

Элонгация у эукариот 125
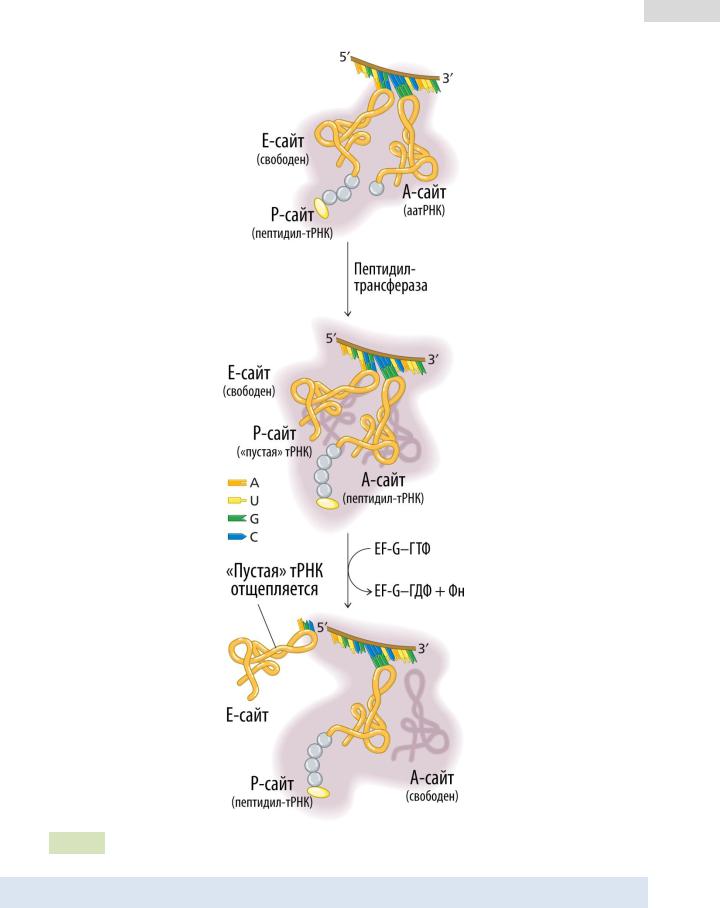
Рис. 84. Элонгация трансляции у прокариот: транспептидация и транслокация.
6.8 Элонгация у эукариот
Элонгация трансляции у эу- и прокариот очень схожа. У эукариот вместо фактора EF-Tu — EF-1α. Вместо EF-Ts — EF-1β. Аналогом EF-G прокариот является EF-2.
126 |
Глава 6 |
Трансляция |
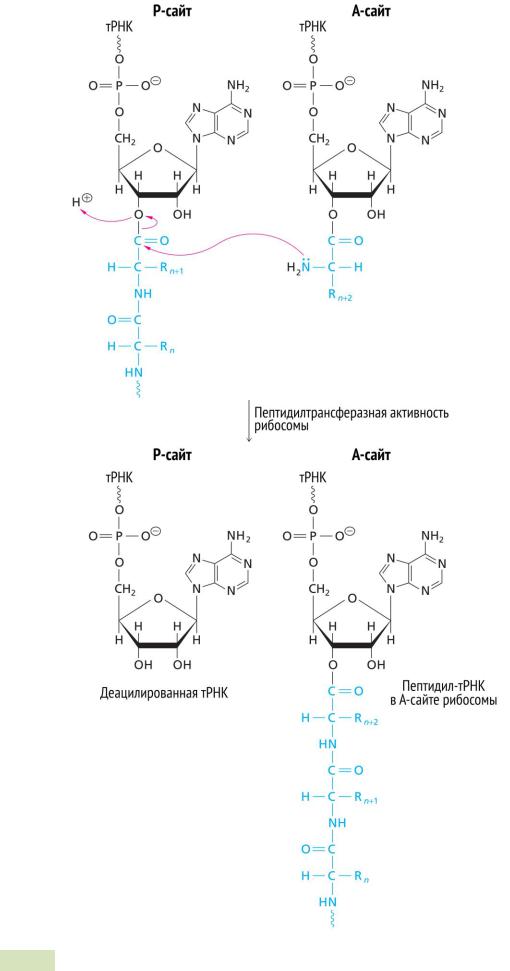
Рис. 85. Образование пептидной связи на этапе транспептидации (элонгация).
Терминация у прокариот 127
6.9 Терминация у прокариот
Терминация — это заключительная стадия трансляции, на которой прекращается синтез белковой цепи, происходит отщепление цепи и диссоциация рибосомы.
Этапы терминации:
1.Как только в А-сайте оказывается один из стоп-кодонов (UAG, UGA, UAA), с ним связывается релиз-фактор: RF-1 или RF-2. Он переносит пептидильную группу с тРНК (полипептидную цепь) на молекулу воды, вызывая её высвобождение.
2.Фактор RF-3 связывается с рибосомой в комплексе с ГДФ. Происходит замена ГДФ на ГТФ. Релиз-фактор RF-1/2 высвобождается. RF-3 гидролизует ГТФ до ГДФ и отщепляется от рибосомы.
3.К рибосоме присоединяется фактор RRF (ribosomal recycling factor) и EF-G + ГТФ. EF-G гидролизует ГТФ, благодаря чему RRF перемещается в P-сайт, а тРНК находящиеся в E- и P-сайтах, отщепляются.
4.Большая и малая субъединицы рибосом диссоциируют. В этом процессе участвует фактор IF-3.
6.10Терминация у эукариот
У эукариот всего один релиз-фактор — eRF-1 — связывается со всеми тремя стоп-кодонами и катализирует гидролиз связи между С-терминальной аминокислотой в синтезированной полипептидной цепи и тРНК. У эукариот существуют т.н. суппрессорные тРНК, позволяющие трансляции продолжаться даже при достижении стоп-кодона.
6.11Гипотеза «качания»
Вэлонгации трансляции выбор правильной тРНК осуществляется на основании кодон-антикодонового взаимодействия, т.е. аминоацильная группа тРНК в этом процессе не участвует (это одна из причин, по которой корректное аминоацилирование — критический этап синтеза белка). Три нуклеотида на мРНК (кодон) связываются с тремя нуклеотидами на тРНК (антикодон) антипараллельно.
Логично предположить, что 61 кодон, кодирующий аминокислоту, будет взаимодействовать с 61 антикодоном тРНК (и, следовательно, 61 молекулой тРНК). Однако на самом деле подавляющее большинство клеток содержит изоакцепторные тРНК, которые связываются с кодоном лишь двумя своими нуклеотидами. Пример: дрожжевая тРНКФен, содержащая антикодон GmAA (Gm — 2’-метилированный гуанозин), «узнаёт» кодоны UUC и UUU (см. Рис. 86 ▼).

128 |
Глава 6 |
Трансляция |
Рис. 86. Кодон-антикодоновое взаимодействие между дрожжевой мРНК и тРНКФен
Кроме того, дрожжевая тРНКАла, имеющая антикодон IGC (I — минорное азотистое основание инозин), «узнаёт» кодоны GCU, GCC, GCA (см. Рис. 87 ▼):
Рис. 87. Кодон-антикодоновое взаимодействие между дрожжевой мРНК и тРНКАла
Обратите внимание, что на 5’-конце антикодона тРНК в обоих случаях находится модифицированное азотистое основание (в первом случае — метилированный гуанин, а во втором — инозин). Очевидно, именно этот третий нуклеотид антикодона может не образовывать Уотсон-Криковскую пару с третьим нуклеотидом кодона мРНК.
Рассмотрев структурные особенности вышеописанных пар, Крик предложил гипотезу «качания», чтобы объяснить, как тРНК способна «узнавать» несколько кодонов. Он предположил, что первые две пары нуклеотидов в кодоне и антикодоне имеют нормальную Уотсон-Криковскую комплементарность, а в третьей паре нуклеотидов происходит некое «качание», способствующее правильной геометрии всего кодон-антикодонового спаривания. Это объясняет образование не-Уотсон-Криков- ских пар: U • G и I • A. Другие варианты приведены в таблице ниже.
|
Таблица 11 |
|
Возможные не-Уотсон-Криковские пары в кодон-антикодоновом |
|
|
|
взаимодействии |
|
|
|
|
|
|
|
|
|
|
|
|
|
5’-антикодоновое |
|
3’-кодоновое |
|
|
основание |
|
основание |
|
|
|
|
|
|
|
C |
|
G |
|
|
|
|
|
|
|
A |
|
U |
|
|
|
|
|
|
|
U |
|
U или G |
|
|
|
|
|
|
|
G |
|
G или C |
|
|
|
|
|
|
|
I |
|
U, C или A |
|
|
(инозин) |
|
|
|
|
|
|
|
|
|
|
|
|
|