

Вопрос 136
Основные течения
Неоламаркизм включает несколько направлений, или разновидностей: неоламаркизм (в полном смысле слова), связанный с попыткой возродить в модернизированном виде все основные стороны учения Ламарка; ортоламаркизм(от греч. orthos — прямой), абсолютизирующий направленность эволюционного процесса, постулирующий его прямолинейность или даже изначальную запрограммированность;психоламаркизм, рассматривавший в качестве причин эволюции сознательные волевые акты;механоламаркизм, сводивший весь процесс эволюции к прямому или функциональному приспособлению путем наследования приобретаемых признаков;ламаркодарвинизм, придававший наследованию приобретенных признаков ведущую роль в эволюции, а отбору — второстепенную.
Современный взгляд
Арсенал аргументов современных сторонников неоламаркизма весьма широк: от панпсихизма до термодинамики и кибернетики. Так, М. Оже настаивает на сознательном исследовании зародышевой плазмой возможностей изменения своего внутреннего физико-химического равновесия, которые ей предлагаются мутациями. Для Р. Элкена эволюция — целенаправленный, сложный, осмысленный процесс, который не может базироваться на случайных взаимодействиях мутаций и отбора. П. Бриан утверждает, что эволюция не может быть математической игрой, так как статистические закономерности не ведут к творчеству. Ст. Бир в эволюции видит те же системы предетерминированности и планирования, какие характерны для управления концернами. Ссылками на законы термодинамики советский физиолог Н. А. Бернштейн, У. Блум, К. С. Тринчер объясняли запрограммированность эволюции в сторону увеличения упорядоченности, а Г. Гутманн,А. А. Любищеви С. В. Мейен считали, что существуют законы формы, канализирующие эволюцию.
Новейшие открытия подняли вопрос о молекулярно-генетических основах наследования приобретаемых признаков. Так, О. Ландманн в качестве механизмов его реализации называет: наследование измененных структур клеточной оболочки; эпигенетические изменения (см. Эпигенез) без каких-либо модификаций ДНК; наследование ДНК — модификаций, т. е. клонально передаваемых изменений в характере локального метилирования ДНК; индуцированную утрату либо приобретение факультативных элементов генотипа. Высказываются предположения о возникновении адаптивных или отбор-зависимых «направленных» мутациях (Дж. Кэйрнс, Б. Холл, Дж. Шапиро, М. Д. Голубовский). О наследовании приобретенных признаков говорит также, открытый в начале 21 века, горизонтальный перенос генов, посредством ретровирусов и мигрирующих генетических элементов (транспозоны и ретротранспозоны).
Вопрос 137
Популяция — группа особей, способная к более-менее устойчивому самовоспроизводству (как половому, так и бесполому), относительно обособленная (обычно географически) от других групп, с представителями которых (при половой репродукции) потенциально возможенгенетическийобмен. С точки зренияпопуляционной генетики, популяция — это группа особей, в пределах которой вероятность скрещивания во много раз превосходит вероятность скрещивания с представителями других подобных групп. Обычно говорят о популяциях как о группах в составевидаилиподвида.
Изменение генотипа популяции(уравнение харди-вайнберга пример решения задач на эту тему)
Популяционная генетика – это раздел генетики, изучающий генетическое строение и динамику генетического состава популяций – группы организмов, принадлежащих к одному и тому же виду и занимающих ограниченную географическую область обитания. Именно этот раздел науки является основой синтетической теории эволюции, принятой в настоящее время большинством учёных. Генотип совместно с внешней средой определяет фенотип организма. Фенотипы, приспособленные к условиям места обитания, сохраняются в процессе естественного отбора; неприспособленные фенотипы подавляются. Для популяции в целом судьба отдельной особи значения не имеет.
|
|
|
|
Частотой аллеля называют отношение количества данных аллелей у всех особей к общему количеству аллелей в популяции. Частоту доминантного аллеля обычно обозначают буквой p, частоту рецессивного аллеля – буквой q. Если ген представлен двумя аллелями, то выполняется математическое равенство
|
p + q = 1. |
Таким образом, зная частоту одного из аллелей, можно определить частоту и другого аллеля. Так, если частота доминантного аллеля равна 78 %, то частота рецессивного аллеля равняется q = 1 – p = 1 – 0,78 = 0,22 (или 22 %).
Для частот аллелей существует условие равновесия Харди–Вайнберга. Частоты доминантного и рецессивного аллелей остаются неизменными, если в популяции выполняются следующие условия:
размеры популяции достаточно велики;
спаривание и размножение особей происходит случайным образом;
естественный отбор отсутствует (все генотипы одинаково приспособлены к внешним условиям);
различные поколения не скрещиваются между собой;
не возникает новых мутаций;
отсутствует обмен генами с другими популяциями.
Невыполнение одного или нескольких из указанных условий может привести к изменению частоты аллелей и вызвать эволюционные изменения в данной популяции.
Используя символы p и q, можно по-новому записать схему моногибридного скрещивания.
|
|
|
Рисунок 11.3.1.2. Моногибридное скрещивание на языке частот |

|
|
|
Рисунок 11.3.1.3. Зависимость между частотами аллелей и генотипов |
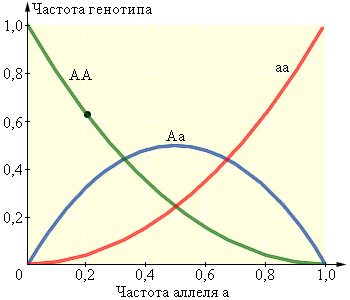
|
p2+ 2pq + q2= 1. |
Эта зависимость называется уравнением Харди–Вайнберга. Используя совместно это уравнение с уравнением p + q = 1, можно вычислить частоту, например, особей, гомозиготных по доминантному аллелю, зная количество носителей рецессивного фенотипа (то есть частоту особей, гомозиготных по рецессивному фенотипу). Пусть q2= 0,0004. Тогда q = 0,02, p = 1 – q = 0,98, p2= 0,9604, 2pq = 0,0392. Следствием уравнения Харди–Вайнберга является значительное превышение (часто на порядки) количества особей, в генотипе которых присутствует рецессивный аллель, над количеством особей с рецессивным фенотипом. Благодаря наличию значительного количества гетерозиготных особей в каждом поколении из генотипа исключается лишь малое количество рецессивных аллелей.
Многие рецессивные гены неблагоприятны для организма и исключаются из генотипа популяции. Другие, наоборот, полезны, благодаря чему частота гетерозиготного генотипа стабилизируется.
Как уже отмечалось, причиной изменений генофонда популяции являются половые рекомбинации генов (кроссинговер, независимое расхождение хромосом, случайное оплодотворение) и мутации. Половые рекомбинации не привносят в популяцию новые аллели; мутации вызывают появление новых аллелей и эволюционные изменения в популяциях. Существуют и другие причины генетической нестабильности, перечисленные ниже.
Небольшая численность популяции может привести к дрейфу генов. Носителей рецессивного аллеля может оказаться настолько мало, что они исчезнут по причинам, не вызванным естественным отбором (например, по причине гибели в результате природного бедствия). Может получиться и обратная ситуация, когда частота рецессивного аллеля случайно вырастет. Это может никак не сказаться на популяции, привести к её вымиранию или наоборот стать благоприятным фактором.
Структурные и поведенческие особенности, благоприятствующие какому-либо признаку (так, брачные игры у животных выигрывает обычно более крупный и сильный самец, а более интенсивно опыляются более яркие и крупные цветки), приводят к неслучайному скрещиванию, то есть к подавлению некоторых аллелей.
Обмен генами между популяциями привносит, с одной стороны, новые гены в популяции, но, с другой стороны, уменьшает общее генетическое разнообразие видов и мешает эволюционным изменениям. Прерывание обмена генами между популяциями является важной предпосылкой для появления нового вида.
Гетерогенность популяций генотипическая(гр. heterogenes — неоднородный по составу) — неоднородность составляющих популяцию особей (клонов), обусловленная разнообразием генотипов в пределах популяционного генофонда.(рассмотрена подробно выше)
Внутрипопуляционный полиморфизм
При внутрипопуляционном полиморфизме внутри единой популяции существуют резко различные, наследственно обусловленные фенотипы (морфы). Различают диморфизм (половой и сезонный), возрастной полиморфизм, фазовость, полиморфизм колоний.
Половой диморфизм- это различие полов по внешним признакам, обусловленное генетическими факторами. Например, самец бабочки крушинницы (лимонницы) имеет лимонно-желтую окраску, а самка - белую. У самцов бабочки голубянки цвет крыльев голубой, а у самок - темно-бурый.
Сезонный диморфизм- различие во внешнем виде организмов одного вида в различные сезоны. Коловратки и дафнии летом и осенью имеют разную форму тела. У бабочки аэшны особи, развивающиеся в разные сезоны, имеют различия в окраске и рисунке крыла.
У общественных насекомых имеется социальный полиморфизм - наличие в семье фенотипически разных групп особей: половых, рабочих, солдат.
При возрастном полиморфизме наблюдаются значительные отличия между личинками и взрослыми. Например, у клешей, насекомых, ракообразных.
Фазовость- фазовая изменчивость, изменение фенотипа особей, определяемая плотностью популяции. Например, у саранчи особи стадной фазы и одиночной фазы отличаются внешне, физиологией и поведением.
Сопряженный полиморфизм- одновременное сохранение нескольких морф в популяции, имеющих преимущество в разных условиях среды. Так, двуточечная божья коровка имеет 2 морфы: красную - хорошо размножающуюся, и черную - хорошо переносящую зимние условия.
Полиморфизм имеет большое биологическое значение, так как он обеспечивает существование вида в разнообразных условиях, дает материал дивергенции - расхождения признаков и групп особей, позволяет популяции лучше использовать многообразные условия среды.