

8. Окислительное фосфорилирование
Суммарный процесс последовательного катаболического превраще-ния одного моля глюкозы в ходе гликолиза, окислительного декарбоксили-рования пирувата и в цикле Кребса приводит к образованию 10 молей NAD(Р)H, 2 молей FADH2, 2 молей АТФ и 2 молей ГТФ. Аэробная клетка может использовать энергию, выделяющуюся при окислении восстано-вительного потенциала NAD(Р)H, для образования максимального коли-чества АТФ. В простейшем виде эта реакция записывается уравнением:
![]()
Энергетический эффект этой реакции примерно соответствует окисле-нию водорода кислородом (220 кДж/моль). Эта энергия должна оптималь-ным образом использоваться для образования АТФ, на биосинтез которого из АДФ и фосфата затрачивается 14 ккал/моль (более 60 кДж/моль). Если учесть, что в сутки организм человека потребляет 2800 ккал/моль, то это соответствует образованию примерно 200 молей или 100 кг АТФ, хотя в организме его стационарное содержание составляет около 50 г. Эти цифры показывают с какой эффективностью работает механизм воспроизводства расходуемого АТФ в зависимости от потребности в нем, которая меняется в зависимости от рода деятельности в каждый отдельный момент (актив-ная работа, отдых, сон и т.д.).
Ещё на ранней стадии биохимических исследований было установ-лено, что энергетическими «фабриками» аэробных эукариотических кле-ток являются митохондрии, а у прокариот синтез АТФ идёт в клеточных мембранах. Митохондрии представляют собой органеллы размером около 0,51,5 мкм красновато-коричневого цвета (основной цвет пигментов, входящих в состав мембран), их число в клетках животных колеблется в пределах от нескольких сотен до нескольких тысяч:
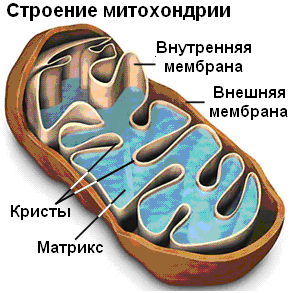
Строение митохондрий из разных организмов единообразно; они ок-ружены двойной мембраной. Внешняя гладкая мембрана содержит транс-портные белки и ферменты, отвечающие за превращения аминогрупп (моноаминоксидазы), а также ферменты, участвующие в обмене жирных кислот. Внутренняя мембрана митохондрий для увеличения её поверхнос-ти образует множество впячиваний, называемых кристами. Итак, содержи-мое митохондрии разделено на два объёма: межмембранное и внутреннее пространство (матрикс), отделённое от межмембранного пространства внутренней мембраной. Митохондрии делятся, причем они содержат свою собственную нитевидную ДНК и набор РНК, что позволяет сделать пред-положение о том, что их присутствие в клетке представляет собой нечто вроде симбиоза: они могли образоваться в результате эндоцитоза аэробных прокариот анаэробными эукариотами и сохраниться в них, получая пита-тельные вещества и снабжая приютившую их клетку аденозинтрифос-фатом. Все митохондрии в клетках организмов, размножающихся половым путем, получены от материнской клетки. Это позволяет использовать ДНК митохондрий для установления родства.
Основные катаболические процессы локализованы в матриксе мито-хондрий. Сюда соответствующими транспортными системами переносятся субстраты для цикла Кребса и катаболического превращения жирных кислот (пируват, глутамат, аспартат, малат, 2‑кетоглутарат, цитрат, жир-ные кислоты), а также АДФ и фосфат, из которых образуется АТФ. Имен-но в матриксе митохондрий протекают конечные стадии катаболического превращения углеводов и жирных кислот, сопровождающиеся образова-нием восстановительного потенциала.
Что же мы знаем о механизме образования АТФ в митохондриях? Субстратный путь образования АТФ, который реализуется живой приро-дой при анаэробном катаболическом превращении глюкозы (перенос макроэргической фосфатной связи на АДФ с 1,3-дифосфоглицерата или с фосфоенолпирувата), очевидно, не может быть достаточно эффективно использован при окислении восстановительного потенциала, заложенного в NAD(P)H и в FADH2. Поэтому эволюция клеток в кислородсодержащей среде пошла по принципиально иному пути, использующему для синтеза АТФ электрохимический потенциал и градиент концентраций протонов с коэффициентом полезного действия, достигающим 60%. Этот процесс называют окислительным фосфорилированием.
Реакция окисления NAD(Р)H может быть представлена двумя полуре-акциями. Сначала NAD(Р)H отдает пару электронов по схеме:
![]()
В стандартных биохимических условиях окислительно-восстанови-тельный потенциал Е этого процесса равен -0,320 В. Затем пара электро-нов переносится на кислород:
![]()
Здесь окислительно-восстановительный потенциал Е имеет значение +0,816 В. Для суммарной реакции окисления NAD(P)H кислородом полу-чаем разность 1,136 В, что соответствует G0 около 220 кДж/моль, тогда как на образование одного моля АТФ затрачивается более 60 кДж.
Электроны, отданные восстановленными формами никотинамидаде-ниндинуклеотида (NADH) или его фосфата (NADPH) и флавопротеинов, переносятся во внутренней мембране митохондрий в результате окисли-тельно-восстановительных превращений молекул убихинона и атомов железа в железосерных белках и в гемовых структурах в составе цито-хромов (их называют обычно пигментами электронпереносящей цепи). Движение электронов по компонентам электронпереносящей цепи сопро-вождается снижением восстановительного потенциала каждого приняв-шего электрон участника процесса. Снижение восстановительного потен-циала сопровождается выделением энергии, которая может быть использо-вана для синтеза АТФ. При этом субстратный путь с образованием макро-эргических соединений из компонент электронпереносящей цепи и фосфа-тов исключается, так как при нарушении целостности внутренней мембра-ны митохондрии она продолжает окислять NAD(Р)H, но перестаёт гене-рировать АТФ. Есть даже митохондрии с пористой внутренней мембраной (митохондрии бурого жира), которые предназначены исключительно для генерирования тепла. Было установлено, что в межмембранном про-странстве устанавливается кислая среда, и есть много соединений, которые хорошо растворяются в липидах мембран как в виде аниона, так и в протонированном виде. Оказалось, что в их присутствии даже сплошная мембрана перестает синтезировать АТФ. Такие вещества называют разоб-щителями окислительного фосфорилирования. Классическим примером таких веществ является 2,4-динитрофенол. Некоторое время его даже ис-пользовали в качестве медикаментозного средства для снижения веса. Его присутствие в организме приводит к непроизводительному расходованию пищевых веществ, выражающемуся в повышении температуры тела. Анти-биотик валиномицин, имеющий структуру баранки, составленной из повторяющихся фрагментов валина, гидроксиизовалериановой кислоты и молочной кислоты, переносит через мембраны ионы калия, снижая тем самым электрохимический мембранный потенциал. Это вещество послу-жило отправной точкой для получения циклических краун-эфиров, исполь-зуемых в качестве катализаторов межфазного переноса. Эти косвенные данные говорят о том, что движущей силой в образовании АТФ является разность концентраций протонов по разные стороны внутренней мембраны митохондрий.
Митохондриальная цепь переноса электронов включает:
никотинамидадениндинуклеотидный акцептор электронов и флаво-протеин с флавинмононуклеотидом;
белки с железосерными кластерами, в которых атомы железа (два и более) связаны с сульфгидрильными группами цистеиновых фраг-ментов белка с участием неорганических сульфидных структурных элементов;
убихинон, связанный с мембраной исключительно за счёт гидро-фобного взаимодействия олигоизопреноидного остатка этого вещества с липидными компонентами мембран;
цитохромы (cyt b, cyt c1, cyt c, cyt a, cyt a3) – белки с различными гемами.
Все компоненты цепи переноса электронов, кроме цитохрома с (этот низкомолекулярный белок легко отмывается солевыми растворами), инте-грированы в мембрану. В старых учебниках по биохимии приводится такая последовательность движения электронов по электронпереносящей цепи: