

7.5. Катаболические превращения аминокислот
Даже при хорошо сбалансированном питании от 15 до 10 % энергии организм человека получает из аминокислот. Это связано в основном с тем, что поступающие пищевые белки не соответствуют по аминокислот-ному составу синтезируемым в организме в разное время белкам, поэтому «лишние» аминокислоты включаются в катаболические превращения. Кро-ме того, в организме постоянно идёт процесс обновления собственных бел-ков. Так, например, многие ферменты имеют время полупревращения, рав-ное нескольким часам, хотя, конечно, структурные белки более долговеч-ны. Процесс сборки-разборки белков (а в него включается до нескольких сот граммов белков в день) не идеален, и часть образовавшихся при этом аминокислот также включается в катаболические превращения.
На начальной стадии катаболического превращения аминокислот при катализе трансаминазами (пиридоксальфосфат-зависимые ферменты) обра-зуются -кетокислоты. В общем виде эта реакция представляется как обмен аминогруппами и кетогруппами между аминокислотами и кетоглу-таровой кислотой:

т.е. аминокислота превращается в -кетокислоту, а -кетоглутаровая кислота превращается в глютаминовую кислоту.
Химизм этого процесса представляется следующим образом: сначала пиридоксальфосфат, связанный с белковой частью фермента (Е) за счёт взаимодействия его альдегидной группы с аминогруппой лизинового фраг-мента белка, образует такую же связь с аминогруппой дезаминируемой аминокислоты. При этом ковалентная связь кофермента с белком разры-вается и образуется комплекс белка с альдимином, образованным пирид-оксальфосфатом и аминокислотой:

После этого происходит миграция двойной связи с образованием уже кетимина из пиридоксаминфосфата и -кетокислоты:
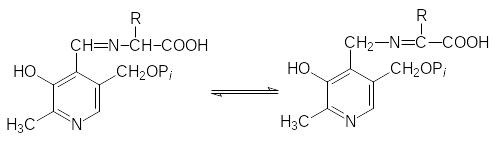
На следующей стадии превращения протекает гидролиз кетимина с образованием -кетокислоты и пиридоксаминфосфата:
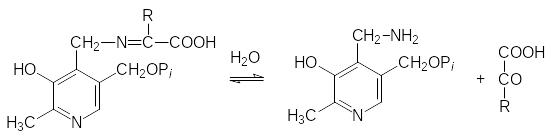
Для дезаминирования другой молекулы аминокислоты пиридоксамин-фосфат должен снова превратиться в пиридоксальфосфат. Этот процесс сопровождается переносом аминогруппы с пиридоксаминфосфата на дру-гую кетокислоту (чаще всего при этом ‑кетоглутаровая кислота превра-щается в глютаминовую кислоту):
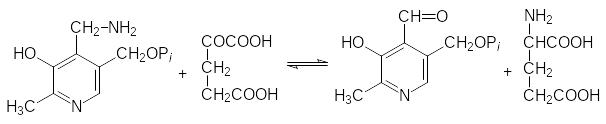
Образовавшая в этой реакции глютаминовая кислота снова превраща-ется в -кетоглутаровую по механизму дезаминирования, который будет рассмотрен ниже (с. 153) при изучении биосинтеза мочевины.
Полученные в результате дезаминирования аминокислот кетокислоты разными путями превращаются в ацетилкофермент А или в молекулы глюкозы. Так, аланин превращается в пируват, глютаминовая кислота и глютамин превращаются в кетоглутарат, аспарагиновая кислота и аспара-гин – в оксалоацетат (эти кетокислоты участвуют в цикле Кребса) и т.д.
При катаболическом превращения фенилаланина и тирозина дезами-нирование должно быть дополнено превращениями ароматического остат-ка с образованием молекул, включающихся в цикл Кребса. Эта схема уни-фицирована для двух аминокислот и поэтому «лишний» фенилаланин сна-чала превращается в тирозин в результате окисления кислородом в при-сутствии фенилаланинмонооксигеназы:

Затем тирозин по представленной выше схеме (с. 148-149) дезамини-руется соответствующей трансаминазой и превращается в гидроксифенил-пировиноградную кислоту:

которая действием кислорода в присутствии соответствующей диоксиге-назы превращается в гомогентизиновую кислоту:
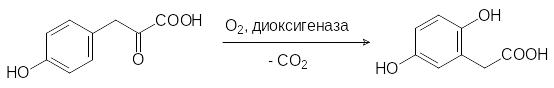
В результате этой реакции происходит окисление (гидроксилирова-ние) ароматического цикла, окислительное декарбоксилирование боковой цепи и её миграция в соседнее положение ароматического цикла.
На следующем этапе гомогентизат-1,2-диоксигеназа окисляет гомо-гентизиновую кислоту с раскрытием ароматического цикла (показана связь, раскрываемая молекулой кислорода):

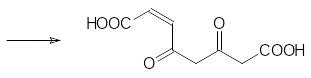
Образовавшаяся малеилацетоуксусная кислота ацилирует кофермент А с участием АТФ и при катализе тиолазой последовательно взаимо-действует с коферментом А с разрывом С-С-связей по -дикарбонильным участкам, расщепляясь на малеилкофермент А и две молекулы ацетил-кофермента А, включающиеся в цикл Кребса (малеилкофермент А для этого гидролизуется и изомеризуется в фумаровую кислоту).
Этот нормальный метаболизм фенилаланина может быть нарушен из-за дефектов генов, кодирующих участвующие в превращении ферменты. При пониженной активности фенилаланинмонооксигеназы избыток по-ступающего с пищей фенилаланина (а это одна из незаменимых амино-кислот) превращается в фенилпировиноградную кислоту. Она выводится из организма через почки (фенилкетонурия), но часть её реагирует с неко-торыми важными биомолекулами нервной ткани (такие реакции, протека-ющие без участия ферментов, называют иногда параметаболическими). Эти превращения приводят к повреждению клеток центральной нервной системы.
Наследственная болезнь фенилкетонурия, приводящая в детском воз-расте к слабоумию, должна быть диагностирована на самом раннем этапе развития новорожденного. Среди белокожего населения США её частота около одного случая на двадцать тысяч (среди афроамериканцев болезнь встречается реже). Это означает, например, что в США рождается в год около 3000 детей с фенилкетонурией. Если с первых дней жизни ребёнка давать ему специальную диету с пониженным содержанием фенилаланина, то он развивается нормально. Поэтому всем новорожденным надо сразу после рождения класть в пелёнки индикаторную бумагу, реагирующую на бензилкетоны. В возрасте 12-14 лет можно уже переходить на обычную диету, но если женщина с фенилкетонурией собирается стать матерью, то ей снова надо перейти на специальную диету ещё до зачатия ребёнка.
При пониженной активности гомогентизиндиоксигеназы непревра-щенная гомогентизиновая кислота выводится через почки и, окисляясь на воздухе, окрашивает мочу в черный цвет, однако, особых последствий для организма при этом не наблюдается. Гомогентизиновая кислота и в норме является исходным продуктом для образования меланина – вещества, окрашивающего кожу при загаре и волосы.
При рассмотрении механизма дезаминирования аминокислот пирид-оксальфосфатзависимыми трансаминазами говорилось о том, что для реге-нерации пиридоксальфосфата из пиридоксаминфосфата живая природа использует кетоглутаровую кислоту, один из метаболитов цикла Кребса. Понятно, что образующаяся при этом глютаминовая кислота должна быть снова превращена в кетоглутарат. Для этого предназначена дегидрогеназа, осуществляющая реакцию:
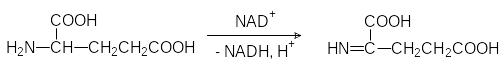
L-Глютаматдегидрогеназа – это регуляторный фермент с молеку-лярной массой около 300 кД, состоящий из шести идентичных субъединиц. Его активность возрастает в присутствии аденозиндифосфата, а отрица-тельным эффектором является ГТФ, образующийся в цикле Кребса. Иминопроизводное кетоглутаровой кислоты гидролизуется водой с образованием кетоглутаровой кислоты и аммиака:

Обратный путь превращения кетоглутарата в глютамат также сущест-вует, но на стадии восстановления иминопроизводного участвует не L‑глютаматдегидрогеназа, а другой NADP+-зависящий фермент.
Аммиак образует в водной среде сильное основание, и поэтому он токсичен для всего живого. Буферные системы жидких сред организма справляются с небольшими концентрациями аммиака, но, тем не менее, он как можно скорее должен быть переведён в нетоксичное производное. Один из путей детоксикации аммиака представлен превращением пирувата в аланин по схеме, аналогичной обратному процессу превращения глют-аминовой кислоты в кетоглутаровую. По другому пути аммиак ацили-руется глютаминовой кислотой с образованием глютамина:
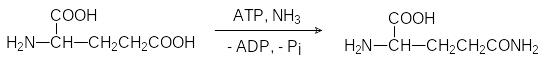
Этот путь играет важную роль в детоксикации аммиака в центральной нервной системе, где глютаминовая кислота содержится в большом коли-честве, участвуя в передаче нервных импульсов. Аланин снова превра-щается в пируват с участием трансаминаз, а глютамин гидролизуется и снова превращается в глютаминовую кислоту и аммиак. В организме теп-локровных этот процесс протекает в клетках печени, но другие виды живого используют иные пути. Проще всего проблему вывода аммиака решают водные животные: они просто выделяют его в воду. В жабрах рыб, личинок амфибий находится фермент глютаминаза, и образовавшийся в результате гидролиза глютамина аммиак просто диффундирует из крови в омывающую жабры воду (рыба тухнет с головы).
Для животных такой путь выведения аммиака не годится. Свободный аммиак не может поступать в мочу, так как для поддержания допустимой для биологических сред концентрации аммиака требуется слишком много воды. Эта проблема не может быть решена и за счёт образования нейтраль-ных солей аммония, так как органические кислоты на это тратить слишком расточительно, а потеря неорганических анионов нарушит водно-солевой баланс организма. Существует также возможность образования бикарбо-ната аммония, и не исключено, что на ранних этапах эволюции этот вари-ант использовался живой природой, но всё же бикарбонат аммония слиш-ком нестоек и его растворы имеют сильно щелочную реакцию. Поэтому в процессе эволюции был выработан механизм выведения аммиака в виде нейтрального амида угольной кислоты – мочевины. Правда, и для выве-дения мочевины надо много воды, поэтому, например, птицы (для них важен вес) и рептилии, живущие часто в зонах с недостатком влаги, выра-ботали иной механизм. Они выделяют избыточный аммиак в виде нерас-творимой в воде мочевой кислоты:
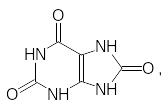
механизм биосинтеза которой здесь не рассматривается.
Оказалось, что мочевина образуется в циклическом процессе, в рас-крытии механизма которого активное участие также принимал Кребс. В ходе исследования биохимических процессов, протекающих в тканях печени, было установлено, что в присутствии трёх аминокислот: орнитина, цитруллина и аргинина


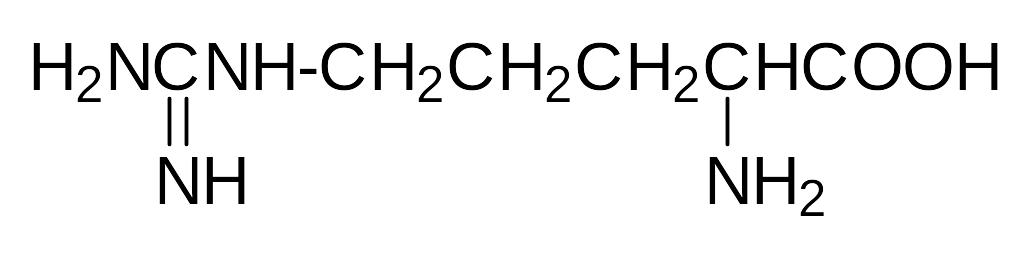
резко возрастает скорость образования мочевины. До этого было известно, что аргинин гидролизуется водой с образованием мочевины и небелковой аминокислоты орнитина, и можно было предположить, что эти амино-кислоты участвуют в образовании мочевины, а связующим между ними служит цитруллин.
Позже было показано, что аргинин действительно образуется из орни-тина через цитруллин и может гидролизоваться далее в присутствии аргиназы по реакции:

В митохондриях аммиак и диоксид углерода (из цикла Кребса) с участием двух молекул АТФ в присутствии фермента карбамоилфосфат-синтетазы I образуют смешанный ангидрид карбаминовой и фосфорной кислот – карбамоилфосфат:
![]()
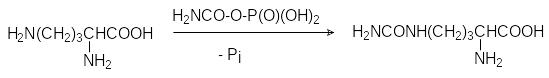
Это соединение ацилирует орнитин по концевой аминогруппе (уходя-щей группой является фосфатный остаток) с образованием цитруллина:
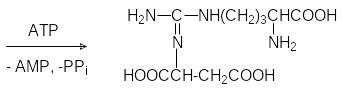
Далее цитруллин с участием АТФ реагирует с аспарагиновой кисло-той с образованием аргининосукцината. Фермент, катализирующий это превращение, называется аргининосукцинатсинтетазой; АТФ при этом распадается на АМФ и пирофосфат, гидролизующийся до фосфата:

Теперь аргининосукцинатлиаза расщепляет этот промежуточный про-дукт на аргинин и фумарат:
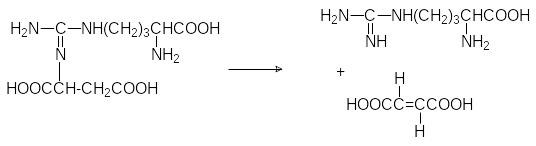
Фумарат встраивается в цикл Кребса и через малат и оксалоацетат с участием трансаминаз снова может превратиться в аспартат. Синтез арги-нина един для всего живого, но аргиназа присутствует только в клетках печени животных, выделяющих аммиак в виде мочевины (их называют уротелическими, в отличие от урикотелических, выделяющих аммиак в виде мочевой кислоты).
В результате этого циклического процесса (из аргинина образуется орнитин, с которого начиналась представленная выше цепь превращений) из организма выводятся два конечных продукта катаболических превра-щений – аммиак и бикарбонат (из СО2). Этот процесс участвует в регу-ляции рН крови, значение которого определяется содержанием бикарбо-ната. Энергетическая стоимость детоксикации аммиака достаточно высока: на один моль мочевины расходуется четыре эквивалента АТФ: два моля АТФ идут на образование карбамоилфосфата и один моль АТФ исполь-зуется для синтеза аргининосукцината, но в этом случае аденозинтрифос-фат разлается на пирофосфат и аденозинмонофосфат, для превращения которого в АТФ нужно провести двойное фосфорилирование. Поэтому аминокислоты очень невыгодны в качестве источника энергии: их катабо-лическое превращение включает образование токсичного аммиака, на связывание которого в мочевину расходуется до 15% всего получаемого из аминокислот АТФ, кроме того, повышается нагрузка на почки.
В организме жвачных животных и верблюдов образовавшаяся в ре-зультате катаболических превращений аминокислот мочевина циркули-рует в крови, и за счёт диффузии через ткани и стенки кровеносных сосу-дов она практически полностью переходит в отделы желудка, в которых живут бактерии, перерабатывающие целлюлозу и использующие мочевину в качестве источника аммиака для синтеза аминокислот.