

Лекция биология VI
.docx

Кодоминирование
При кодоминировании, в отличие от неполного доминирования, у гетерозигот признаки, за которые отвечает каждый из аллелей, проявляются одновременно (смешанно). Типичный пример кодоминирования — наследование групп крови системы АВ0 у человека. Всё потомство людей с генотипами АА (вторая группа) и ВВ (третья группа) будет иметь генотип АВ (четвертая группа). Их фенотип не является промежуточным между фенотипами родителей, так как на поверхности эритроцитов присутствуют оба агглютиногена (А и В). При кодоминировании назвать один из аллелей доминантным, а другой — рецессивным нельзя, эти понятия теряют смысл: оба аллеля в равной степени влияют на фенотип. На уровне РНК и белковых продуктов генов, видимо, подавляющее большинство случаев аллельных взаимодействий генов — это кодоминирование, ведь каждый из двух аллелей у гетерозигот обычно кодирует РНК и/или белковый продукт, и оба белка или РНК присутствуют в организме.
Экспрессивность и пенетрантность
У некоторых гетерозиготных организмов при полном доминировании наблюдается различная степень выраженности доминантного признака. В этом случае говорят о различной экспрессивности и пенетрантности генов.
Экспрессивность – степень проявления доминантного признака у гетерозигот.
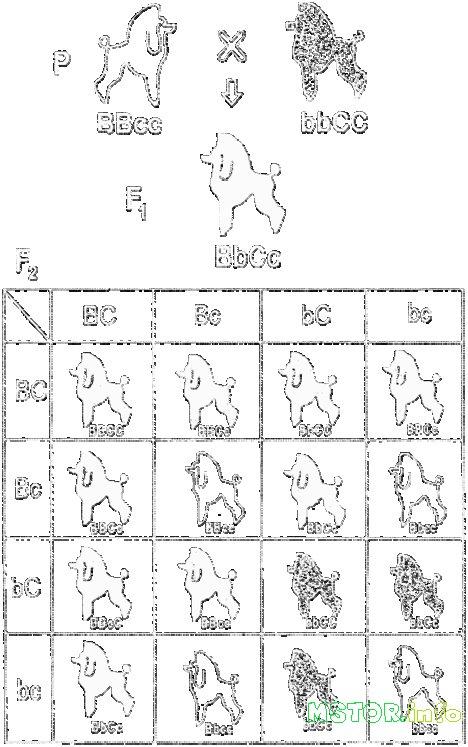
Неалле́льные ге́ны — это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой.
При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы и взаимодействия неаллельных генов:
-
комплементарность;
-
эпистаз;
-
полимерия.
-
Пенетрантность (генетика популяций) — показатель фенотипического проявления аллеля в популяции. Определяется как отношение (обычно — в процентах) числа особей, у которых наблюдаются фенотипические проявления наличия аллеля, к общему числу особей, у которых данный аллель присутствует в необходимом для фенотипического проявления количестве копий (в зависимости от характера доминирования, для фенотипического проявления может быть достаточно только одной копии аллеля или двух, если для фенотипического проявления необходимо, чтобы особь былагомозиготна по данному гену).
-
Например, фраза «аллель A обладает пенетрантностью 95 %» означает, что из всех особей, у которых данный аллель имеется в необходимом числе копий, лишь у 95 % наличие этого аллеля можно установить по показателям фенотипа. Полная пенетрантность — это 100 % фенотипическое проявление наличия данного аллеля в пределах популяции.
-
Проще говоря, это частота проявления гена в признак.
Комплементарность
Материал из Википедии — свободной энциклопедии
Сюда перенаправляется запрос «комплиментарность». На эту тему нужна отдельная статья.
Комплемента́рность:
-
Комплементарность в химии, молекулярной биологии и генетике — взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими (комплементарными) фрагментами молекул или их структурных фрагментов вследствие супрамолекулярных взаимодействий.
-
В философии комплементарными называют несходные или даже противоположные теории, концепции, модели и точки зрения, отражающие различные взгляды на действительность.
-
Комплементарность в генетике — форма взаимодействия неаллельных генов, при котором одновременное действие нескольких доминантных генов дает новый признак.
-
Комплементарные блага (комплементы) — это несколько товаров (два и более), которые дополняют друг друга и потребляются одновременно. Примерами таких товаров могут служить: автомобильи бензин; компьютер, монитор, клавиатура и мышка; подушка и одеяло.
-
Комплементарность в электронике — точное взаимосоответствие параметров при различных условиях. Пример: Комплементарная пара транзисторов, Комплементарная МОП логика.
-
Эписта́з — взаимодействие генов, при котором активность одного гена находится под влиянием другого гена (генов), неаллельного ему. Ген, подавляющий фенотипические проявления другого, называется эпистатичным; ген, чья активность изменена или подавлена, называется гипостатичным.
-
Примеры эпистатического влияния тесно связанных генов на приспособленность можно обнаружить в супергенах и главном комплексе гистосовместимости. Эффект может проявляться как напрямую — на уровне генов (при этом продукт эпистатичного гена предотвращает транскрипцию гипостатичного), так и на уровне фенотипов.

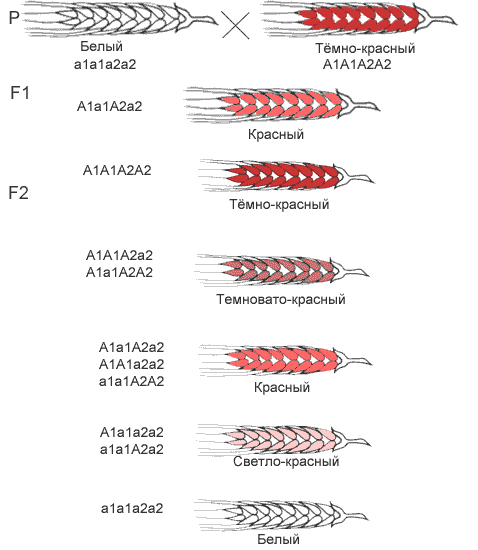
Полимерия – одновременное действие на признак нескольких генов.
Гены, лежащие в одной и той же хромосоме, называют сцепленными. Все гены какой-либо одной хромосомы образуют группу сцепления; они обычно попадают в одну гамету и наследуются вместе. Таким образом, гены, принадлежащие к одной группе сцепления, обычно не подчиняются менделевскому принципу независимого распределения. Поэтому при дигибридном скрещивании они не дают ожидаемого отношения 9:3:3:1. В таких случаях получаются самые разнообразные соотношения. У дрозофилы гены, контролирующие окраску тела и длину крыла, представлены следующими парами аллелей (назовем соответствующие признаки): серое тело - черное тело, длинные крылья - зачаточные (короткие) крылья. Серое тело и длинные крылья доминируют. Ожидаемое отношение фенотипов в F2 от скрещивания между гомозиготой с серым телом и длинными крыльями и гомозиготой с черным телом и зачаточными крыльями должно составить 9: 3: 3: 1. Это указывало бы на обычное менделевское наследование при дигибридном скрещивании, обусловленное случайным распределением генов, находящихся в разных, негомологичных хромосомах. Однако вместо этого в F2 были получены в основном родительские фенотипы в отношении примерно 3: 1. Это можно объяснить, предположив, что гены окраски тела и длины крыла локализованы в одной и той же хромосоме, т.е. сцеплены.
Практически, однако, соотношение 3:1 никогда не наблюдается, а возникают все четыре фенотипа. Это объясняется тем, что колкое сцепление встречается редко. В большинстве экспериментов по скрещиванию при наличии сцепления помимо мух с родительскими фенотипами обнаруживаются особи с новыми сочетаниями признаков. Эти новые фенотипы называют рекомбинантными. Все это позволяет дать следующее определение сцепления: два или более генов называют сцепленными, если потомки с новыми генными комбинациями (рекомбинанты) встречаются реже, чем родительские фенотипы.
В клетках человека имеются 23 пары хромосом. 22 пары одинаковы у женщин и мужчин. 23-я пара у женщин содержит две одинаковые хромосомы (их называют Х-хромосомами), а у мужчин - две разные хромосомы (Х-хромосому и Y-хромосому). Эти хромосомы, по которым отличаются между собой мужчины и женщины, называются половыми. В половые клетки попадает только хромосома из каждой пары. Поэтому мужские половые клетки у человека несут в себе или Х-хромосому, или Y-хромосому. От того, с какой из этих клеток сольётся при оплодотворении женская половая клетка (всегда несущая Х-хромосому), будет зависеть пол зародыша. Женщина получает две Х-хромосомы: одну от отца и одну от матери. Мужчина получает одну Х-хромосому от матери и одну Y-хромосому от отца. Некоторые признаки определяются генами, которые находятся в половых хромосомах. Такие признаки называются признаками, сцепления с полом. Например, у человека в Х-хромосомах встречается рецессивный ген, вызывающий тяжёлую болезнь - не свёртываемость крови (гемофилию). Этот рецессивный ген обычно проявляется только у мужчин.

Рис. 5. Вид половых хромосом человека в метафазе митоза.
Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х - хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом, например цветовой слепоты, раннего облысения и гемофилии у человека. Гемофилия - сцепленный с полом рецессивный признак, при котором нарушается образование фактора VIII, ускоряющего свертывание крови. Ген, детерминирующий синтез фактора VIII, находится в участке Х - хромосомы, не имеющем гомолога, и представлен двумя аллелями - доминантным нормальным и рецессивным мутантным.
Хромосомная теория наследственности
Хромосомная теория наследственности
I Хромосо́мная тео́рия насле́дственности (греч. chrōma цвет, окраска + sōma тело)
основная теория современной генетики, согласно которой главными материальными носителями наследственности являются хромосомы и расположенные на них в определенной линейной последовательности гены
II Хромосо́мная тео́рия насле́дственности
теория наследственности, согласно которой передача признаков от родителей потомству происходит через хромосомы; экспериментально подтверждена Т. Морганом.
Генетические карты хромосом — это схема взаимного расположения и относительных расстояний между генами определенных хромосом, находящихся в одной группе сцепления.
Впервые в 1913 — 1915 годах на возможность построения генетических карт хромосом указывают Т. Морган и его сотрудники. Они экспериментально показали, что основываясь на явлениях сцепления генов и кроссинговера можно построить генетические карты хромосом [1]. Возможность картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др.), грибов (дрожжи, аспергилл), для бактерий и вирусов.
Генетические карты человека используются в медицине при диагностике ряда тяжелых наследственных заболеваний человека. В исследованияхэволюционного процесса сравнивают генетические карты разных видов живых организмов. Помимо генетических, существуют и другие карты хромосом.
Физическая карта – графическое представление порядка следования физических маркеров (фрагментов молекулы ДНК), расстояние между которыми определяется в парах нуклеотидов.
Томас Хант Морган (англ. Thomas Hunt Morgan; 25 сентября 1866, Лексингтон — 4 декабря 1945, Пасадина) — американский биолог, один из основоположников генетики, председатель Шестого Международного конгресса по генетике в Итаке, Нью-Йорк (1932). Лауреат Нобелевской премии по физиологии и медицине 1933 года «За открытия, связанные с ролью хромосом в наследственности».
Томас Морган и его ученики (Г. Дж. Меллер, А. Г. Стертевант и др.) обосновали хромосомную теорию наследственности; установленные закономерности расположения генов в хромосомах способствовали выяснению цитологических механизмов законов Грегора Менделя и разработке генетических основ теории естественного отбора. Умер 4 декабря 1945 года от желудочного кровотечения.
ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ - схемы относительного расположения генов в хромосомах, позволяющие предсказывать характернаследования изучаемых признаков организмов.