

Биосинтез ДНК. Этапы биосинтеза ДНК: инициация, элонгация и терминация синтеза. Размеры молекул ДНК. Представление об укладке ДНК в хроматине и хромосомах. Репликация ДНК и фазы клеточного цикла. Репарация повреждений и ошибок репликации ДНК.
Молекулярная масса ДНК варьирует от 1 000 до 1 000 000 кД. Общая длина всех молекул ДНК гаплоидного набора составляет 150 млн п.н. Репликация такой большой молекулы со скоростью 50 нуклеотидов в минуту шла бы примерно 800 ч.
Компактизация и суперспирализация ДНК осуществляются с помощью разнообразных белков, взаимодействующих с определёнными последовательностями в структуре ДНК. Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы: гистоновые и негистоновые белки. Комплекс белков с ядерной ДНК клеток называют хроматином.

Гистоны - белки с молекулярной массой 11-21 кД, содержащие много остатков аргинина и лизина. Благодаря положительному заряду гистоны образуют ионные связи с отрицательно заряженными фосфатными группами, расположенными на внешней стороне двойной спирали ДНК.
Существует 5
типов гистонов. Четыре гистона Н2А, Н2В,
НЗ и Н4 образуют октамерный белковый
комплекс (Н2А, Н2В, НЗ, Н4)2,
который называют "нуклеосомный
кор" (от
англ. nucleosome
core).
Молекула ДНК "накручивается" на
поверхность гистонового октамера,
совершая 1,75 оборота (около 146 пар
нуклеотидов). Такой комплекс гистоновых
белков с ДНК служит основной структурной
единицей хроматина, её называют
"нуклеосома".
ДНК,
связывающую нуклеосомные частицы,
называют линкерной ДНК. В среднем
линкерная ДНК составляет 60 пар нуклеотидных
остатков. Молекулы гистона H1 связываются
с ДНК в межнуклеосомных участках
(линкерных последовательностях) и
защищают эти участки от действия нуклеаз
(рис. 4-8).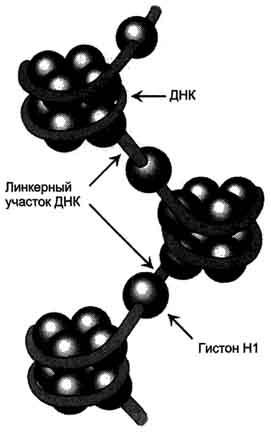
Рис. 4-8. Структура нуклеосом. Восемь молекул гистонов (Н2А, Н2В, НЗ, Н4)2 составляют ядро нуклеосомы, вокруг которого ДНК образует примерно 1,75 витка.
Негистоновые белки хроматина
В ядре эукариотической клетки присутствуют сотни самых разнообразных ДНК-связывающих негистоновых белков. Каждый белок комплементарен определённой последовательности нуклео-тидов ДНК (сайт ДНК). К этой группе относят семейство сайт-специфических белков типа "цинковые пальцы". Каждый "цинковый палец" узнаёт определённый сайт, состоящий из 5 нуклеотидных пар. Другое семейство сайт-специфических белков - гомодимеры. Фрагмент такого белка, контактирующий с ДНК, имеет структуру "спираль-поворот-спираль". К группе структурных и регуляторных белков, которые постоянно ассоциированы с хроматином, относят белки высокой подвижности (HMG-белки - от англ, high mobility gel proteins). Они имеют молекулярную массу менее 30 кД и характеризуются высоким содержанием заряженных аминокислот. К негистоновым белкам принадлежат также ферменты репликации, транскрипции и репарации. При участии структурных, регуляторных белков и ферментов, участвующих в синтезе ДНК и РНК, нить нуклео-сом преобразуется в высококонденсированный комплекс белков и ДНК. Образованная структура в 10 000 раз короче исходной молекулы ДНК.
Различают гетерохроматин и эухроматин.
• Гетерохроматин - транскрипционно неактивный и конденсированный хроматин интерфазного ядра, составляет 10% от общего хроматина. Пример-тельце Барра (инактивированная одна из Х-хромосом)
• Эухроматин - составляет 90% от общего хроматина, из них 10% - транскрипционно активная часть.
Хромосомы (рис. 2-17) образуются, когда полностью конденсированный хроматин образует многочисленные плотно упакованные петли. Каждая хромосома содержит одну длинную двуцепочечную молекулу ДНК и ДНК-связывающие белки. Результатом взаимодействия ДНК с ДНК-связывающими белками является компактизация хроматина. Длина молекулы ДНК в составе одной хромосомы приблизительно составляет 4 см, тогда как длина метафазных хромосом равняется 4 мкм.
Репликация ДНК.
Процесс удвоения хромосом называют репликацией (редупликацией). Она происходит в течение S-фазы клеточного цикла.
Хромосома
содержит одну непрерывную двухцепочечную
молекулу ДНК. При репликации каждая
цепь родительской двухцепочечной ДНК
служит матрицей для синтеза новой
комплементарной цепи. Вновь образованная
двойная спираль имеет одну исходную
(родительскую) и одну вновь синтезированную
(дочернюю) цепь. Такой механизм удвоения
ДНК получил название "полуконсервативная
репликация" (рис.
4-14).
Рис. 4-14. Полуконсервативная репликация.
Репликацию можно разделить на 3 этапа: образование репликативной вилки (инициация), синтез новых цепей (элонгация), завершение синтеза двух дочерних цепей ДНК (терминация).
Ориджины репликации
Инициация синтеза ДНК происходит в нескольких сайтах хромосомы, которые называют сайтами инициации репликации, или ориджинами (от англ. origin - происхождение) репликации (рис. 4-17). Ориджины репликации имеют определённую нуклеотидную последовательность. Последовательность ДНК, ограниченную двумя ориджинами репликации, называют единицей репликации, или репликоном.
А. Инициация репликации
Инициацию репликации регулируют специфические сигнальные белковые молекулы - факторы роста. Они связываются рецепторами мембран клеток, которые передают сигнал, побуждающий клетку к началу репликации.
Синтез новых одноцепочечных молекул ДНК может произойти только при расхождении родительских цепей. В определённом сайте (точка начала репликации) происходит локальная денатурация ДНК, цепи расходятся и образуются две репликативные вилки, движущиеся в противоположных направлениях.
В образовании репликативной вилки принимает участие ряд белков и ферментов. Cемейство ДНК-топоизомераз (I, II и III) участвует в регуляции суперспирализации ДНК. Например, ДНК-топоизомераза I разрывает фосфоэфирную связь в одной из цепей двойной спирали и ковалентно присоединяется к 5'-концу в точке разрыва (рис. 4-15). По окончании формирования репликативной вилки фермент ликвидирует разрыв в цепи и отделяется от ДНК.
ДНК-хеликаза осуществляет разрыв водородных связей в двухцепочечной молекуле ДНК. Фермент ДНК-хеликаза использует энергию АТФ для расплетения двойной спирали ДНК.
В результате происходит раскручивание участка суперспирализованной молекулы ДНК.
В поддержании этого участка ДНК в раскрученном состоянии участвуют SSB-белки (от англ, single strand binding proteins, т.е. белки, связывающиеся с одноцепочечными нитями ДНК). SSB-белки связываются с одноцепочечной ДНК по всей длине разделившихся цепей и таким образом предотвращают их комплементарное скручивание и образование "шпилек".

Рис. 4-15. Участие ДНК-топоизомеразы I в образовании репликативной вилки. 1 - фермент расщепляет одну цепь ДНК; между остатком тирозина молекулы фермента и фосфорным остатком цепи образуется ковапентная связь; 2 - происходит локальное раскручивание двойной спирали при участии ДНК-хеликазы; ДНК-топоизомераза I восстанавливает фосфоэфирную связь.