

3.2. Липидные компоненты клеточных мембран
До 50% липидов клеточных мембран составляют производные фосфатидных кислот, которые представляют собой продукт этерификации диацилглицеринов фосфорной кислотой по первичной спиртовой группе, например:
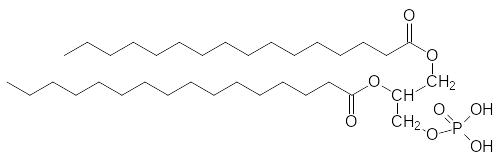
В свою очередь фосфатидные кислоты можно рассматривать как про-дукты диацилирования L-глицерин-3-фосфата или, что то же самое, D-гли-церин-1-фосфата, но стерические эффекты у жиров и производных фосфа-тидных кислот особой роли не играют. На основе фосфатидных кислот в организме образуется несколько важных амфифильных соединений, пред-ставляющих собой продукты этерификации одной из кислотных групп фосфорной кислоты гидроксильными группами холина, этаноламина или серина:

Эта формула соответствует фосфатидилхолину (лецитин, эфир фосфа-тидной кислоты и холина HOCH2CH2N+(CH3)3), другими соединениями этой группы являются фосфатидилэтаноламин (кефалин, эфир фосфатид-ной кислоты и этаноламина HOCH2CH2NH2) и фосфатидилсерин (эфир фосфатидной кислоты и аминокислоты серина НОСН2СН(NH2)СООН).
Амфифильные свойства могут быть также обусловлены неионогеннми гидрофильными остатками. При этерификации одной из гидроксильных групп фосфатного фрагмента остатком инозита – гексагидроксицикло-гексана – образуется фосфатидилинозит, а его гидроксильные группы могут быть дополнительно фосфорилированы. Определённую роль в построении мембран играют и простые эфиры глицерина – вещества, по-строенные по типу фосфатидных производных, у которых один из остат-ков жирной кислоты (у первичной гидроксильной группы) заменен на енольный остаток или остаток жирного спирта (плазмалогены), например:
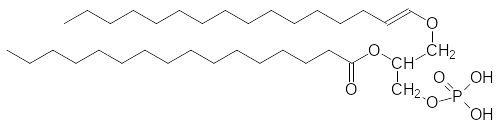
В этих веществах фрагмент фосфорной кислоты также образует эфир-ные связи с различными гидрофильными остатками.
Близки фосфатидилхолину по биологическим функциям сфингомие-лины, построенные на основе ненасыщенного аминодиола – сфингозина

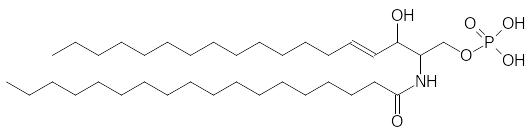
Ацилированный по аминогруппе остатком жирной кислоты сфингозин – это церамид, образующий аналогичный фосфатидной кислоте эфир с фосфорной кислотой (приведена формула фосфата церамида):
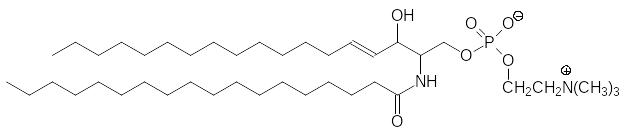
Эфир фосфата церамида и холина – это сфингомиелин:
В церамидных производных гидрофильный фрагмент может быть представлен остатками полиоксисоединений, в частности сахаров. Цер-амид, соединённый гликозидной связью с глюкозой – это глюкоцереб-розид:

В результате присоединения к первичной гидроксильной группе цер-амидов олигосахаридных остатков образуются ганглиозиды, которые фор-мируют антигенные и рецепторные участки на клеточных мембранах.
В построении клеточных мембран участвуют и липиды терпеноидной природы, построенные из фрагментов изопрена. В их число входят полине-насыщенные линейные и алициклические соединения:

Сквален
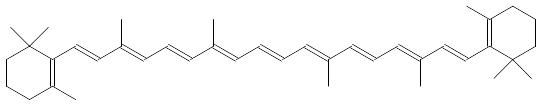
β-Каротин
В состав мембран входят также стероиды, например, холестерин и эргостерин:
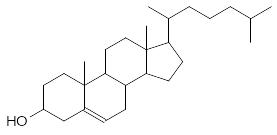

Холестерин Эргостерин
Из дегидрохолестерина (эргостерина) на свету образуется витамин D, регулирующий, в частности, кальциевый обмен. В норме в организм чело-века он должен поступать (или образовываться) в количестве около 10 мкг в сутки, а доза в 1,5 мг этого витамина – уже яд. Скорость образования витамина D в значительной мере зависит от светового потока, поэтому лю-ди южных широт имеют тёмную кожу (светлокожие загорают), а северяне – блондины. Правда, есть ещё смуглые и черноволосые эскимосы, но они получают много витамина D с рыбой и мясом морских животных.
Вещества стероидной природы очень важны в составе клеточных мем-бран всего живого. Они определяют их лабильность, проницаемость для многих необходимых клетке веществ, для которых нет специальных транс-портных систем. Если мембраны клеток животных включают преимущест-венно холестерин, то мембраны растительных клеток содержат большое число (в зависимости от растения) стероидов иного строения. Первичным продуктом стероидной природы, образующимся из сквалена (тритерпен линейного строения), становится чаще всего ланостерин:
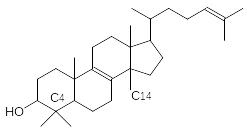
Ланостерин трижды подвергается деметилированию (две метильные группы у С4 и метильная группа С14) и трансформации по боковой цепи. Окисленные стероиды с карбоксильной группой в укороченной боковой цепи – это желчные кислоты. Они играют роль поверхностно-активных веществ в пищеварительной системе. Без них скорость переваривания жиров резко снижается. Растительные клетки, наоборот, усложняют эту боковую цепь. Паразитирующие на растениях грибы чаще всего исполь-зуют в качестве исходного продукта для своего мембранного стероида – эргостерина – ланостерин. Нарушение деметилирования ланостерина по С14 лежит в основе действия современных противогрибковых препаратов – фунгицидов. Стероидную природу имеют многие гормоны и токсины.