

Молекулярні основи мутагенезу
Типи мутацій
Хімічна модифікація
Радіаційне пошкодження
Помилки ДНК-полімерази і систем репарації
Розтягування подвійної спіралі
Біологічний мутагенез
Спрямований мутагенез
1. Типи мутацій
Мутації – наслідувані зміни генетичного матеріалу. Висока стабільність геному забезпечує постійність ознак, які передаються з покоління в покоління. Проте, постійно і з низькою частотою відбуваються мутації, тобто зміна спадкової інформації, закодованої в ДНК. Якщо вони не летальні, то передаються потомству. Мутації є наслідком дії хімічних чинників та опромінення внаслідок зміни структури деяких основ, розриву фосфодиефірних зв’язків чи порушення організації подвійної спіралі. Можливі мутації і при реплікації чи рекомбінації ДНК.
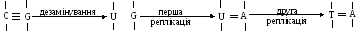
Іншими джерелами мутацій є вставка довших чи коротших фрагментів ДНК в процесі рекомбінації. Інтеркаляція певних циклічних структур органічних речовин також може вести до вставки нуклеотидів. Мутації можуть проявлятися і не проявлятись фенотипово.
Мутації поділяють на 2 типи: заміну основ і зсув рамки. Заміна основи включає транзиції – заміну однієї пари пурин-піримідин іншою, та трансверсії – заміну пари пурин-піримідин парою піримідин-пурин. Зсув рамки зчитування є більш радикальною мутацією ніж заміна основи. В результаті або випадають, або вставляються одна чи багато пар основ.

Точкові мутації
Ці мутації в зоні ДНК, яка кодує білок, майже завжди викликають зміну амінокислотної послідовності відповідної ДНК.

„Помилка” в ДНК транскрибується в РНК і ця змінена РНК слугує матрицею для синтезу білка. Оскільки транскрибується лише одна нитка ДНК, то АТ-пара не рівноцінна ТА-парі. Генетичний код вироджений, тобто в деяких випадках одну амінокислоту кодує більше одного триплету, тому не всі мутації проявляються у зміні структури білка. Рисунок демонструє можливі мутації одного тирозинового кодону. Як видно, зміна РНК з UAС на UAU не призводять до заміни амінокислоти. Такого типу мутації називаються„мовчазними”, тобто кодовані області майже завжди відбуваються в третій основі кодону. Проте аргінін та лейцин можуть мати „мовчазні” мутації також у першій позиції. Заміна першої чи другої основи часто веде до істотних змін білка. Наприклад, одинична заміна UAC на AAC викликає заміну в білку залишку тирозину на аспарагін. Цей тип мутацій отримав назву міссенс-мутації (sense – sequence of amino acids). Кодований поліпептид змінений. Якщо зміна відбувається в критичнім місці поліпептидного ланцюга, то білок може бути або зовсім неактивним, або зі зниженою (збільшеною) активністю. Ще одна можливість – утворення стоп-кодону. В результаті відбудеться передчасна зупинка синтезу білка, який, швидше всього, буде нефункціонуючим. Ці мутації називають нонсенс-мутаціями. Вони змінюють кодон амінокислоти (sense codon) на стоп-кодон (nonsense codon).
Не всі мутації із заміною амінокислот ведуть до утворення нефункціональних білків. Результат залежить від того, в якому місці поліпептиду і яка саме заміна відбулась, як це впливає на вкладання білкової молекули, її каталітичні властивості. Міссенс-мутації можуть призвести до змін, які модифікують чутливість білка до підвищеної температури. Такого типу мутації називають температурно-чутливими. Наприклад, температурно-чутливі мутанти бактерій живуть нормально при 30 0С, але не можуть рости при 400С, хоча „дикий” тип росте при двох температурах. Такі мутації часто називають умовно летальними, оскільки бактерії не можуть рости за одних умов і ростуть за інших. Чутливі до температури фенотипи часто утворюються тому, що мутантний білок може мати коректну укладку структури при низьких температурах, але частково змінену (денатуровану) при високій температурі. Окрім вставки та делеції однієї пари нуклеотидів можливі аналогічні зміни з багатьма парами. Великі делеції чи вставки відбуваються при генетичній рекомбінації. Вставки можуть інактивувати ген, якщо вони розривають його. Багато мутантів в результаті вставки утворюються через перенесення специфічних послідовностей ДНК довжиною 700-1400 пар. Вони називаються вставними послідовностями – типом транспозонних елементів.
Інший тип великих мутацій пов’язаний з реорганізацією в результаті помилок в рекомбінації. Вони включають транслокації та інверсії. При транслокаціях великі фрагменти хромосом переміщуються в нові місця (у еукаріотів інколи на інші хромосоми).
При інверсіях певні фрагменти ДНК змінюють орієнтацію відповідно до оточуючої ДНК. Все це говорить про так звані прямі мутації. Є ще так звані зворотні мутації. Точкові мутації можуть бути оберненими. Найчастіше в такому випадку мова йде про бактерії. Відновлені мутанти мають фенотип диких, який був утрачений у мутантах. Їх іменують ревертантами. Розрізняють два типи ревертантів з реверсіями по тому ж самому місці і по іншому. В першому випадку мутація, що відновлює активність, відбувається в тому ж самому місці, що і вихідна мутація. У другому випадку мутація відбувається в іншому місці ДНК. У випадку реверсії на іншому місці можливе відновлення фенотипу „дикого типу” в результаті кількох типів супресорних мутацій, які відновлюють вихідний фенотип. Супресорні мутації – це нові мутації, які компенсують ефект вихідної мутації. Відомо кілька типів супресорних мутацій:
1. Мутації в іншому місці того ж гену, які відновлюють функцію ферменту, наприклад, мутації зсуву рамки зчитування.
2. Мутації в іншому гені, які можуть відновити фенотип „дикого” типу.
3. Мутації можуть призводити до утворення іншого ферменту, який може замістити мутантний фермент введенням метаболічного шляху, відмінного від того, що використовувався немутантним ферментом.
В останньому випадку вихідний фермент не утворюється, хоча це відбувається в перших двох випадках.
Мутації РНК. РНК-віруси
Швидкість мутуванння РНК-геномів ~1000 разів вища, ніж ДНК. Цьому є кілька причин: 1. РНК-реплікази не мають репаразної активності подібно до ДНК-полімераз. 2. Існує багато ДНК-репаразних механізмів, чого не знайдено для РНК. Це пов’язано, можливо, з тим, що, як правило, ДНК має двоспіральну організацію, а РНК – односпіральну. Інтерес до мутування РНК окрім академічного інтересу, має прикладний, адже багато вірусів є збудниками хвороб, а зміна геному створює додаткові проблеми в їх лікуванні та профілактиці.
2. Хімічна модифікація основ
Основи в ДНК чутливі до дії багатьох хімічних речовин, включаючи азотисту кислоту (НNO2), гідроксиламін (NH2OH) і різні алкілюючі агенти – диметилсульфат і N-метил-N8-нітро-N-нітрозогуанідин.
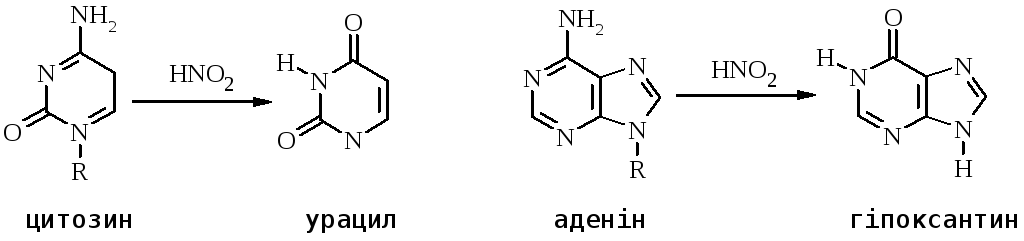
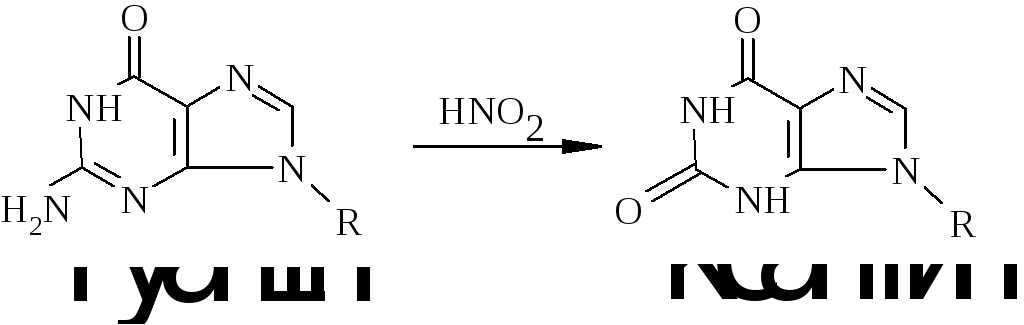
Перетворення гуанідину на ксантин під дією азотистої кислоти не змінює властивостей утворювати водневі зв’язки. Нова основа спарюється з цитозином – нормальним партнером гуаніну. Проте перетворення АHpx або СU змінює нормальне утворення водневих зв’язків у подвійній спіралі.
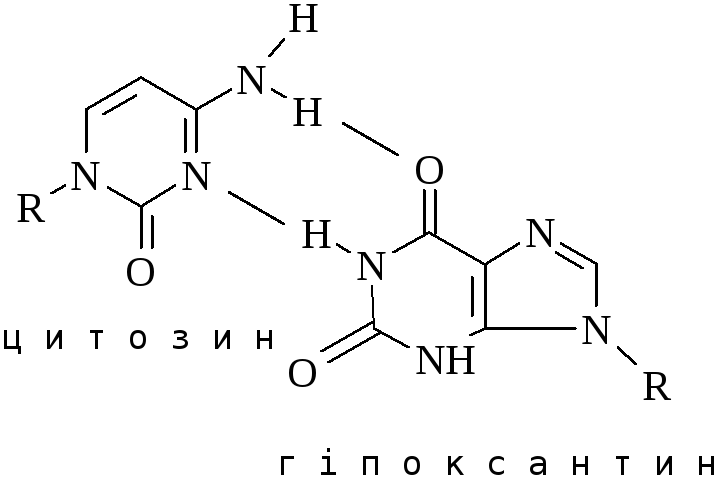
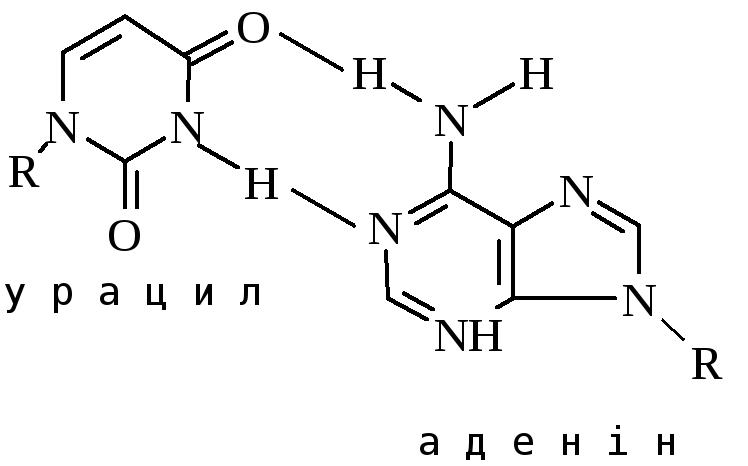
При наступній реплікації відбувається заміна відповідних основ і закріплення заміни.
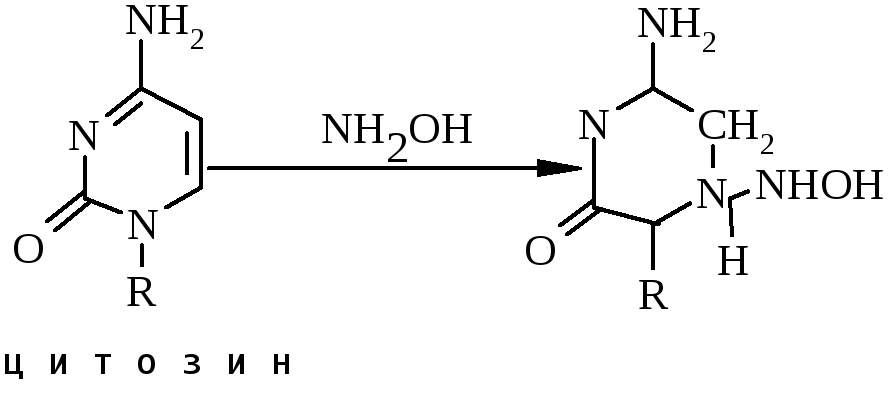
Алкілюючі агенти можуть змінювати структуру азотистих основ, руйнувати фосфодиефірні зв’язки і фрагментувати ланцюги ДНК. Окрім того деякі алкілюючі агенти можуть взаємодіяти ковалентно з двома ланцюгами, утворюючи мостики.
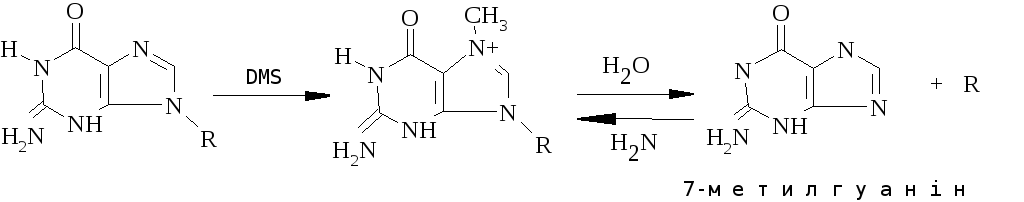
Активовані форми кисню (O2-, H2O2, OH·), які утворюються як побічні продукти нормальних клітинних процесів, також викликають спонтанні зміни ДНК. Спонтанне дезамінування цитозину в ДНК людини відбувається з частотою біля 100 пар основ на геном за день, а депуринізація – в 50 разів частіше.
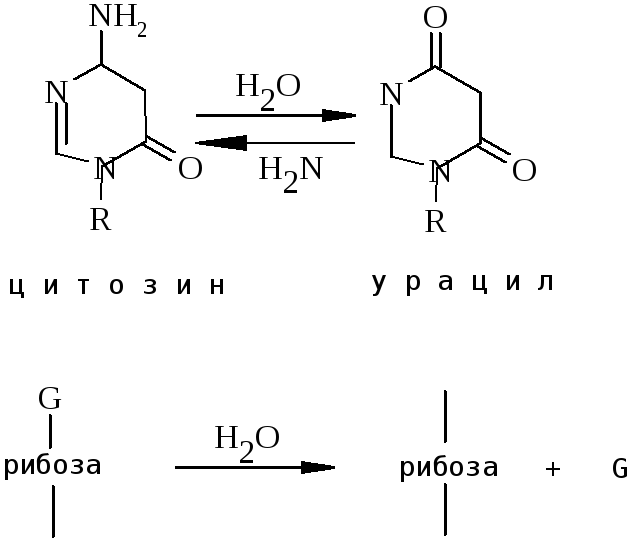
3. Радіаційне пошкодження.
Ультрафіолетове світло, включаючи сонячне, рентгенівські промені поглинаються молекулами ДНК і в залежності від енергії можуть викликати різного типу зміни – від розриву ланцюгів до утворення іонізованих форм азотистих основ. Іонізовані основи не можуть спарюватись з нормальними партнерами. Це істотно збільшує частоту мутацій синтезованої молекули ДНК.
УФ-промені поглинаються азотистими основами, що викликає утворення димерів між розташованими близько піримідиновими основами. Активація етиленових зв’язків цих основ часто призводить до фотохімічної димеризації – утворення Т-Т-зшивок.

Найчастіше таке перетворення відбувається з тимінами. Рідше можливе утворення димерів цитозину, або тиміну з цитозином. Високоенергетична радіація (рентгенівські промені, γ-промені) можуть розривати фосфодиефірні зв’язки, циклічні структури тощо. Радіація в присутності кисню зумовлює утворення великої кількості окислених продуктів.

4. Помилки ДНК-полімерази
З певною частотою ДНК-полімераза приєднує некоректні нуклеотиди. Проте, вони коректуються репаразною активністю ДНК-полімерази. Система корекції невірно вставлених нуклетидів відслідковує порядок по материнській молекулі ДНК, виявляє невірне спарювання. Така корекція здійснюється мультиферментним комплексом, який вирізає невірно вставлені нуклеотиди тільки із нової синтезованої молекули. В молекулі ДНК є спеціальні послідовності GATC,в яких А етилюється, але не зразу, а згодом. Тому комплекс диференціює дві різні молекули ДНК.
ДНК-полімераза нездатна відрізнити нормальні та близькі за структурою дезоксирибонуклеозидтрифосфати. Це також може бути причиною мутацій. Прикладами такого механізму є використання 5-бромурацилу та 2-амінопурину для експериментального мутагенезу. Пара 5-бромурацил-А замінюється на GC-пару з заміною АТ на GC.
Нормально система репараз виправляє пошкодження ДНК. Проте, в деяких випадках репараційна система сама вводить мутації. Часто пошкодження ДНК індукують системи репарації. Описаний складний клітинний механізм, названий SOS-регуляторною системою. Вона активується в результаті пошкодження ДНК, ініціюючи багато процесів репарації ДНК. Проте, в системі SOS деякі процеси репарації відбуваються за відсутності матриці. Це призводить до появи багатьох помилок.
У випадку SOS-системи пошкодження ДНК слугує стресовим сигналом, викликаючи дерегресію (індукцію) чисельних клітинних функцій, включаючи репарацію ДНК. Зазвичай, SOS-система репресована білком LexA.
5. Розтягування подвійної спіралі
Певні хімічні сполуки з ароматичними кільцями і відповідними розмірами та геометрією можуть бути вмонтованими між парами основ у подвійній спіралі ДНК. Цей процес називається інтеркаляцією. При інтеркаляції сусідні пари основ розділюються Це дозволяє стати між ними молекулі інтеркалятора і викликає витягування подвійної спіралі. Такі зміни руйнують неперервність послідовності ДНК і перешкоджають зчитуванню з матриці ДНК-полімеразою. В результаті утворюється дочірня молекула ДНК з додатково вбудованими основами. Такого типу мутації іменуються зсувом рамки. Інтеркаляторами є акридини, атиділбромід тощо. Ці речовини є досить ефективними мутагенами.
6. Біологічний мутагенез.
Мутації можна ввести без використання хімічних чи фізичних чинників. Їх можна викликати так званим транспозонним мутагенезом. Суть його полягає в тому, що якщо всередину гену вставити рухливий генетичний елемент (транспозон), то в результаті розриву гену, останній втрачає функціональну активність. Наприклад, бактеріофаг Muможе слугувати мутагеном, розриваючи в ДНК послідовності, які кодують певні білки. Оскільки рухливі елементи можуть вмонтуватись в будь-якому місці ДНК, то вони широко використовуються для мутагенезу в бактеріях.
7. Спрямований мутагенез (site - directed)
Технологія рекомбінантної ДНК і використання синтетичної ДНК дозволили викликати специфічні мутації специфічних генів. Процедура проведення мутагенезу в специфічних місцях геному отримала назву спрямованого мутагенезу (site-directedmutagenese). Суть методу полягає в наступному. Отримують специфічний ген-ділянку ДНК і визначають його нуклеотидну послідовність. Визначають місця, які кодують початок і кінець синтезу пептиду. На основі цього проводять заміну конкретних основ. Далі змінену ДНК вводять в клітину-реципієнт і бактерії скринінгують за певною ознакою для виявлення мутанту. Ці мутанти будуть відрізнятись від вихідних організмів однією (чи кількома) чітко визначеними наперед замінами. Цілеспрямований мутагенез широко використовується в генетиці мікроорганізмів і молекулярній біології, а також для структурно-функціональних досліджень ферментів та інших білків.