


133
Глава 7. Регуляция биосинтеза белка
7.1Регуляция экспрессии генов у прокариот
АОбщая характеристика
Регуляция экспрессии генов у прокариот осуществляется в основном на уровне транскрипции. Возможно, это связано с тем, что прокариотические мРНК имеют короткий «срок жизни» — всего несколько минут.
В большинстве случаев прокариоты используют негативную регуляцию с помощью ген-специфичных репрессоров, которые связываются с промоторами структурных генов. Все белки прокариот (в т.ч. и ферменты) можно разделить на конститу-
тивные и регулируемого синтеза.
Конститутивные белки синтезируются постоянно в любых условиях. Их количество не изменяется в процессе жизнедеятельности клетки или при изменении условий окружающей среды. Клетка «считает», что эти ферменты будут нужны ей всегда, поэтому кодирующие их гены постоянно экспрессируются («включены»). Пример: ферменты, метаболизирующие глюкозу.
Клетка затрачивает значительные ресурсы на синтез белков, поэтому ей не выгодно синтезировать белок, если в данный момент он ей не нужен. Обеспечить экономию позволяет наличие белков регулируемого синтеза. Клетка может изменять скорость их синтеза и концентрацию (иногда в тысячи раз) в соответствии со своими нуждами. Таким образом, синтезируются только те белки (ферменты), которые необходимы клетке в данных условиях. Белки регулируемого синтеза можно разделить на две группы:
1.В случае индуцибельных белков регуляция направлена на «включение» синтеза ранее не требовавшегося фермента, когда в нем возникла необходимость. «Включение» синтеза белка называется индукцией, а «включающее» синтез химическое вещество — индуктором (или депрессором). Индуктор реагирует с репрессором, инактивируя его. Как правило, индуцибельными являются ферменты катаболических путей (сбраживание сахаров, распад аминокислот и др.).
2.В случае репрессируемых белков цель регуляции — прекращение (репрессия) их синтеза. Обычно репрессируемыми являются ферменты анаболизма (синтеза аминокислот, азотистых оснований и т.д.).
БИндукция. Лактозный оперон
Лактозный оперон (lac-оперон) — это оперон кишечной палочки (E. coli), содержащий три структурных гена lacZ, lacY, lacA, которые кодируют 3 белка: β -галак-
тозидазу, галактозидпермеазу, тиогалактозид-трансацетилазу. Эти белки участвуют в усвоении лактозы кишечной палочкой: они транспортируют её в клетку и расщепляют на глюкозу и галактозу, которые далее включаются в гликолиз.

134 Глава 7 Регуляция биосинтеза белка
Оперон может находиться в двух состояниях: активном (включён) и неактивном (выключен). В норме (при отсутствии лактозы в среде) лактоз-
ный оперон неактивен (выключен):
промотор оперона блокирован lac- репрессором — этот белок препятствует транскрипции, поскольку не даёт РНК-полимеразе возможности корректно начать её (связаться с промотором полимераза всё-таки может, как было показано в последние годы)
(см. Рис. 88 ).
Как только лактоза появляется в среде, она попадает в клетку и связывается с lac-репрессором. Это меняет его конформацию, он становится неактивен, утрачивает своё сродство к промотору и отщепляется. Lac-оперон включается и на нём начинается транскрипция, а затем происходит трансляция мРНК на белок (см. Рис. 89 ). Синтезированные ферменты помогают клетке «усвоить» лактозу:
1.Галактозидпермеаза служит мембранным переносчиком лактозы в клетку (ускоряет её перенос в разы);
2.β-Галактозидаза расщепляет лактозу до глюкозы и галактозы, которые затем включаются в гликолиз.
3.Тиогалактозид-трансацетилаза переносит ацетильную группу с аце- тил-КоА на β-галактозиды. Её функция в контексте lac-оперона ещё недостаточно изучена. Есть гипотеза, что этот фермент обезвреживает неметаболизируемые пиранозиды, ацетилируя их.
Вэтом механизме регуляции — индукции — лактоза является индуктором, в отсутствии неё репрессор выключает транскрипцию на опероне. Такую регуляцию называют негативной.
Рис. 89. Активация лактозного оперона: лактоза появляется в среде, связывается с репрессором, выключает его, начинается транскрипция и трансляция lac-оперона.

Регуляция экспрессии генов у прокариот 135
В Катаболическая репрессия. Лактозный оперон
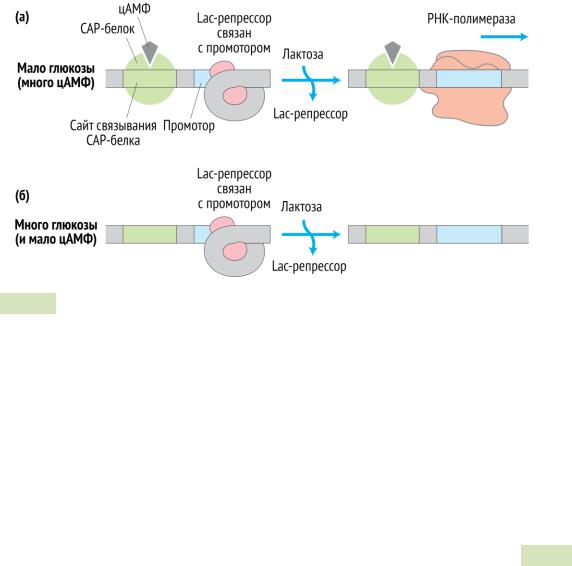
Помимо индукции, у кишечной палочки существует механизм позитивной регуляции — катаболическая репрессия (см. Рис. 90 ▼). Клетки кишечной палочки
E.coli способны усваивать как глюкозу, так и лактозу. Возможны два случая:
1.В среде есть глюкоза и лактоза. Глюкозы вполне достаточно, чтобы снабжать клетку энергией, поэтому в усвоении лактозы нет необходимости. Поэтому транскрипция на lac-опероне должна быть выключена, чтобы клетка не тратила ресурсы. Такой эффект достигается с помощью осо-
бого белка — CAP (англ. catabolite gene activator protein). CAP связыва-
ется с цАМФ и образует комплекс CAP + цАМФ. Этот комплекс присоединяется к промотору lac-оперона и помогает РНК-полимеразе начать транскрипцию. Однако, если в клетке достаточно глюкозы, концентрация цАМФ низкая12, комплекс CAP + цАМФ не образуется и активации транскрипции не происходит.
2.В среде есть только лактоза. Концентрация цАМФ повышается, он связывается с CAP-белком и образует активный комплекс CAP + цАМФ. Комплекс присоединяется к промотору и «усиливает» его. Транскрипция на lac-опероне начинается. Синтезируемые белки усваивают лактозу.
12Концентрация цАМФ у кишечной палочки E. coli регулируется с помощью фосфотрансферазной системы. Она служит для переноса моносахаридов в клетку и их фосфорилирования (это препятствует обратному выходу сахара из клетки, поскольку фосфорилированные моносахариды имеют отрицательный заряд и не «пропускаются» клеточной мембраной). Если глюкозы много, белок EIIA фосфотрансферазной системы находится в дефосфорилированном состоянии (остаток фосфорной кислоты переносится с него на глюкозу, в результате сам белок дефосфорилирован) и ингибирует аденилатциклазу (синтезирующую цАМФ) и галактозидпермеазу (транспортёр лактозы в клетку). Если глюкозы много, белок EIIA фосфорилирован и не может ингибировать эти ферменты. Концентрация цАМФ в таком случае вырастает.
136 |
Глава 7 |
Регуляция биосинтеза белка |
Рис. 90. Схема катаболической репрессии (слева — до появления лактозы, справа
— после её появления). А) Если глюкозы в клетке мало, концентрация цАМФ высока. Молекулы цАМФ связываются с CAP-белком, образуя комплекс CAP + цАМФ. Этот комплекс присоединяется к промотору и «усиливает» его. Транскрипция на lac- опероне начинается. Б) Если глюкозы в клетке много, концентрация цАМФ низкая. Комплексов CAP + цАМФ почти не образуется, и транскрипция на lac-опероне не идёт, поскольку промотор этого оперона «слабый», и сродство РНК-полимеразы к нему низкое.
ГРепрессия. Триптофановый оперон
Триптофановый оперон (trp-оперон) — это оперон кишечной палочки E. coli,
кодирующий 5 полипептидных цепей, которые затем образуют 3 белка (см. Рис. 91 ▼).
Эти белки синтезируют триптофан из хоризмовой кислоты:
1.Антранилатсинтаза;
2.N-(5’-фосфорибозил)-антранилатизомераза и индол 3-глицеролфос-
фатсинтаза (комплексный фермент с двумя активными центрами);
3.Триптофансинтаза.
Внорме триптофановый оперон включён: на нём идёт транскрипция, trp- репрессор синтезируется неактивным (в отличие от lac-репрессора, который после синтеза сразу активен). Как только белки, кодируемые trp-опероном, синтезируют избыток триптофана, его молекулы соединяются с trp-репрессором и активируют его. Комплекс trp-репрессор + триптофан связывается с оператором trp-оперона и снижается частоту транскрипции на нём примерно в 70 раз. Как только концентрация триптофана в среде снова снижается, trp-репрессор переходит в неактивное состояние, и транскрипция на trp-опероне возобновляется.
Вэтом механизме регуляции — репрессии — триптофан является корепрессором. Репрессия trp-оперона — другой пример негативной регуляции.