

Особенности передачи возбуждения в химическом синапсе:
-
Синаптическая задержка до 0,5 мс за счет диффузии;
-
Одностороннее проведение возбуждения;
-
Постсинаптические потенциалы
-
амплитуда постсинаптического потенциала зависит от числа рецепторов постсинаптической мембраны, прореагировавших с медиатором,
-
не подчиняются закону “все или ничего”,
-
постсинаптические потенциалы способны суммироваться,
-
распространяются электротонически, с декрементом;
-
-
Синаптическое проведение утомляемо, зависит от условий среды, является мишенью для действия фармпрепаратов, токсинов.
Действие ядов и препаратов:
-
Стрихнин конкурирует с тормозными медиаторами за рецепторы постсинаптической мембраны, вызывает судороги.
-
Столбнячный токсин нарушает экзоцитоз тормозных медиаторов из терминальных бляшек, вызывает судороги.
-
Ионы магния, марганца, никеля конкурируют с ионами кальция на пресинаптической мембране, препятствуют входу кальция в терминальную бляшку и блокируют нервно-мышечную передачу.
-
Токсин Cl. botulinum блокирует экзоцитоз ацетилхолина в нервно-мышечных синапсах.
-
Кураре конкурирует с ацетилхолином за рецепторы постсинаптической мембраны нервно-мышечных синапсов. (Недеполяризующая миореллаксация)
-
Дитилин, избыток ацетилхолина (образующийся, например, вследствие ингибирования ацетлхолинэстеразы) вызывают стойкую деполяризацию постсинаптической мембраны и не дает формироваться там потенциалу действия. (Деполяризующая миорелаксация).
Проверь себя – синаптическая передача
В чем самое главное отличие постсинаптического потенциала от потенциала действия (один ответ)?
-
Более низкая амплитуда
-
Одновременное повышение натриевой и калиевой (калиевой и хлорной) проводимости
-
Отсутствие Екр
-
Постсинаптический потенциал формируется только в определенном месте (синапсе)
Электрический синапс
-
Распространены в гладкой мускулатуре, миокарде; в ЦНС немногочисленны.
-
Возбуждающие.
-
Двухстороннее проведение.
-
Формируют функциональные синцитии.
-
Механизм проведения:
-
В целом, как по волокну: электротоническое распространение потенциала, а затем – генерация ПД на постсинаптической мембране.
-
В щелевых контактах (нексусах) – коннексоны: белки, формирующие ионные каналы, расположенные друг напротив друга в пре- и постсинаптической мембране. За счет коннексонов обеспечивается проводимость щелевых контактов.
-
Коннексоны могут закрываться при понижении рН и увеличении концентрации кальция внутри клетки.
-
Физиология мышц: поперечено-полосатые (скелетные), гладкие, сердечные
Мышца как возбудимая ткань
По сравнению с нейронами:
-
Возбудимость, скорость проведения меньше
-
Длительность ПД (РП) больше
Закон “все или ничего”
-
В скелетной мускулатуре – для отдельного волокна, но не для мышцы в целом.
-
В гладкой мускулатуре и миокарде – для целой мышцы (функциональный синцитий).
Скелетная мускулатура
Функциональная организация скелетной мускулатуры - двигательные единицы (ДЕ)
-
ДЕ – один мотонейрон и группа иннервируемых им мышечных волокон.
-
Чем меньше волокон в одной ДЕ, тем более тонкое движение можно совершить.
-
По лабильности ДЕ:
-
быстрые (длительность ПД от 1 мс, высокая скорость сокращения, гидролиза и ресинтеза АТФ)
-
медленные (длительность ПД до 10-15 мс, низкая скорость сокращения, гидролиза и ресинтеза АТФ).
-
Механизм мышечного сокращения
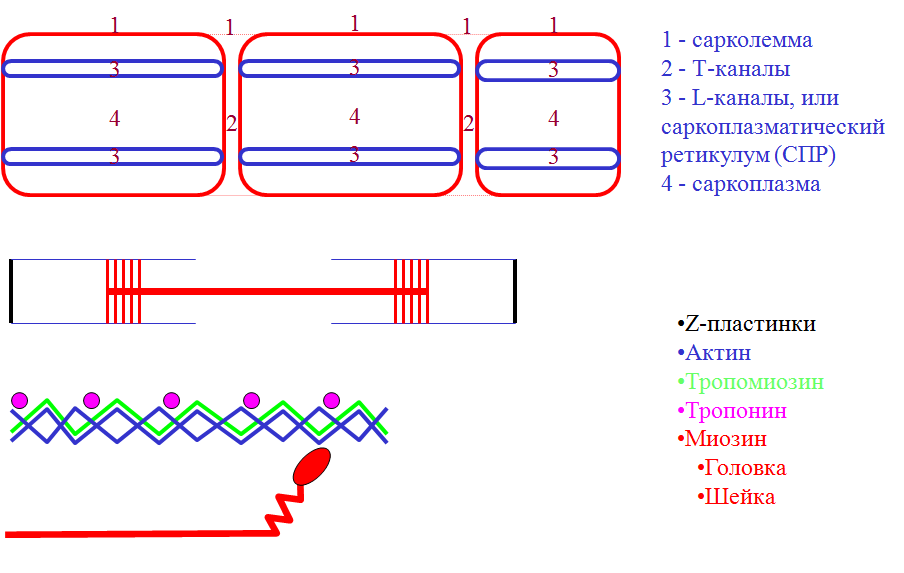
-
Распространение ПД по Т-каналам ® деполяризация СПР ® выход Са2+.
-
Са2+-тропонин ® сдвиг тропомиозина ® открытие центров связывания головки миозина на актине.
-
Соединение головки миозина с актином ® жесткость (ригидность) мышцы.
-
АТФ-азная активность головки миозина ® гидролиз АТФ ® гребковое движение (генерация усилия).
-
Присоединение новой молекулы АТФ к миозиновой головке ® разрыв поперечного мостика (исчезновение ригидности).
-
Присоединение головки миозина к другому центру актина.
-
Активация Са-АТФазы СПР фосфатом (гидролиз АТФ) ® возврат Са2+ обратно ® расслабление мышцы.
Регуляция силы и скорости сокращения
Суммация
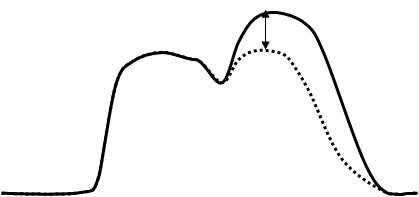
-
амплитуды сокращения – после первого сокращения часть Са2+ не успевает возвратиться в СПР (движение против градиента концентраций требует АТФ) и суммируется с вновь поступившей порцией.

-
Слитное сокращение – тетанус.
-
Гладкий – новая фаза укорочения накладывается на предыдущую.
-
Зубчатый – новая фаза укорочения накладывается на фазу удлинения.
-
-
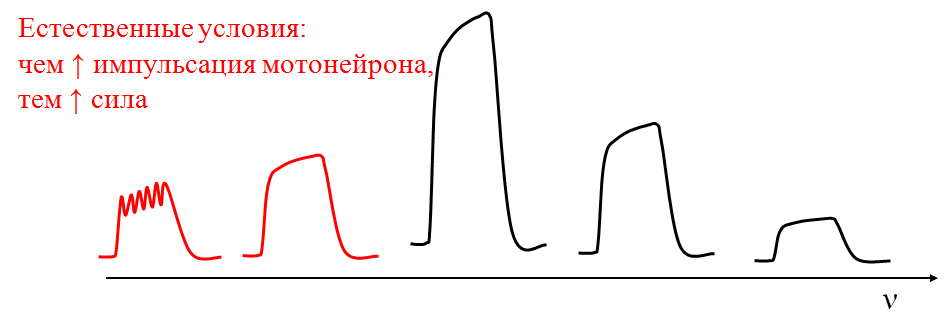
В естественных условиях гладкий тетанус целой мышцы возникает за счет несинхронности зубчатых тетанусов отдельных ДЕ.
-
Так как возврат Са2+ медленнее его выхода, чем частота возбуждения, тем амплитуда и сила сокращения;
-
Но небезгранично: имеется оптимум частоты возбуждения, после которого развивается пессимальное торможение (пессимум).
Зависимость длина-сила (зависимость Франка-Старлинга)

Зависимость длина-сила (зависимость Франка-Старлинга) - зависимость силы мышцы от степени ее растяжения
¯ силы при больших длинах:
-
¯ перекрывания нитей актина и миозина
¯ силы при малых длинах:
-
¯ электромеханического сопряжения;
-
“сморщивание” саркомера
Режимы сокращений
-
Изотонический – при постоянном напряжении
-
Изометрический – при постоянной длине (без укорочения) – максимальная сила
-
Концентрический – усилие при укорочении
-
Эксцентрический – усилие при удлинении
Зависимость нагрузка-скорость (сила-скорость)

Чем больше нагрузка, тем медленнее сокращение. Чем быстрее сокращение, тем оно слабее.
Причины:
-
Длительность актомиозинового взаимодействия ~ сила.
-
При большой скорости укорочения поперечные мостики не успевают распадаться.