

Биоэнергетика живых систем и Макроэргические соединения
Свободная энергия и биологическое окисление.
Движение и перемещение в пространстве живого микроскопического, как бактерия, или макроскопического, как спортсмен, объекта, как и внутренние, невидимые невооруженному глазу, биохимические процессы, требуют затрат энергии (как любая работа). Необходимую для совершения работы энергию все живые биологические объекты получают с пищей, извлекая часть ее из химических связей молекул (пищи, конечно). Однако на полезную (понимай – созидательную) работу может быть затрачена не вся, а только свободная энергия (G) системы (например, молекулярной) при постоянной температуре и давлении. Обладание (кроме «занятой») некоторой долей свободной энергии – неотъемлемое (да и необходимое) свойство живой материи. Физикой живые биологические объекты рассматриваются как открытые, неравновесные системы, причем далекие от термодинамического равновесия, означающего для них – «смерть». В этом смысле человеческий спорт – способ быть подальше от «смертельного равновесия».
В биохимии, условимся, под свободной энергией системы понимать способность совершать полезную работу, такую, которая может быть затрачена на созидание – образование чего-то нового, более сложного и упорядоченного (на креатив, как теперь выражаются продвинутые).
На нашей планете главный источник свободной и доступной для использования живыми организмами энергии – Солнце – ближайшая звезда. В результате протекающих на Солнце ядерных реакций (вспоминайте, про синтез атомов гелия) огромное количество энергии в виде электромагнитного излучения (γ-лучей и ультрафиолета, видимого и инфракрасного – теплового, миллиметрового и далее) и элементарных частиц ежесекундно выбрасывается в окружающее пространство (и Земле достается лишь малая толика от него). Значительную часть высокоэнергетического излучения – жесткого, способного разрывать химические связи (и таким образом «мешать жить») задерживает атмосфера Земли. Видимое (нами и многими другими живыми организмами), тепловое (невидимое, но ощущаемое органами чувств) и длинноволновое (радио-) излучение не разрушает вещество. Попадающие на поверхность Земли солнечные фотоны (они же электроны, когда «пойманы») живая природа успешно «приручила» с помощью фотосинтезирующих систем. Вообще, электромагнитные волны видимого диапазона практично используются биологическими системами (и, конкретно, Вами и сейчас в качестве носителя информации).
Фотосинтезирующие системы растений ступенчато – порциями поглощают кинетическую энергию электронов, превращая ее в потенциальную энергию химических связей органических молекул – углеводов (школьный курс: они синтезируются растениями из H2O и CO2). Углеводы, особенно, высокомолекулярные (крахмал, гликоген), «насыщенны» потенциальной энергией, локализованной на химических связях (которые электронными облаками прочно связывают атомы в молекуле – «браке по любви и расчету»). Фактически, растения накапливают и удерживают электромагнитную энергию Солнца, а вся зеленая (и не зеленая), наземная (и подземная) растительная биомасса Земли это огромный «аккумулятор», рис. 2. К примеру, картофель накапливает энергию «про запас», конечно, себе, а не нам, поэтому заполняет клубни крахмалом, а не мышечным гликогеном («спортивно» устрашающего культуриста).
 Рис.
2. Солнце – первичный источник энергии
живых организмов, в том
Рис.
2. Солнце – первичный источник энергии
живых организмов, в том
числе, спортсменов.
Окисляя («сжигая») органические вещества растений (крахмал и другие) многие живые организмы на Земле (из них и колорадские жуки, и животные, и люди) «пристрастились» использовать выделяющуюся энергию, не позволяя ей полностью (и попусту) рассеяться. (Таким образом, энергию, заработанную, между прочим, упорным трудом фотосинтезирующих систем картофельной ботвы, так называемые высшие фактически отбирают у растений, заодно приобретая массу биологического строительного вещества, да еще безвозмездно. А может и «возмездно», если вспомнить про культурное земледелие).
Окисление веществ в природе (вне живого) в ходе химических реакций (коррозия железной конструкции на воздухе, «прение» осенних листьев, горение пропан-бутановой смеси в газовой горелке или пожар, «упаси …») – это свободное окисление (с участием кислорода). Оно сопровождается выделением тепла в окружающую среду, соответственно:
- положительным изменением энтальпии +ΔH, характеризующей теплосодержание системы (становится горячо на пожаре);
- отрицательным изменением свободной энергии –ΔG, поскольку система утратила некоторую ее часть, (что сгорело, то пропало);
- положительным изменением энтропии +ΔS, указывающей, что система стала менее упорядоченной (беспорядка на пожарище больше).
При свободном окислении, подчиняющемся законам термодинамики (и химии) величина –ΔG, которая могла бы быть потрачена на совершение полезной работы, «теряется». (А вот дома, чтобы эта –ΔG не «терялась», Вы используете систему – газовую горелку, и часть выделяющегося тепла направляете другой системе – чайнику).
Процесс окисления в биологической системе – биологическое окисление, подчиняется тем же законам природы, что и свободное. Оно протекает на клеточном уровне и при участии ферментативных систем. И это окисление уже «не свободное», а «целевое» – жизнеобеспечивающее. Процесс биологического окисления в живых организмах «запрограммирован» генетически и постоянно контролируется самой биологической системой (во времени и в пространстве). По сути, он является процессом обратным фотосинтезу (поэтому пока растения растут, животные жуют).
Требования живого к биологическому окислению понятны: минимум затрат и потерь, удовлетворительный выход энергии, хороший КПД (чем больше, тем лучше), возможность контролировать процесс тепловыделения (тут «гореть» на работе ни к чему). Живым организмам энергия нужна для осуществления множества самых разнообразных процессов. На микро уровне энергообеспечение необходимо для биохимических реакций синтеза, процессов ионного переноса, создания потенциалов. На макро уровне без затрат энергии невозможны движение (вращение жгутиков, взмах крыльев или плавников, перестановка ног и бег), труд, творчество, спорт, и, в целом, разумное поведение (последнее одними только затратами энергии не определяется).
Энергетическое сопряжение как приводной механизм жизни.
Высокая степень упорядоченности живой биологической системы, и, следовательно, низкий уровень энтропии, создается самой же системой в процессе хорошо налаженного обмена веществ с окружающей средой – метаболизма (за миллионы лет эволюции живая система сама себе его и организовала способом, который термодинамика называет самоорганизацией). Метаболизм в живых организмах осуществляется за счет энергии, выделяющейся в реакциях биологического окисления. Окисляемые химические соединения (пища – углеводы и прочие), поступают в систему из внешней среды (чаще всего, по принуждению). Общий термодинамический вывод: реакции биологического окисления используются живыми организмами как средство для уменьшения своей «собственной» энтропии (внутри системы) за счет увеличения «чужой» (снаружи). Для практической реализации этого средства в процессе эволюции живыми организмами изобретен (и «генетически» запатентован) уникальный способ – энергетическое сопряжение биохимических реакций. Математическая суть способа это подключение реакций с –ΔG к реакциям с +ΔG при таком условии, чтобы l–ΔGl > l+ΔGl, именно: первая – отрицательная разность должна быть больше второй – положительной. То есть, реакции с отрицательным изменением свободной энергии должны протекать в химическом «зацеплении» с реакциями с положительным изменением свободной энергии, рис. 3.
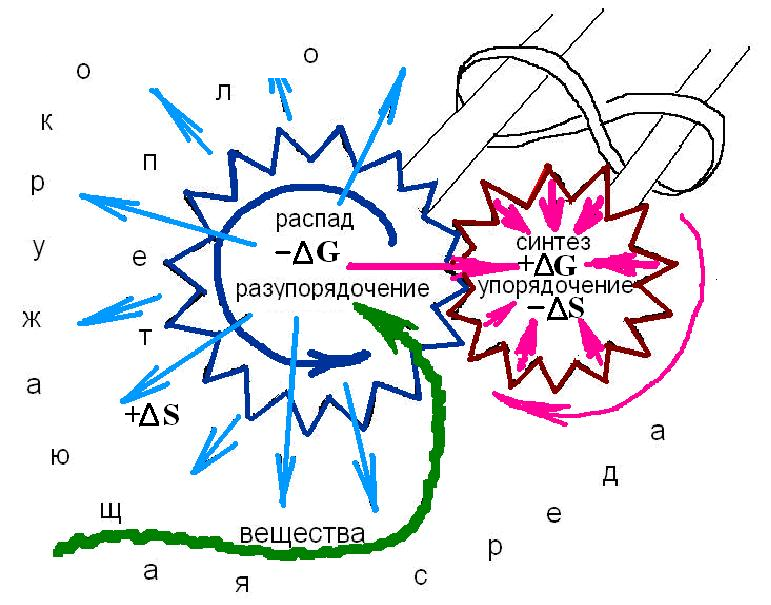
Рис. 3. Энергетическое сопряжение.
Подключая реакцию с большим отрицательным изменением свободной энергии –ΔG (реакцию окисления, например) к реакции, протекающей с положительным изменением свободной энергии +ΔG (к реакции синтеза) биологическая система совершает химическую работу и синтезирует сначала промежуточное (мало подвижное и «короткоживущее») соединение (делает «зацепление»). Химические связи этого соединения задерживают на некоторое время часть свободной энергии, выделяющейся в ходе реакции, препятствуя ее рассеиванию – превращению в тепло. И уже из промежуточного соединения синтезируется лабильный (то есть подвижный) и достаточно устойчивый (а значит и «долгоживущий»), хорошо управляемый (ферментами) «посредник» – АТФ – аденозинтрифосфорная кислота. Она аккумулирует свободную энергию «порциями» в химических связях фосфатных остатков кислоты H3PO4, их называют макроэргическими. Синтез АТФ как «посредника» оправдан: в живой биологической системе невозможно прямое, без преобразований, использование энергии химических соединений на совершение полезной работы.
Первые «патенты» на «макроэрги» (макроэргические соединения), наверно, следовало бы выдать микробам. АТФ – одно из главных изобретений химической эволюции. Это не высокомолекулярное соединение уникально и обладает рядом ценных качеств. За миллиарды лет эволюция отобрала ряд соединений – «помощников», но не нашла равнозначной замены АТФ (и уже не ищет – зачем зря время тратить? Не исключено, что «нашу» АТФ используют и в других живых мирах).
«АТФ – универсальный аккумулятор и переносчик энергии в живом организме».
Такое краткое и точное определение приводят учебники биохимии. Специфические энергетические и уникальные универсальные функции в живых организмах АТФ выполняет благодаря своим «потребительским качествам» – химическим свойствам. Она хорошо растворима и легко подвергается гидролизу (с помощью ферментов в нужное время и в нужном месте), может накапливаться (в относительно большой концентрации) в клетках, удерживая при этом свободную энергию.
Специфические энергетические функции АТФ, которые она выполняет в любом живом организме на Земле, – «посредничество» в энергетических операциях в клетке. С одной стороны процесса обмена веществ (это на стороне анаболизма) при своем образовании (синтезе из АМФ и H2PO4- или АДФ и HPO4-2) АТФ в макроэргических фосфатных связях аккумулирует свободную энергию, выделяющуюся в ходе реакций биологического окисления. С другой стороны, того же обмена веществ (на стороне катаболизма), АТФ при своем распаде высвобождает исходную АДФ (или АМФ) и передает фосфатный остаток HPO4-2 (или H2PO4-) вместе с «локально» аккумулированной на нем «порцией» свободной энергии в энергозатратную реакцию биологического синтеза.
АТФ + Н2О ↔ АДФ + Фнеорг +30 кДж/моль
АДФ + Н2О ↔ АМФ + Фнеорг +30 кДж/моль
Подвижность и возможность превращения свободной энергии химических связей АТФ в электрический потенциал ΔμH+ (на мембранах митохондрий) позволяет этому соединению выполнять уникальную функцию – «конвертирование» – превращение одной формы энергии в другую, причем эквивалентное. АТФ – уникальный, подвижный клеточный «обменник» свободной энергии. Именно это свойство позволило эволюции изобрести «механику» живых систем, в основе которой – мышечное сокращение (перемещение на молекулярном – микро- и передвижение на макро уровне).
Использовать разность потенциалов ΔμH+ для практических нужд научились еще бактерии: они перемещаются, вращая жгутиками (или фибриллами как бледные спирохеты – возбудители сифилиса). Но только изобретение эволюцией (и соответствующее «генетическое патентование») мышечного волокна, в котором осуществляется преобразование энергии химических связей АТФ в разность потенциалов ΔμH+ и далее в механическую работу обеспечило живым организмам – макросистемам огромные возможности скоростного перемещения в пространстве: полет, плавание, прыжки, бег и все возможные виды спорта.
«Циклическая жизнь» АТФ или пролог темы 3.
В живых биологических системах АТФ «работает» на микро уровне, в нужную область макро системы она легко переносится, например, потоком крови. Полезную созидательную работу, энергетически обеспечивая синтез новых соединений, АТФ выполняет за счет своего распада. Она отдает на необходимую биохимическую реакцию часть своей свободной энергии вместе с остатком фосфорной кислоты, и превращается в АДФ, соединение энергетически более бедное (чем не пожертвуешь ради жизни!). АДФ тоже может отдать остаток фосфорной кислоты, но уже «менее охотно». Уменьшение концентрации АТФ в клетке «автоматически» тормозит протекание энергетически затратных процессов: синтез веществ и их транспорт, перенос ионов и т. д. Клетка поддерживает необходимый ей гомеостаз, регулируя биохимические процессы и состав своей внутренней среды. «Рабочая» концентрация АТФ в клетке поддерживается за счет ее восстановления – повторного синтеза из продуктов распада (АДФ, АМФ и неорганического фосфата) или ресинтеза. Таким образом, в цитоплазме клетки молекулы АТФ постоянно «рождаются», «живут», «жертвуют» собой и снова «воскресают». Процесс постоянного восстановления АТФ из продуктов ее распада называется ресинтезом АТФ.
Для обеспечения потребностей единичной неподвижной клетки такого простого восстановления, достаточно, но самостоятельное перемещение в пространстве макрообъекта требует больших (макро) энергетических затрат. Работа мышц нуждается в постоянной «подпитке» энергией АТФ, а ее запас в клетке ограничен (клетка «не резиновая»). С целью увеличения числа сокращений и, соответственно, продолжительности непрерывной работы мышцы эволюция живого «изобрела» дополнительный накопитель энергии – креатинфосфат. Это макроэргическое соединение образуется из креатина путем фосфорилирования, то есть присоединения к нему фосфатного остатка, отщепляемого от АТФ. Креатинфосфат «умеет» (с помощью креатинфосфокиназы – фермента) передавать свою фосфатную группу на АДФ, которая превращается – ресинтезируется в АТФ. Накопив запас креатинфосфата в симпластах можно обеспечить ресинтез АТФ и, таким образом продлить непрерывную работу мышцы на некоторое время (на которое позволит запас).
КФкиназа
АДФ + креатинфосфат →→→ АТФ + креатин
Запас креатинфосфата (а не съеденного креатина моногидрата!) это энергетический резерв мышцы – ее энергетическая емкость (или показатель «спортивной» мощности).
Механохимическое преобразование энергии в мышце.
Выполнение непосредственной механической работы в мышце возможно только при посредничестве белков – структурной основы организма. Молекула АТФ связывается в особый комплекс с миозином – мышечным структурным белком, который одновременно выполняет и каталитическую функцию – гидролитическое отщепление фосфатного остатка АТФ (последнего – «на хвосте»). Освобождающаяся при этом энергия используется на совершение механической работы – укорочение участка мышечного волокна – сокращение. Сокращение вызывается «подтягиванием» белковых тяжей (пучков скрученных нитей) во встречном продольном направлении.
АТФ + миозин →→ миозин --- АТФ
Миозин --- АТФ →→ миозин + АДФ + Фнеорг + работа
На работу мышцы и, соответственно, образование мышечных комплексов «АТФ---миозин» расходуется АТФ, ее «рабочая» концентрация (некоторое время) поддерживается ресинтезом АТФ за счет запаса креатинфосфата в мышечной ткани (и, чем больше запас, тем большее можно развить усилие).
В период отдыха запас креатинфосфата восстанавливается: он образуется в обратной реакции АТФ + креатин.
И это еще не все. По теме лекции №2 на лабораторных, практических занятиях студенты изучают следующие вопросы:
Образование АТФ в процессах сопряженного окисления.
Субстратное фосфорилирование.
Окислительное фосфорилирование.
Разобщение дыхания и фосфорилирования.
Митохондрии их основная функция.
Работа митохондрий в мышечных волокнах.
Лекция № 3