

Суммарное уравнение анаэробного гликолиза.
Глюкоза + 2 АДФ + 2 H3PO4 = 2 Лактат + 2 АТФ + 2 Н2О
В анаэробном гликолизе АТФ образуется только за счет субстратного фосфорилирования (2 моль АТФ на 1моль глюкозы).
Значение анаэробного гликолиза особенно велико для скелетных мышц. В интенсивно работающих скелетных мышцах мощность механизма транспорта кислорода оказывается недостаточной для обеспечения энергетической потребности, в этих условиях резко усиливается анаэробный гликолиз, и в мышцах накапливается молочная кислота. Особое значение анаэробный гликолиз имеет при кратковременной интенсивной работе. Так, бег в течение 30 с полностью обеспечивается анаэробным гликолизом. Однако уже через 4-5 мин бега энергия поставляется поровну анаэробным и аэробным процессами, а через 30 мин - почти целиком аэробным процессом. При длительной работе в аэробном процессе используется не глюкоза, а жирные кислоты.
Эритроциты вообще не имеют митохондрий, поэтому их потребность в АТФ удовлетворяется только за счет анаэробного гликолиза. Интенсивный гликолиз характерен также для злокачественных опухолей.
Цикл Кори (глюкозо-лактатный цикл) и глюкозо-аланиновый цикл.
Молочная кислота не является конечным продуктом обмена, но ее образование - тупиковый путь метаболизма: ее дальнейшее использование связано опять с превращением в пируват. Из клеток, в которых происходит гликолиз, образующаяся молочная кислота поступает в кровь и улавливается в основном печенью, где и превращается в пируват. Пируват в печени частично окисляется, а частично превращается в глюкозу. Часть пирувата в мышцах путем трансаминирования превращается в аланин, который транспортируется в печень, и здесь снова образует пируват.
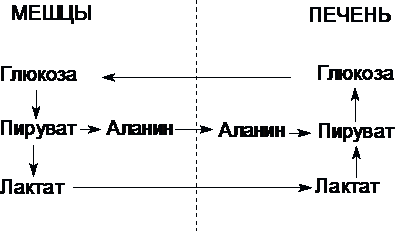
Рис. Цикл Кори и глюкозо-аланиновый цикл
Биосинтез глюкозы (глюконеогенез).
Глюконеогенез протекает в основном по тому же пути, что и гликолиз, но в обратном направлении. Однако три реакции гликолиза необратимы, и на этих стадиях реакции глюконеогенеза отличаются от реакций гликолиза.
Обходные пути глюконеогенеза.
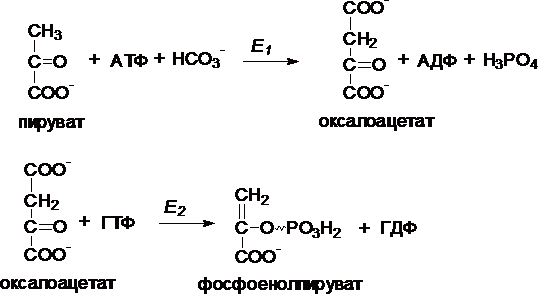
I. Первый обходной путь - превращение пирувата в фосфоенолпируват.
Это превращение осуществляется под действием двух ферментов - пируваткарбоксилазы (Е1) и карбокикиназы фосфоенолпирувата (Е2).
II. Превращение фруктозо-1,6-дифосфата во фруктозо-6-фосфат катализирует фермент фруктозо-1,6-дифосфатаза.

III. Образование глюкозы из глюкозо-6-фосфата катализирует фермент глюкозо-6-фосфотаза .
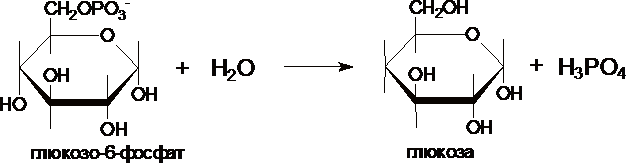
Этот фермент отсутствует в клетках мозга, скелетных мышц и др. тканях, поэтому глюконеогенез, протекающий в этих тканях не может поставлять глюкозу в кровь. Глюкозо-6-фосфатаза локализована в почках и, особенно, в печени, и глюкозо-6-фосфат может превращаться в глюкозу, если ее уровень в крови снижен.
Пентозофосфатный путь (пфп). Значение пфп.
Большая часть глюкозы расщепляется по гликолитическому пути. Наряду с гликолизом существуют второстепенные пути катаболизма глюкозы, одним из которых является пентозофосфатный путь (ПФП). Пентозофосфатный путь скорее даже - анаболический путь, который использует 6 атомов С глюкозы для синтеза пентоз и восстановительных эквивалентов в виде НАДФН, необходимых для образования липидов в организме. Однако, этот путь - окисление глюкозы, и при определенных условиях может завершиться полным окислением глюкозы до СО2 и Н2О. Главные функции этого цикла следующие:
Генерирование восстановительных эквивалентов в виде НАДФН для восстановительных реакций биосинтеза. Ферменты, которые катализируют реакции восстановления, часто используют в качестве кофактора НАДФ+/НАДФН, а окислительные ферменты - НАД+/НАДН. Восстановительные реакции биосинтеза жирных кислот и стероидных гормонов требуют НАДФН, поэтому клетки печени, жировой ткани и др., где протекает синтез липидов, имеют высокий уровень ферментов ПФП. В печени около 30% глюкозы подвергаются превращениям в ПФП.
НАДФН используется при обезвреживании лекарств и чужеродных веществ в монооксигеназной цепи окисления в печени.
НАДФН предохраняет ненасыщенные жирные кислоты эритроцитарной мембраны от аномальных взаимодействий с активным кислородом. Кроме того, эритроциты используют реакции ПФП для генерирования больших количеств НАДФН, который необходим для восстановления глутатиона, участвующего в системе обезвреживания активного кислорода, что позволяет поддерживать нормальную степень окисления Fe2+ в гемоглобине.
Снабжение клеток рибозо-5-фосфатом для синтеза нуклеотидов, нуклеотидных коферментов и нуклеиновых кислот. Превращение рибонуклеотидов в дезоксирибонуклеотиды также требует НАДФН, поэтому многие быстро пролиферирующие клетки требуют больших количеств НАДФН.
Хотя это не значительная функция ПФП, он принимает участие в метаболизме пищевых пентоз, входящих в состав нуклеиновых кислот пищи и может превращать их углеродный скелет в интермедиаты гликолиза и глюконеогенеза.