

Кординація діяльності ЦНС. Координація діяльності всієї ЦНС підлягає певним принципам і зв’язана з різними засобами встановлення зв’язків між нейронами.
1)Визначають 1)лінійні 2)конвергентні 3)дивергентні 4)реверберуючі ланцюги нейронів
1)Лінійні ланцюги забезпечують передачу інформації без перешкод і трансформації
2)Конвергентні ланцюги забезпечують посилення інформаційного сигналу, його концентрацію у нервових центрах та обмеження об’єму інформації, що йде таким ланцюгом
3)Дивергентні ланцюги обумовлюють посилення інформаційного сигналу, надійність його передачі за рахунок дублювання інформаційних каналів, детальний аналіз інформації; вони являються морфологічною основою іррадіації збудження, а також реціпрокних зв’язків
4)Реверберуючі ланцюги також здатні забезпечити посилення сигналу й збільшення тривалості його існування, але, якщо у такий ланцюг є вбудованим гальмівний нейрон, він забезпечує зворотне гальмування
2)До принципів координації діяльності ЦНС належать: 1)принцип боротьби за кінцевий шлях 2)принцип загального кінцевого шляху Шеррингтона 3)принцип концентрації та іррадіації збудження у ЦНС 4)принцип оклюзії та полегшення (фасилітації) проведення збудження у ЦНС 5)принцип індукції у ЦНС 6)принцип виникнення і панування домінанти (Ухтомського)
1)Принцип боротьби за кінцевий шлях полягає у тому, що організм, як правило, сприймає не одне, а декілька подразнень, кожне з яких може претендувати на реагування з боку організму, що зумовлює конкуренцію між ними. Але унаслідок аналітичної діяльності ЦНС відбувається вибір найбільш суттєвого подразнення, що потребує рефлекторної відповіді. Цей вибір здійснюється не тільки за критерієм сили діючого подразника, але й за поточним функціональним станом всього організму й ЦНС
2)Принцип загального кінцевого шляху Шеррингтона полягає у тому, що кінцева частина різних рефлекторних дуг може бути однаковою. Наприклад, якщо людина вбігає у річку і наштовхується ногою на скло – нога, що пошкоджена миттєво згинається, друга, навпаки, випрямляється і людина утримує рівновагу й не тоне. Або тренер звертається до спортсмена із проханням зігнути одну ногу і випрямити іншу у вертикальному положенні. В обох випадках ми маємо однаковий результат, але зовсім різні дуги рефлексів.
В рамках відносно простої рефлекторної дуги у 1 випадку спрацьовує задана генотипом морфологічна детермінанта – дивергенція аферентного входу, що автоматично включає так звані реципрокні відносини, тобто спряженні різноспрямовані рефлекторні реакції. В рамках багаторівневої дуги рефлексу у 2 випадку спрацьовують нейронні ансамблі, що поєднані як зв’язками, що задані генотипом, так і набутими у процесі життя зв’язками між нейронами. В першому випадку ми маємо справу з відносно простим рефлекторним актом, у другому – з прикладом вже достатньо складного поведінкового акту саме за способом його нервової регуляції.
3)Принцип концентрації та іррадіації збудження у ЦНС полягає у зосередженні збудження у певних нервових центрах, або його розповсюдженні по ЦНС. Це відбувається на підставі відповідних ланцюгів нейронів – завдяки наявності й збалансованості процесів збудження й гальмування у ЦНС та аналітичній, інтегративній діяльності кожного нейрону. Феномен іррадіації збудження по нервовим центрам є протилежним концентрації і може відбуватися при порушенні збалансованості між процесами збудження і гальмування – унаслідок занадто сильного подразнення, що зумовлює дуже велику силу збуджувальних процесів, яким не може протистояти гальмування; або при послабленні сили гальмування у ЦНС
4)Принцип окклюзії при проведенні збудження у ЦНС полягає у тому, що сумарна відповідь на кілька подразнень, у наслідок перекриття нервових шляхів, що передають інформацію у відповідні центри, опиняється меншою, ніж очікувана як арифметичний підсумок відповідей на кожне окреме з цих подразнень. Принцип полегшення (фасилітації) проведення збудження є протилежним оклюзії і полягає у збільшенні сумарної відповіді, у порівнянні з очікуваною, унаслідок підключення до неї підпорогових зон нервових шляхів, в яких під впливами кількох подразнень реакція досягає порогового значення

5)Явище пост-тетанічної потенціації – посилення відповіді нейронів під впливом попередньої високочастотної ритмічної (тетанічної) стимуляції. Має спільну основу з феноменом післядії – припинення реакції нейрону відбувається значно пізніше, ніж дія подразнення, що його викликало. Відносно короткочасна післядія може бути наслідком слідової деполяризації постсинаптичної мембрани. Тривала післядія є наслідком циркуляції збудження по замкненим ланцюгам нейронів (на підставі кільцевого, реверберуючего ланцюга). Циркуляція збудження по таким ланцюгам може продовжуватися до тих пір, поки не відбудеться втома одного з синапсів або не виникне гальмування. При такій циркуляції збудження зміни стану нейронів аналогічні змінам при тетанічній(зовнішній) стимуляції. Отже, може виникати післядія на зразок пост-тетанічної потенціації. Це переважно обумовлено зміною властивостей пресинаптичної мембрани, яка набуває здатності виділяти більші порції медіатору у відповідь на кожний новий ПД – частково тому, що в синаптичній щілині зберігається залишки попередньої порції. А також завдяки синапсам на пресинаптичних терміналях, які виділяють відповідні медіатори і потенцюють збудження /наприклад, зменшують проникність для іонів калію, що веде до збільшення тривалості ПД і, відповідно, виділення медіатору/
6)Принцип реципрокності і синергічності рефлекторних реакцій. З аферентним гальмуванням щільно зв’язані так звані реципрокні взаємовідносини (реципркність) – спряжені (на підставі дивергенції аферентного входу), різноспрямовані (збудження-гальмування) рефлекторні реакції, які викликаються одним і тим же подразником і забезпечуються антагоністичними рефлекторними центрами (наприклад, на іпсі-латеральному подразненню боці стимуляція мотонейрону згинача супроводжується гальмуванням мотонейрону розгинача, а на контр-латеральному боці, навпаки, гальмується мотонейрон згинача). Тому аферентне гальмування часто визначають як реципрокне (наприклад, 2-бічна стимуляція рецепторів шкіри шкідливими, больовими факторами супроводжується взаємним гальмуванням мотонейронів згиначів з обох сторон).
Слід зазначити, що не завжди включення 2-х (або більш) аферентних каналів супроводжується взаємним гальмуванням рефлекторних реакцій, які вони викликають. Дивергентний аферентний вхід поєднує і синергічні (спів-дружні) центри /принцип синергічності – протилежний за суттю принципу реципрокності/ (наприклад, на контр-латеральному боці від стимульованого мотонейрону згиначеві стимулюється також мотонейрон розгиначеві). У такому випадку включення (2-х або більш) аферентних каналів призводить до взаємного посилення рефлекторних реакцій, що відбуваються (наприклад, стимуляція м’язових веретен /пропріорецепторів/ розгинача на одному боці і шкірних больових рецепторів на іншому боці може привести до посилення як скорочення розгинача /з боку подразнення пропріорецепторів/, так і до посилення згинання /на протилежному боці, де подразнювалися больові рецептори/).
7)Принцип індукції полягає у тому що у кожному нейроні або нервовому центрі один нервовий процес може переходити у свою протилежність. Зміну збудження на гальмування визначають як негативну індукцію. І, навпаки, якщо гальмування змінюється на збудження індукція має назву позитивної. При одночасній індукції навколо осередку збудження формується гальмівна зона, а осередок гальмування оточується зоною збудження.
Якщо індукція виступає як перехідний процес у часі, то після збудження у певному осередку нейронів відбувається гальмування, й навпаки – після гальмування – збудження. Цей феномен, зокрема, лежить у основі так званих ритмічних рефлексів, наприклад чесання.
8)Принцип виникнення і панування домінанти (Ухтомського) полягає у тому, що у певних функціональних умовах збудливість певних нервових центрів може різко зростати, що надає таким центрам властивостей домінантного осередку у ЦНС, якому є притаманні: а)здатність до виникнення сильного та інертного збудження б)здатність до „перетягування” збудження з „чужих” рецепторних ланок, тобто якщо, раніш такий центр не реалізовував у нервовий імпульс незначні за амплітудою ЗПСП, що доходили до нього з різних ділянок нервової системи, то, завдяки високій збудливості, став реагувати на них імпульсом (ПД) в)здатність до пригнічення будь-яких інших рефлекторних відповідей (навіть, якщо вони адекватні даному подразненню), замість яких домінантний рефлекторний центр нав’язує виключено „свою” відповідь. У таких умовах на більшу кількість зовсім різних подразнень відбувається стереотипна рефлекторна реакція, що зумовлюється домінантним осередком ЦНС.
ЦНС
Збудження у ЦНС відбувається у 2-х видах:
1.Місцеве збудження – ЗПСП (виникає на сомі та дендритах нейрону)
2.Генералізоване збудження – ПД (виникає на аксонному горбику і поширюється по аксону
1.Місцеве збудження викликає ПД, що приходить до синаптичної бляшки, встановленої на сомі або дендритах нейрону. ПД зумовлює вхід туди Са++. Це забезпечує вихід медіатору у синаптичну щілину. Медіатор зв’язується з мембранним рецептором соми або дендритів → посилюється потік іонів натрію у середину нейрона. Але викликана деполяризація залишається місцевою, так як на сомі і дендритах мало потенціал-залежних натрієвих каналів і багато хемо-залежних натрієвих каналів (завдяки явищу конвергенції – сходженню на тілі і коротких відростках нейрону багатьох синаптичних входів). Отже ΔЕ (поріг збудження) там досить великий, а збудливість, відповідно низька.
збуджувальний нейрон
↓
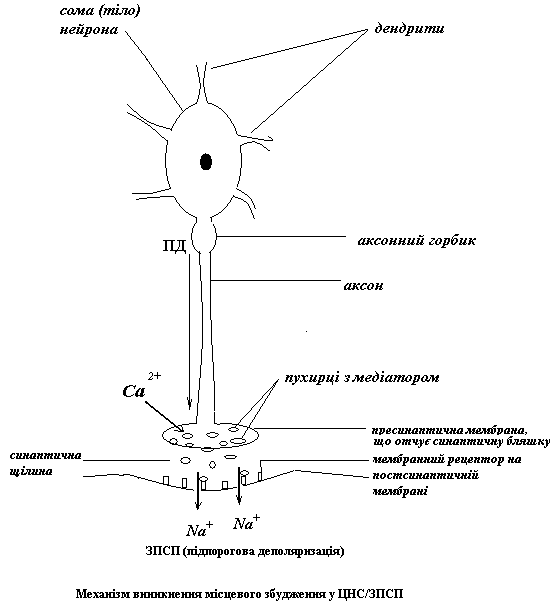
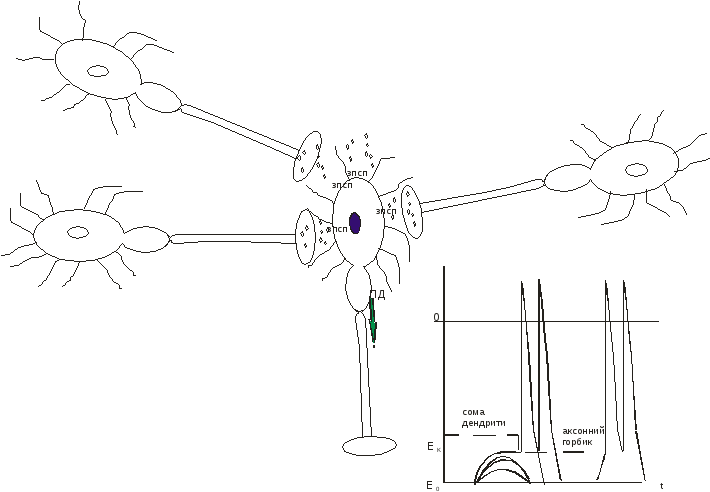
2.Генералізоване збудження – ПД – виникає на аксонному горбику (АГ) зазвичай унаслідок сумації ЗПСП. У зоні АГ, навпаки, багато потенціал-залежних натрієвих каналів і маленький ΔЕ (висока збудливість). ЗПСП – мають досягти АГ, а оскільки місцеві відповіді, віддаляючись від місця виникнення, втрачають амплітуду, одному ЗПСП це не по силам. Отже, вони мають підтримати один одного.
Якщо розширюється рецептивне поле, що зараз подразнюється (збільшується кількість збуджених одночасно рецепторів), то збільшується кількість збуджених аферентів, що сходяться на одному нейроні. Відповідно, збільшується кількість ЗПСП, що виникають на цьому нейроні одночасно. Ці ЗПСП підтримують один одного на шляху просування деполяризації до АГ. Отже, деполяризація не «згасає», а набуває здатності досягти АГ і зсунути там Е0 до Екр. Така сумація називається просторовою/одночасною, і передумовою для її здійснення є збільшення одночасно збуджених рецепторів.
Зазвичай на нейроні відбувається просторова сумація і ЗПСП, і ГПСП. Алгебраїчний підсумок цих протилежних явищ є вирішальним для того – чи буде в АГ ПД.
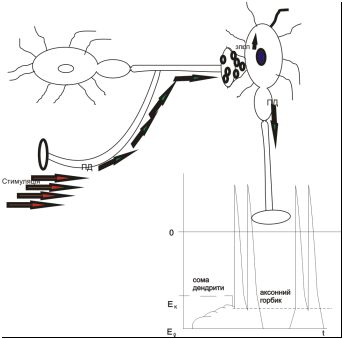
Механізм просторової сумації ЗПСП
Якщо один і той же аферент підлягає частій стимуляції, то медіатор, що виділився у синаптичну щілину, не встигає усунутися. І кожна нова порція медіатору накладається на залишки попередньої. Це зумовлює збільшення зв’язків медіатор+рецептор → збільшується інтенсивність потоку іонів натрію у середину нейрону. Як результат – зростає амплітуда ЗПСП у даному синаптичному контакті. Така сумація називається часовою/послідовною. Її передумова – часта стимуляція одного синаптичного контакту.
Механізм часової сумації
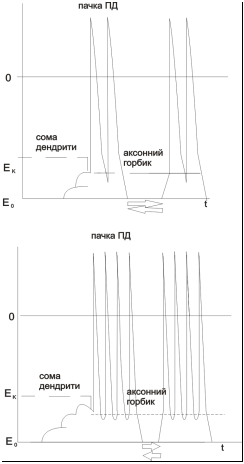
ПД, що виникає у АГ і розповсюджується по аксону, не буває поодиноким. Він існує у вигляді розряду багатьох нервових імпульсів (ПД). При цьому, незалежно від результату ефективної сумації(порогова чи надпорогова деполяризація у АГ), амплітуда ПД залишається сталою. Однак, в залежності від цього результату, змінюється частотний розподіл ПД у розряді. Порогова деполяризація у АГ – рідкі пачки імпульсів з малою кількістю ПД у окремій пачці. Чим більш надпорогова деполяризація у АГ – тим більш частота слідування пачок і кількості ПД у кожній окремій пачці.
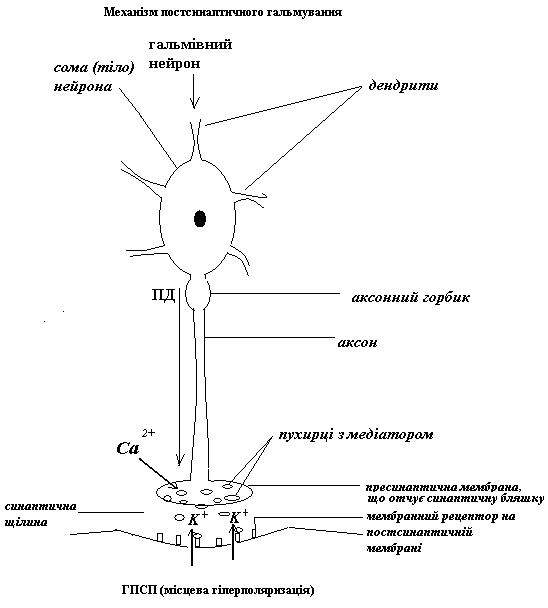
Найбільш поширений варіант механізму ГПСП
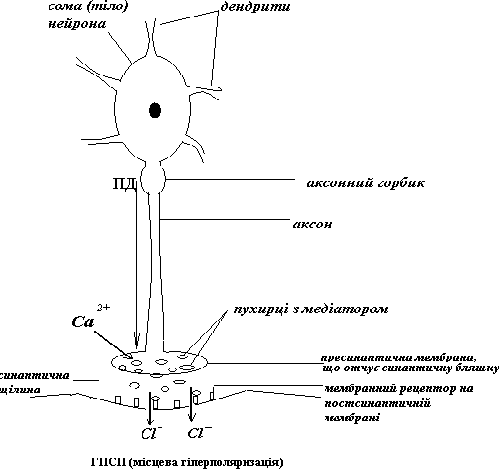
Можливий варіант механізму ГПСП
Отже, ГПСП заважає просуванню до аксонного горбику всіх тих ЗПСП, на шляху у яких стоїть гальмівний нейрон, що його викликав. Особливо ефективний такий гальмівний синапс поблизу від АГ. Пресинаптичне ж гальмування впливає лише на один ЗПСП – той, що викликає аксон збуджувального нейрону, з яким утворений аксо-аксональний синапс.
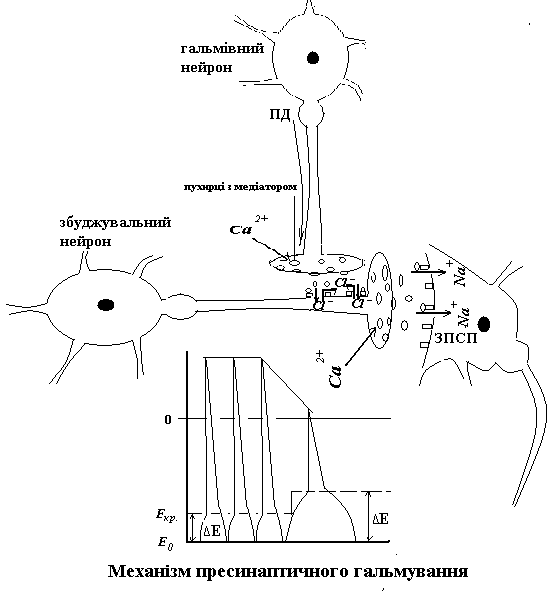
Гальмівний медіатор (зокрема, ГАМК)+рецептор → вхід С1- у середину аксона → С1- «тягне» МПС вниз (гіперполяризація), а лавиноподібний вхід іонів натрію під час ПД, що йде по аксону збуджувального нейрону, відповідно, вверх → підпорогова деполяризація замість ПД і часткова інактивація натрієвих каналів у регіоні пресинаптичної мембрани збуджувального синапсу → зменшення амплітуди наступного ПД, що прямує до пресинаптичної мембрани збуджувального синапсу (або повна його блокада) → зменшення (або припинення) входу Са++ у синаптичну бляшку → зменшення (або припинення) виходу збуджувального медіатору → зменшення (або зникнення) ЗПСП.
Рефлекси спинного мозку, що починаються від м’язових веретен
Увага! Це єдині рецептори, від яких починається моносинаптична рефлекторна дуга.
-
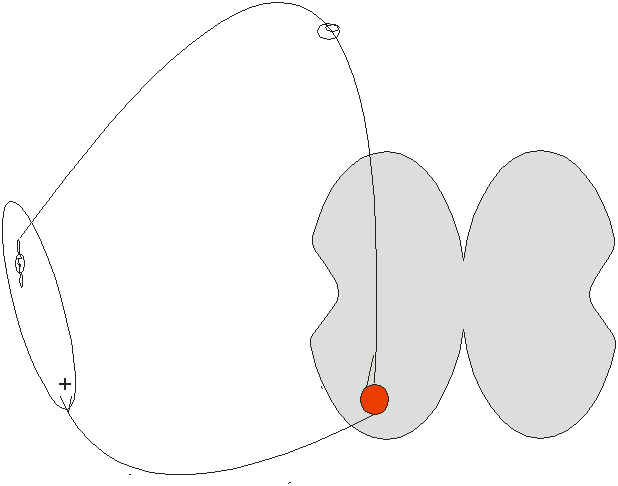
Міотатичний рефлекс – забезпечується на спінальному рівні – формується відносно слабке тонічне скорочення м’язу:
розслаблення м’язового волокна → розтягнення спіралі м’язового веретена і його збудження → збудження аферентного волокна і передача збудження на ά-мотонейрон → скорочення цього волокна и припинення збудження веретена;
під час скорочення одного волокна інше волокно – розслаблюється і започатковує таким самим образом збудження. Отже, одні волокна розслаблюються і започатковують збудження, інші – скорочуються і збудження припиняється – як результат слабке тонічне скорочення
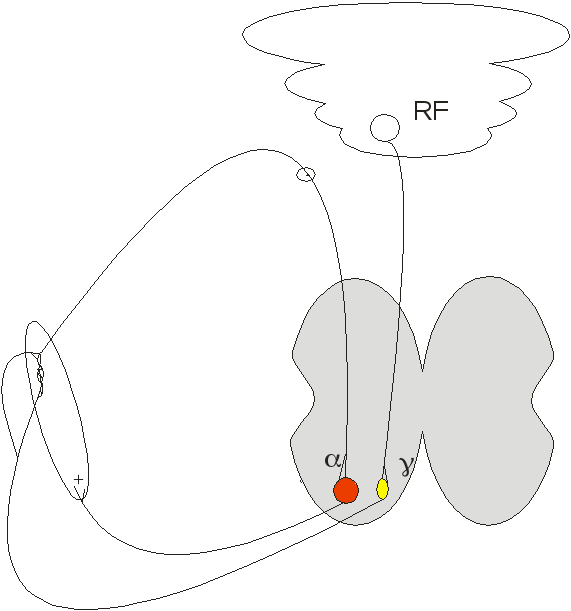
При γ-стимуляції викликається скорочення скоротливих елементів ядерної сумки м’язового веретена → спіраль веретена розтягується навіть на фоні скороченого м’язового волокна. Отже, формується «вічний двигун» - хоч розслаблене волокно, хоч скорочене, а веретено збуджене і, відповідно, ά-мотонейрон теж. Це значно посилює інтенсивність тонічного скорочення. Але на спінальному рівні γ-стимуляція майже не здійснюється. Для цього потрібні стовбурні структури головного мозку, зокрема, ретикулярної формації.
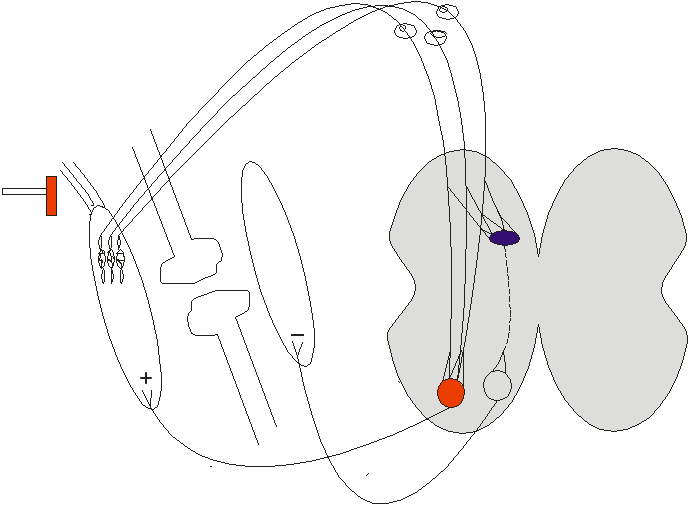
2)Сухожилковий рефлекс
Студент(ка), увага! у назві – підстава! Рефлекс починається від м’язових веретен (а ні сухожилкових рецепторів Гольджи)
Удар по сухожилку викликає сильне розтягнення найближчої ділянки м’язу і одночасне розтягнення кількох м’язових веретен. Отже, кілька веретен збуджується синхронно. Це зумовлює сильне збудження ά-мотонейрону вдареного м’язу і адекватне реципрокне гальмування ά-мотонейрону антагоністичного м’язу – вдарений м’яз різко скорочується, а його антагоніст розслаблюється. Скорочення набуває фазичного, а ні тонічного характеру.
Доречи, скорочуватися таким чином може не тільки екстензор (як прийнято вважати у студентському середовище), але і флексор, якщо вдарили по сухожилку флексора.
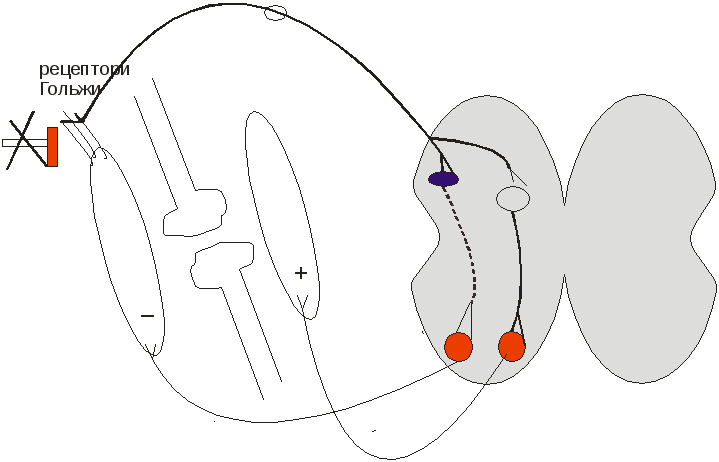
Сухожилкові рецептори Гольджи започатковують гальмування м’яза, якщо він сильно скоротився (рецептор Гольджи → гальмівний нейрон → гальмування ά-мотонейрону сильно скороченого м’язу
↓
→ збуджувальний нейрон → збудження ά-мотонейрону його антагоніста).
Це може пояснювати 2 фазу сухожилкового рефлексу.
Реципрокні механізми спинного мозку при збудженні шкірних рецепторів
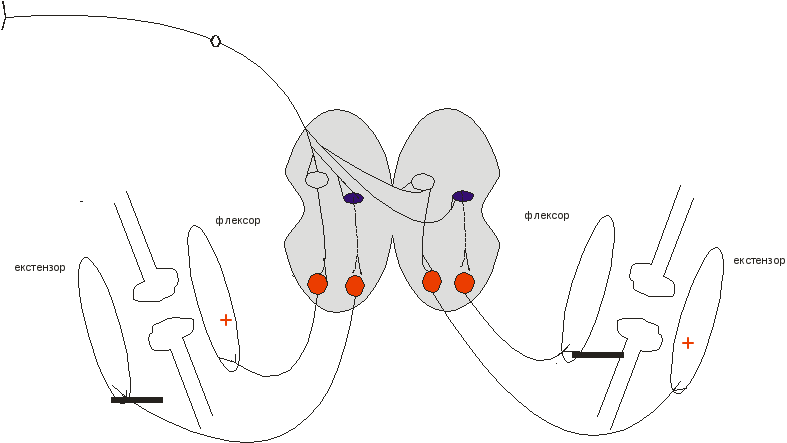
І.Стовбурно-спінальна регуляція м’язового тонусу (найпростіша схема)
Якщо уявити перерізку під червоним ядром (ЧЯ), можна пояснити механізм децеребраційної ригідності – зникнення стимулюючих впливів на флексори і гальмуючих на екстензори).
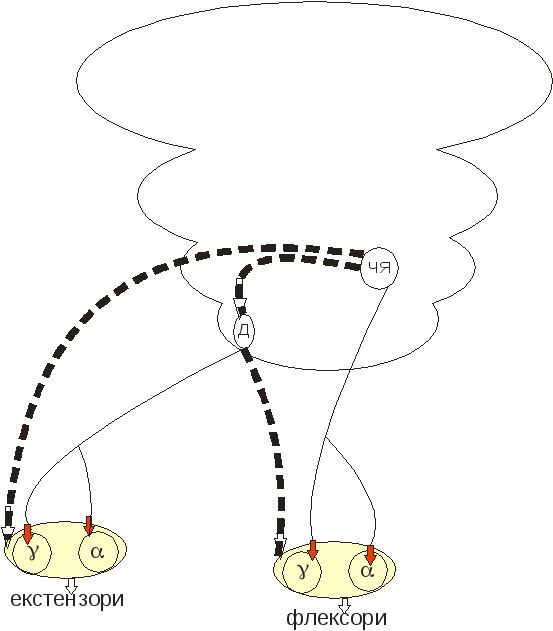
ІІ.Стовбурно-спінальна регуляція м’язового тонусу (трохи складніша схема)
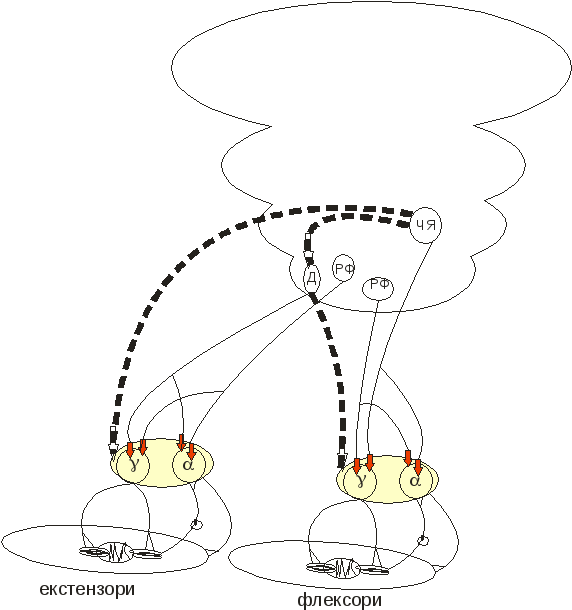
З цієї схеми можна бачити, що перерізка аферентних шляхів до α-мотонейронів зробить марними зусилля γ-стимуляції. Таким чином можна послабити прояви децеребраційної ригідності.
Організація
«входів» на мозочок і виходів його на
ядра стовбуру та підкоркові структури.
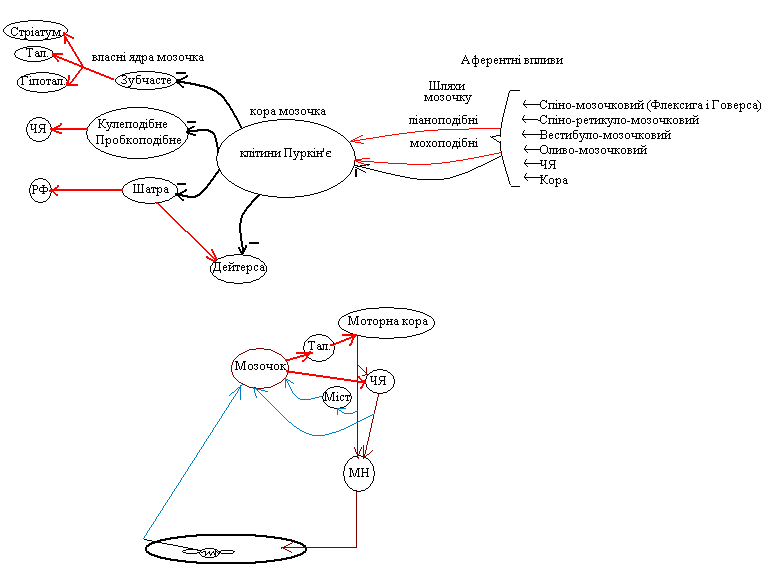
Мозочок отримує інформацію про команди до мотонейронів м’язів, які віддає кора і червоне ядро і про те, як ці команди виконуються м’язами (від стукачів-пропріорецепторів). Тому він надсилає «поправку до цих команд к моторній корі і к червоному ядру. Це пояснює симптомокомплекс при враженні мозочку.
Базальні ганглії переднього мозку – стріапалідарна система – включають бліду кулю і хвостате ядро, що разом з корою і структурами середнього мозку забезпечують регуляцію точних рухів (ця система має дуже високий розвиток у птахів).
Чорна речовина середнього мозку зв’язана з хвостатим ядром і шкаралупою ДОФАмінергічними нейронами. Хвостате ядро і шкаралупа передають інформацію до блідої кулі і чорної речовини за допомогою ГАМК. Від кори на базальні ганглії надходять переважно стимулюючі холінергічні (або за допомогою глутамату) впливи.
Цикл шкаралупи
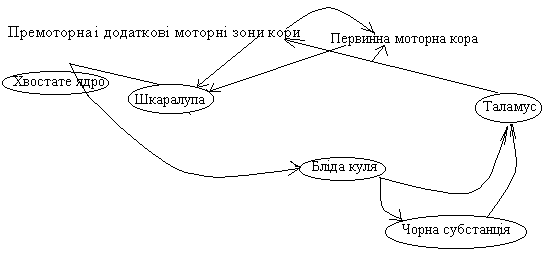
Цикл забезпечує разом з моторними зонами кори складні рухи.
Цикл хвостатого ядра
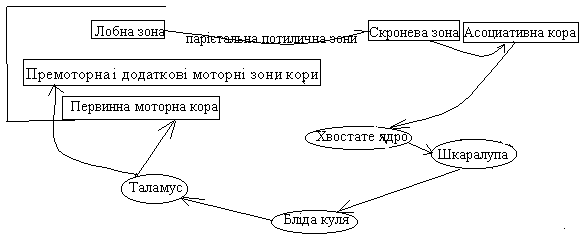
Цикл регулює ціліспрямовані моделі рухів (наприклад, поведінка при зустрічі з небезпечним звіром)
Ураження шкарклупи, хвостатого ядра – гіперкінезія (+гіпотонус) – тремор спокою і тремор довільного руху (хвороба Паркінсона).
Хвороба Паркінсона зв’язана з дефіцитом дофаміну (причиною може бути дисфункція чорної субстанції)
Ураження блідої кулі – гіпокінез (+гіпертонус – „воскові фігури”)
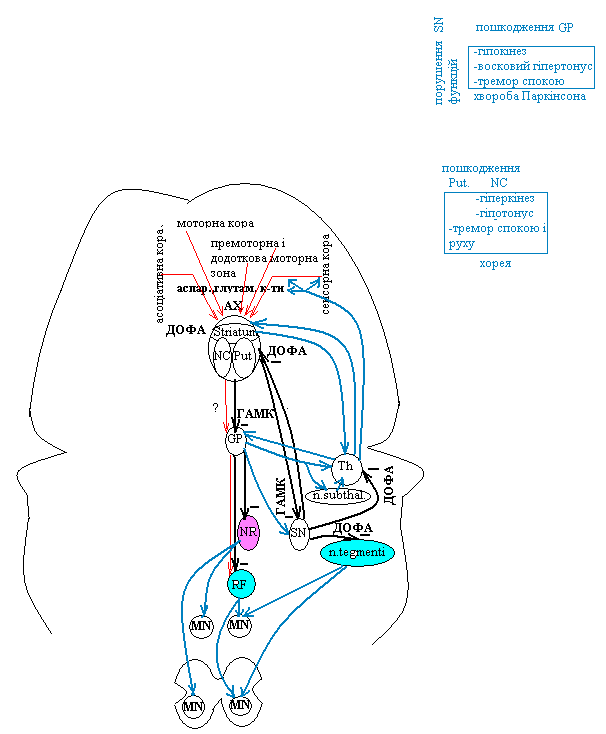
Таламус
|
Джерело інформації/Провідні шляхи |
Вид ядра таламусу, що її сприймає |
Коркова зона, яка сприймає інформацію від таламусу |
|
|
Специфічні |
|
|
|
Перемикаючі |
|
|
|
СЕНСОРНІ |
|
|
Всі екстер-, частково пропріо- й вісце- рорецептори (але не нюхові)/інформація: 1)від тактильних,пропріо-, вестибуло- (частково, вісцеро-) рецепторів по волок нам Аa,b,g йде до ядер Голля і Бурдаха (і головного сенсорного ядру Vпари;VІІІ), а потім перехрещується на рівні олив і прямує у PV у складі медіальної петлі (лемнисковий тракт) 2)від рецепторів тиску, термо- й ноцицеп- торів (екстеро- і вісцеро-)по волокнам Аd йде до сенсорних ядер заднього рогу (і ядра спінального тракту V пари),прямує на протилежний передній ріг і у складі неоспинно-таламічного (вентро-латераль- ного)тракту йде до PV |
Поствентраль- ний комплекс (медіобазаль- ний) – PV/MB/ |
Сомато-сенсорна зона: 1)постцентральна звивина (тім’яна зона) S1 2)латеральна борозна,що відокремлює тім’яну і скроневу кору S2 3)зона під ролландовою бороздою у верхнього краю сильвієвої борозди 4)поблизу від сильвієвої та циркулярної борозд – дода- ткова смакова зона
|
|
Зорові рецептори, від яких інформація прямує по аферентам ІІ пари (зоровий нерв) і специфічному зоровому шляху |
Латеральні колінчаті тіла LKT |
Шпорна борозна потиличної кори (стріарна зона,первинна зорова кора–V1/17поле)+вторинні V2-4 |
|
Слухові рецептори, від яких інформація прямує по аферентам VІІІ пари (слуховий нерв) і специфічному слуховому шляху |
Медіальні колінчаті тіла MKT |
Звивина Гешле(поперечні скроневі звивини верхньої частини скроне- вої кори–первинна слухова кора /41поле/ й вторинна зона /42 п/ |
|
Вісцеральні рецептори |
ЛІМБІЧНІ V |
Лімбічна кора, лімбічна система |
|
Вентральний комплекс |
||
|
Пропріо-, вестибулорецептори,від яких ін-- формація прямує відповідно у задній ріг і вестибулярні ядра(VІІІ пари) довгастого мозку, звідки у складі спинно- і вестибу- ломозочкових трактів йде до VL |
РУХОВІ VL |
Моторна кора:1)прецентральна звивина лобної кори 2)попереду від 1) – премоторна зона 3)додаткова зона на медіальній поверхні півкуль |
|
Вентро-латера- ний комплекс
|
||
|
Специфічні перемикаючі ядра та- ламусу і специфічні сенсорні й моторні зони кори
|
АСОЦІАТИВНІ |
1)Тім’яна асоціативна зона
2)Лобна асоціативна зона
3)Зорова, слухова асоціативні зони навколо зорової/латеральна зона Р/ й слухової /медіальна зона Р/ сенсорної |
|
1)Латеральний комплекс L 2)Медіо-дорзальний комплекс MD 3)Подушка Pulvinar (P)
|
||
|
Всі рецептори, від яких інформація по коллатералям прямує до РФ стовбуру мозку, звідки йде до неспецифічних ядер таламусу |
Неспецифічні |
Широкі зони кори, з якими встановлюється зв’язок через підкоркові синапси
|
КОРА.
Поверхня великих півкуль (кора) поділена бороздами, між котрими знаходяться звивини. У кожній півкулі виділяють по 4 основних долі: лобну, тім’яну, скроневу і потиличну. Клітинний склад кори відзначується морфологічно-функціональною неоднорідністю, що дозволяє користатися поняттям „локалізація функцій у КГМ”.
У філогенетичному та структурно-функціональному аспекті у корі великих півкуль відзначають неокортекс (нову кору) та стару лімбічну кору. Функції неокортексу певною мірою протиставляється лімбічній корі, що завершує лімбічну систему мозку (до якої надходять певні коркові та підкоркові утворення), й яка, у порівнянні з поверхнево розташованим неокортексом, має достатньо специфічне „приховане” розташування у мозку. Але функціональна специфіка цих видів кори заслуговує окремого розгляду і буде окремою темою.
Неокортекс підрозділяють на:
І)сенсорні зони, в яких здійснюється сприйняття і кінцевий аналіз поточної інформації, а саме:1)зорова потилична кора (шпорна борозна) 2)слухова скронева кора (звивина Гешле) 3)сомато-вісцеро-сенсорна тім’яна кора (постцентральна звивина, верхній відділ бокової борозни, додаткова зона у краю сильвієвої борозди, поруч з якою знаходиться смакова зона кори);
ІІ)сомато-моторні зони, що створюють рухову кору, яка забезпечує найбільш довершену регуляцію руху (лобна кора - прецентральна звивина та додаткові моторні зони); треба підкреслити, що моторні клітини кори – клітини Беца – напрямки зв’язані зі спінальними мотонейронами відповідних м’язів, що робить можливим здійснення довільних рухів;
ІІІ)асоціативні зони, які містяться навколо зорової, слухової кори і особливо відзначаються у лобній і тім’яній ділянках кори; асоціативна кора забезпечує впізнання, осмислення поточної інформації, й має виключено важливе значення у координації діяльності ЦНС та збереженні інформації у пам’яті
Якщо розглядати функціональну специфіку кори у вертикальній проекції, то слід виділити 6 більш менш однорідно організованих шарів нейронів, які постійно зустрічаються у неокортексі (лімбічна кора може бути у різних ділянках позбавлена деяких з цих шарів): 1.молекулярний 2.зовнішній гранулярний 3.зовнішній пірамідний 4.внутрішній гранулярний 5.внутрішній пірамідний 6.веретеновидний. Нижчі шари (4,5,6-й) у ділянках сенсорних і рухових зон – це первинні проекційні зони, що зв’язані аферентними та еферентними волокнами з периферійними органами. Над ними розташовані вторинні зони (2,3), що обумовлюють зв’язок з сусідніми корковими ділянками і окремими структурами стовбуру, проміжного мозку. Вторинні зони мають велике значення у поєднанні діяльності рухових і сенсорних зон мозку (переважне значення тут мають верхні шари нейронів). Над первинними і вторинними зонами розташовані третинні (1), що не мають безпосередніх зв’язків з периферією і виконують асоціативну функцію – забезпечують найбільш складну діяльність мозку, яка лежить у основі вищих психічних пізнавальних процесів (тут значущість верхніх шарів максимальна).
Відомо, що людина віддзеркалюється у сомато-сенсорній і сомато-моторній зонах у перевернутому виді у вигляді „гомункулюса” з великою головою, особливо її лицевою частиною, величезними кистями, стопами і маленькими тулубом, ручками і ніжками. Тобто, відносна кількість нейронів, що сприймають у корі інформацію від язику, обличчя набагато більше кількості клітин, до яких йде інформація, наприклад, від тулуба, так само, як і відносна кількість командних клітин моторної кори, що зумовлюють рухи, наприклад, кисті набагато більш за тих, що зумовлюють рух тулуба.
Центральне перероблення інформації від периферійних рецепторів можна розглядати на 3 функціональних рівнях – аферентної, інтегративної і еферентної підсистем. Аферентний, або сенсорний рівень сомато-вісцеральної системи зв’язаний зі спинним мозком, стовбуром, таламусом та корою великих півкуль головного мозку. Інтегративні функції виконують асоціативна і лімбічна система, які, зокрема, забезпечують спряження сигналів від різних сенсорних систем з інформацією, що зберігається у пам’яті. Еферентні функції виконують рухова і вегетативні системи. Складна взаємодію цих 3 функціональних підсистем лежить у основі поведінки. Така взаємодія у присутності сенсорних стимулів забезпечує сприймання і реакції з 5 компонентами: 1)когнітивним (що включає усвідомлене розрізнення) 2)афективним (що зв’язаний з емоціями) 3)мотиваційним (що зв’язаний з спонуканнями, або драйвами) 4)руховими і 5)вегетативними