

3. Механизм действия гормонов
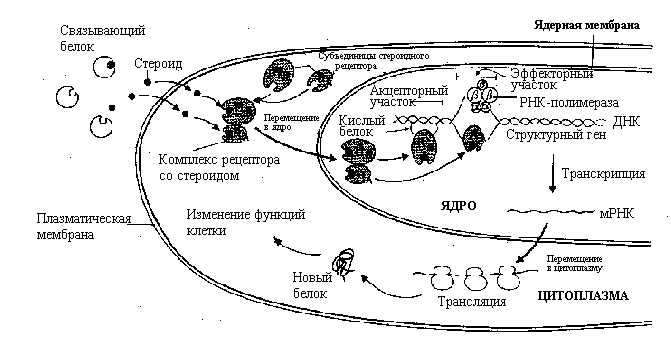
Гормоны стероидной природы реализуют свои эффекты, проникая в клетку. Плазматическая мембрана не является для них непреодолимой преградой ввиду их липофильности и небольшой молекулярной массы. Поступив в клетку, стероиды образуют комплекс со специфическим цитозольным рецептором, который после предварительной трансформации поступает в ядро, где связывается со специфическими регуляторными элементами ДНК – т.е. с определенными нуклеотидными последовательностями, чувствительными к гормонам (в ряде случаев гормон-рецепторные комплексы действуют как энхансеры или сайленсеры). Результатом этого взаимодействия является синтез всех видов РНК и синтез на рибосомах новых количеств белка.
В отличие от стероидных гормоны белково-пептидной природы не способны проникать через клеточную мембрану и потому реализуют свои эффекты через трансдукционные системы, т.е. специальные системы, передающие сигнал снаружи внутрь клетки. Можно выделить 4 основных класса трансдукционных систем:
1) Системы, передающие сигнал при участии G-белков. Последние представляют собой семейство низкомолекулярных мембранных белков, связывающих гуаниловые нуклеотиды. Образовавшийся на поверхности клеточной мембраны гормон-рецепторный комплекс активируетG-белок. Механизм активации заключается в замещении ГДФ, содержащейся в неактивномG-белке, на ГТФ и последующей диссоциации-субъединицы от ингибиторных-субъединиц. После передачи сигналаG-белок возвращается в исходное неактивное состояние вследствие действия ГТФазы, имеющейся уG-белка и гидролизующей ГТФ до ГДФ. Описаны две системы, функционирующие с участиемG-белков:
а) трансдукционная система, в которой эффекторным ферментом является аденилатциклаза, катализирующая образование цАМФ. Примерами могут служить системы, включаемые Н2-гистаминовым рецептором,V2-рецептором вазопрессина, рецептором глюкагона,1- и2-адренорецепторами и т.д.
б) трансдукционная система, в которой эффекторным ферментом является фосфолипаза С. Последняя гидролизует мембранный фосфатидилинозитолдифосфат (ФИФ2) с образованием вторичных посредников: инозитолтрифосфата (ИФ3) и диацилглицерола (ДАГ). Примерами могут служить системы, включаемыеV1-рецептором вазопрессина,1-адренорецептором, рецептором ангиотензина и т.д.
2) Система, в которой эффекторным ферментом является гуанилатциклаза, катализирующая образование цГМФ. Примером может служить система, включаемая рецептором предсердного натрий-уретического пептида.
3) Системы, в которых эффекторным ферментом является тирозинкиназа – фермент, катализирующий фосфорилирование белков-субстратов по остаткам тирозина. Примерами могут служить системы, включаемые рецептором инсулина и различных ростовых факторов: фактора роста нервов, тромбоцитов, эпидермиса и т.д.
4) Системы, в которых рецептор одновременно является ионным каналом. Примером может служить рецептор ГАМК, являющийся одновременно каналом для CI-, а также никотиновый ацетилхолиновый рецептор одновременно являющийсяNa+-каналом.
Таким образом, белково-пептидные гормоны внутрь клетки не проникают, но в результате активации соответствующих трансдукционных систем внутри клетки образуются (или поступают извне) химические посредники, которые реализуют функции гормона внутри клетки. Эти посредники получили название вторичных мессенджеров (посредников). При этом в роли первого сигнала рассматривается сам гормон. В настоящее время можно выделить не менее 4 типов вторичных гормональных посредников: 1) циклические нуклеотиды (цАМФ и цГМФ), 2) ионы Са, 3) метаболиты фосфатидилинозитола и 4) циклическая АДФ-рибоза.