная ткань, образуется много СО2 и кислот. Высокие уровни СО2 и Н+ стимулируют освобождение О2 из гемоглобина, и таким образом удовлетворяется потребность в большом количестве кислорода в метаболически активных тканях. После освобождения кислорода дезоксигемоглобин присоединяет Н+ и СО3. Для альвеолярных капилляров легких характерна более высокая концентрация кислорода, и там по мере связывания дезо-ксигемоглобином кислорода происходит освобождение Н+ и СО2.
Транспорт СО2 эритроцитами.С02 диффундирует из тканей в плазму крови, а из нее — в эритроциты по градиенту парциального давления С02. В эритроцитах С02 образует с водой угольную кислоту, которая диссоциирует на Н+ и HCO3. (С02 + Н20 = Н2СО3 = Н+ + HCO3). Эта реакция протекает быстро, поскольку С02 + Н20 = Н2СОэ катализируется ферментом карбоангидразой мембраны эритроцитов, которая содержится в них в высокой концентрации.
Н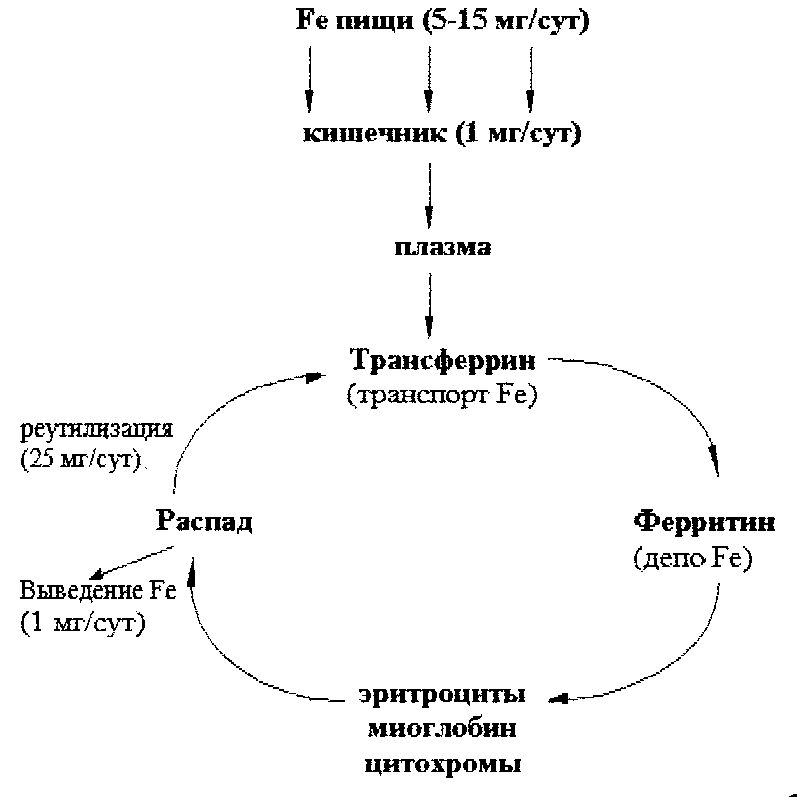 ебольшое
количество углекислого газа (10%)
транспортируется в виде карбогемоглобина
– соединения CO2 c
гемоглобином.
ебольшое
количество углекислого газа (10%)
транспортируется в виде карбогемоглобина
– соединения CO2 c
гемоглобином.
Обмен железа. Лабораторные показатели дефицита железа в организме. Понятие о физиологической желтухе новорожденных.
В организме взрослого человека содержится 3-4 г железа, из этого количества около 3,5 г находится в плазме крови. Гемоглобин эритроцитов содержит примерно 68 % всего железа организма, ферритин – 27 % (резервное железо печени, селезенки, костного мозга), миоглобин (в мышцах) – 4 %, трансферрин (в плазме крови) – 0,1. На долю всех содержащих железо ферментов приходится примерно 1 % железа, имеющегося в организме.
В обмене железа принимает участие ряд белков.
Апоферритин. Белок связывает железо в эритроцитах и превращается в ферритин, который остается в энтероцитах. Таким способом регулируется поступление железа в капилляры крови из клеток кишечника. Когда потребность организма в железе невелика, скорость синтеза апоферритина повышается. При недостатке железа в организме апоферритин в энтероцитах почти не синтезируется.
Трансферрин. Это транспортный белок, относится к гликопротеинам, синтезируется в печени. Он имеет два центра связывания железа. Трансферрин транспортирует железо с током крови к местам депонирования и использования. В норме трансферрин насыщен железом приблизительно на 33 %.
Ферритин. Олигомерный белок с молекулярной массой 450 к Да. Он состоит из 24 идентичных протомеров, образующих полую сферу. Железо депонируется в ферритине в виде гидроксифосфата. Содержание железа в молекуле ферритина непостоянно. Функция ферритина – депонирование железа. Ферритин содержится почти во всех тканях, но в наибольшем количестве в печени, селезенке, костном мозге.
Дефицит железа. В общем анализе крови при железодефицитной анемии будут регистрироваться снижение уровня гемоглобина и эритроцитов. Умеренная эритроцитопения может проявляться при Нb <98 г/л, однако снижение эритроцитов <2 х 1012/л для ЖДА не характерно. При ЖДА будут регистрироваться изменения морфологических характеристик эритроцитов и эритроцитарных индексов, отражающих количественно морфологические характеристики эритроцитов.При развитии ЖДА в биохимическом анализе крови будут регистрироваться: • уменьшение концентрации сывороточного ферритина; • уменьшение концентрации сывороточного железа; • повышение ОЖСС (общая железосвязывающая способность сыворотки ); • уменьшение насыщения трансферрина железом.
Физиологическая желтуха новорожденных — физиологическое преходящее состояние, которое отмечается у большинства здоровых новорожденных и является следствием временного повышения уровня билирубина в крови ребенка. У новорожденных после рождения отмечается повышенный распад красных кровяных телец (эритроцитов), что приводит к увеличению содержания в крови продукта их распада — желтушного пигмента (билирубина). Вследствие возрастной незрелости печени билирубин выводится из организма несколько медленно, благодаря чему у некоторых детей кожные покровы и склеры глаз временно могут приобретать желтый оттенок. Симптомы физиологической желтухи новорожденных появляются в большинстве случаев на вторые или третьи сутки после рождения ребенка, а затем медленно исчезает к двум-трем неделям.
Схема распада гемоглобина. Механизм токсического действия билирубина. Продукты распада гемоглобина в крови, моче, кале в норме.
Начальным этапом распада гемоглобина является разрыв одного метинового мостика с образованием вердоглобина. В дальнейшем от молекулывердоглобина отщепляются атомжелеза и белокглобин. В результате образуется биливердин. Затем биливердин, восстанавливаясь,превращается в билирубин – пигмент, выделяемый с желчью и поэтому называемыйжелчным пигментом. Образовавшийся билирубин называется непрямым(неконъю-гированным) билирубином,поскольку вследствие плохой растворимости в воде он легко адсорбируется на белках плазмы крови (альбуминах).
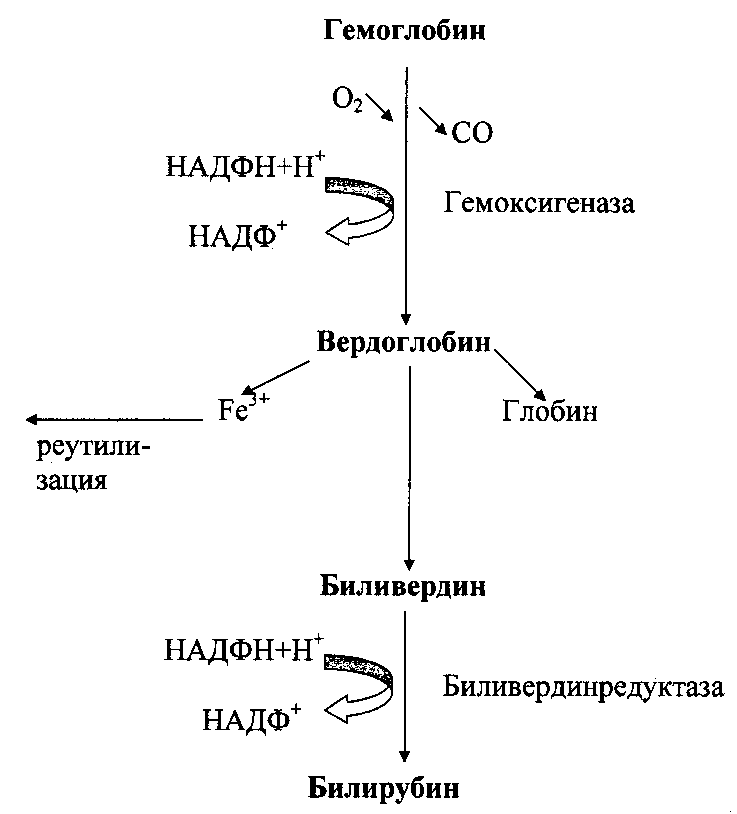
Комплекс альбумин-билирубин с током крови попадает в печень, где происходит его превращение в прямой билирубин путем коньюгации с глюкуроновой кислотой. Реакцию катализирует фермент УДФ-глюкуронилтрансфераза. Образующийся билирубиндиглюкуронид получил название прямого (коньюгированного) билирубина или связанного.Прямой билирубин – это нормальный компонент желчи, попадающий в кровь в незначительном количестве.
Механизм токсического действия билирубина:
- снижение синтеза АТФ
- снижении скорости реакций, идущих с затратой АТФ
- разобщение процессов тканевого дыхания и окислительного фосфорилирования
Распределение желчных пигментов в норме: кровь – общий билирубин – 8,5 – 20,5 мкмоль/л; непрямой билирубин – 1,7 – 17,1 мкмоль/л; прямой билирубин – 2,2 – 5,1 мкмоль/л; моча – стеркобилиноген – 4 мг/сутки; кал – стеркобилиноген.