

Вопрос 33
Амитозом называется деление клетки, находящейся в состоянии интерфазы. К амитозу иногда относят все случаи немитотического деления клетки (рис. 66). При этом не происходит конденсации хромосом, распада ядерной оболочки и образования веретена деления; амитоз осуществляется при вытягивании ядра и его последующем делении на две части. Еще более неупорядоченное дробление ядра на два или более неидентичных комка получило название фрагментации; оно, безусловно, носит патологический характер. Однако между амитозом и фрагментацией резкой и принципиальной границы провести нельзя.
Прямое деление клетки впервые было описано Р. Ремаком в 1841 г. у животных и Э. Страсбергером в 1882 г. у растений. Вначале амитоз рассматривали как более примитивную форму деления ядра в отличие от митоза. Современными исследованиями это представление полностью опровергнуто на основании данных по сравнительной цитологии и эмбриологии, показавших, что митоз, встречающийся даже у простейших организмов, является первичной формой размножения. Амитоз по сравнению с ним наблюдается редко и возникает преимущественно в клетках высокодифференцированных тканей или дегенерирующих, не способных к дальнейшему воспроизведению. Так, напримёр, у растений амитоз обнаруживается в клетках отмирающих или временных тканей стенок завязи, нуцеллуса, эндосперма, в паренхиме клубней и т. д.
А — начальная фаза: / — появление в ядре второго маленького ядрышка, 2 — повышение ба-зофилии ядрышка; £ —средняя и заключительная фазы амитотнческого деления ядра: 1, 2 — появление на поверхности ядра вздутия, 3, 4— отделение одного ядра от другого. По Прокофьевой-Бельговской.[ ...]
Различия в состоянии ядер, возникших путем амитоза, по-видимому, обусловлены тем, что одно из них старое, уже функционировавшее, а другое молодое, заново в нем развившееся. Такое-различие в свойствах ядер А. А. Прокофьева-Бельгов-ская объясняет изменением свойств структурных белков в период крахмалообразования. По данным радиоавтографических исследований, прямое деление клетки может осуществляться как в период синтеза ДНК, так и в промитотический (постсин-тетический) период клеточного цикла. Однако увеличение количества ДНК при амитозе обнаруживается не во всех делящихся ядрах и, кроме того, неравномерно в отличие от митоза, при котором всегда происходит кратное увеличение ДНК, что очень важно для оценки функционального значения митоза и амитоза.
58 . Классификация генов Накопленные знания о структуре, функциях, характере взаимодействия, экспрессии, мутабильности и других свойствах генов породили несколько вариантов классификации генов.
Накопленные знания о структуре, функциях, характере взаимодействия, экспрессии, мутабильности и других свойствах генов породили несколько вариантов классификации генов.
По месту локализации генов в структурах клетки различают расположенные в хромосомах ядра ядерные гены и цитоплазматические гены, локализация которых связана с хлоропластами и митохондриями.
По функциональному значению различают структурные гены, характеризующиеся уникальными последовательностями нуклеотидов, кодирующих свои белковые продукты, которые можно идентифицировать с помощью мутаций, нарушающих функцию белка, и регуляторные гены — последовательности нуклеотидов, не кодирующие специфические белки, а осуществляющие регуляцию действия гена (ингибирование, повышение активности и др.).
По влиянию на физиологические процессы в клетке различают летальные, условно летальные, супервитальные гены, гены-мутаторы, гены-антимутаторы и др.
Следует отметить, что любые биохимические и биологические процессы в организме находятся под генным контролем. Так, деление клеток (митоз, мейоз) контролируется несколькими десятками генов; группы генов осуществляют контроль восстановления генетических повреждений ДНК (репарация). Онкогены и гены — супрессоры опухолей участвуют в процессах нормального деления клеток. Индивидуальное развитие организма (онтогенез) контролируется многими сотнями генов. Мутации в генах приводят к измененному синтезу белковых продуктов и нарушению биохимических или физиологических процессов.
Гомеозисные мутации у дрозофилы позволили открыть существование генов, нормальной функцией которых является выбор или поддержание определенного пути эмбрионального развития, по которому следуют клетки. Каждый путь развития характеризуется экспрессией определенного набора генов, действие которых приводит к появлению конечного результата: глаза, голова грудь, брюшко, крыло, ноги и т. д. Исследования генов комплекса bithorax дрозофилы американским генетиком Льюисом показали, что это гигантский кластер тесно сцепленных генов, функция которых необходима для нормальной сегментации груди (thorax) и брюшка (abdomen). Подобные гены получили название гомеобоксных. Гомеобоксные гены расположены в ДНК группами и проявляют свое действие строго последовательно. Такие гены обнаружены и у млекопитающих, и они имеют высокую гомологию (сходство).
В зависимости от локализации генов в структурах клетки различают ядерные и митохондриальные гены (рис. IV. 14). По своему функциональному назначению гены могут быть разделены на две группы. Первая группа представлена генами, кодирующими белки; вторая группа — генами, контролирующими синтез РНК. Среди генов, кодирующих белки, различают: — гены «домашнего хозяйства», продукты которых необходимы для обеспечения функции любого типа клеток; — гены терминальной дифференцировки, т. е. гены, обеспечивающие специализированные функции клеток; — гены траскрипционных факторов, контролирующие особые ядерные белки, способные соединяться с регуляторными областями многих структурных генов, вызывая либо активацию, либо подавление транскрипции. РНК-кодирующие гены определяют синтез различных видов РНК, необходимых для синтеза рибосом, обеспечения процессов трансляции, сплайсинга, а также для синтеза молекул РНК, влияющих на функционирование других генов (регуляторное действие). Гены человека, как правило, представляют собой функционально прерывистую последовательность нуклеотидов. Относительно короткие кодирующие последовательности оснований чередуются в них с длинными некодирующими последовательностями. Последовательности гена, представленные в молекуле зрелой иРНК, получили название экзонов. Именно экзоны являются кодирующими участками гена, контролирующими аминокислотную последовательность белков. Экзоны разделены некодирующими участками — нитронами, которые вырезаются (сплайсинг) в процессе созревания иРНК и не участвуют в процессе трансляции. В настоящее время в понятие «ген» включаются не только транскрибируемые области (экзоны и интроны), но и фланкирующие ген последовательности. Фланкирующие области гена, как правило, высоко консервативны, т. е. характеризуются постоянством нук-леотидной последовательности, наблюдаемым даже при сравнении представителей различных видов. Фланкирующие области гена содержат последовательности, необходимые для его правильной работы: например, промоторная область в начале 5'-области или хвостовая нетранслируемая область поли-А, расположенная на З'-конце гена. Так, ТАГА — бокс (последовательность чередования тимина и аденина) обеспечивает правильную ориентацию РНК-полимеразы, что необходимо для транскрипции РНК. Размеры генов человека могут колебаться от нескольких десятков пар нуклеотидов (п.н.) до многих тысяч и даже миллионов п.н. Так, самый маленький из известных генов содержит всего 21 п.н., а один из самых крупных генов — ген дистрофина — имеет размер более 2,6 млн п.н. Уникальные гены У человека, по разным оценкам, 30-50 тыс. генов. Большинство генов - уникальны. Но даже в них есть повторяющиеся элементы. Это - некоторые экзоны. Все гены разделяют на гены "домашнего хозяйства" и гены "роскоши". Гены "домашнего хозяйства" кодируют то, что всегда нужно любой клетке независимо от ткани. По разным оценкам таких генов у человека 10-20 тыс. Это гистоновые гены, гены tРНК, rРНК и т.п. Гены "роскоши", которых заведомо больше в 2-3 раза, это гены, которые экспрессируются в клетках определенных тканей и в определенное время. Например, все гены белковых гормонов - гены "роскоши". Иная классификация генов: 1. Уникальные гены, имеющие специализированную функцию. Например, глобиновый, инсулиновый и другие гены. Они экспрессируются лишь в определенных клетках. 2. Уникальные гены, обладающие общими функциями, экспрессирующиеся в подавляющем большинстве клеток. Эти гены плохо изучены. 3. Множественные сгруппированные гены. Это гены rРНК, часть генов tРНК, часть гистоновых генов. 4. Множественные рассеянные гены. Это оставшаяся часть гистоновых генов, оставшиеся гены tРНК и большинство генов sРНК, а так же МДГ (мобильные диспергированные (рассеяные) гены).
Та же информация но другими словами внизу.
По
месту локализации генов в структурах
клетки различают расположенные в
хромосомах ядра ядерные гены и
цитоплазматические гены, локализация
которых связана с хлоропластами и
митохондриями.
По
функциональному значению различают
структурные гены, характеризующиеся
уникальными последовательностями
нуклеотидов, кодирующих свои белковые
продукты, которые можно идентифицировать
с помощью мутаций, нарушающих функцию
белка, и регуляторные гены —
последовательности нуклеотидов, не
кодирующие специфические белки, а
осуществляющие регуляцию действия гена
(ингибирование, повышение активности
и др.).
По
влиянию на физиологические процессы в
клетке различают летальные, условно
летальные, супервитальные гены,
гены-мутаторы, гены-антимутаторы и
др.
Следует
отметить, что любые биохимические и
биологические процессы в организме
находятся под генным контролем. Так,
деление клеток (митоз, мейоз) контролируется
несколькими десятками генов; группы
генов осуществляют контроль восстановления
генетических повреждений ДНК (репарация).
Онкогены и гены — супрессоры опухолей
участвуют в процессах нормального
деления клеток. Индивидуальное развитие
организма (онтогенез) контролируется
многими сотнями генов. Мутации в генах
приводят к измененному синтезу белковых
продуктов и нарушению биохимических
или физиологических процессов.
Гомеозисные
мутации у дрозофилы позволили открыть
существование генов, нормальной функцией
которых является выбор или поддержание
определенного пути эмбрионального
развития, по которому следуют клетки.
Каждый путь развития характеризуется
экспрессией определенного набора генов,
действие которых приводит к появлению
конечного результата: глаза, голова
грудь, брюшко, крыло, ноги и т. д.
Исследования генов комплекса bithorax
дрозофилы американским генетиком
Льюисом показали, что это гигантский
кластер тесно сцепленных генов, функция
которых необходима для нормальной
сегментации груди (thorax) и брюшка (abdomen).
Подобные гены получили название
гомеобоксных. Гомеобоксные гены
расположены в ДНК группами и проявляют
свое действие строго последовательно.
Такие гены обнаружены и у млекопитающих,
и они имеют высокую гомологию (сходство).
Функции генов
В
процессе реализации наследственной
информации, заключенной в гене, проявляется
целый ряд его свойств. Определяя
возможность развития отдельного
качества, присущего данной клетке или
организму, ген характеризуется
дискретностью действия (от лат. discretus —
разделенный, прерывистый), прерывностью
(интроны и экзоны). Дискретность
наследственного материала, предположение
о которой высказал еще Г. Мендель,
подразумевает делимость его на части,
являющиеся элементарными единицами, -
гены. В настоящее время ген рассматривают
как единицу генетической функции. Он
представляет собой минимальное количество
наследственного материала, которое
необходимо для синтеза тРНК, рРНК или
полипептида с определенными свойствами.
Ген несет ответственность за формирование
и передачу по наследству отдельного
признака или свойства клеток, организмов
данного вида. Кроме того, изменение
структуры гена, возникающее в разных
его участках, в конечном итоге приводит
к изменению соответствующего элементарного
признака.
Ввиду
того что в гене заключается информация
об аминокислотной последовательности
определенного полипептида, его действие
является специфичным. Однако в некоторых
случаях одна и та же нуклеотидная
последовательность может детерминировать
синтез не одного, а нескольких полипептидов.
Это наблюдается в случае альтернативного
сплайсинга у эукариот и при перекрывании
генов у фагов и прокариот. Очевидно,
такую способность следует оценить как
множественное, или плейотропное, действие
гена (хотя традиционно под плейотропным
действием гена принято понимать участие
его продукта – полипептида – в разных
биохимических процессах, имеющих
отношение к формированию различных
сложных признаков). Например, участие
фермента в ускорении определенной
реакции (см. рис.), которая является
звеном нескольких биохимических
процессов, делает зависимыми результаты
этих процессов от нормального
функционирования гена, кодирующего
этот белок. Нарушение реакции A→B,
катализируемой белком α, в результате
мутации гена ведет к выключению
последующих этапов формирования
признаков D и E.

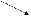 α
D
α
D
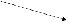
 A
B
A
B
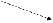 C
E
Определяя
возможность транскрибирования мРНК
для синтеза конкретной полипептидной
цепи, ген характеризуется дозированностью
действия, т.е. количественной зависимостью
результата его экспрессии от дозы
соответствующего аллеля этого гена.
Примером может служить зависимость
степени нарушения транспортных свойств
гемоглобина у человека при
серповидно-клеточной анемии от дозы
аллеля НЬS. Наличие в генотипе человека
двойной дозы этого аллеля, приводящего
к изменению структуры β-глобиновых
цепей гемоглобина, сопровождается
грубым нарушением формы эритроцитов и
развитием клинически выраженной картины
анемии вплоть до гибели. У носителей
только одного аллеля НЬS при нормальном
втором аллеле лишь незначительно
изменяется форма эритроцитов и анемия
не развивается, а организм характеризуется
практически нормальной жизнеспособностью.
C
E
Определяя
возможность транскрибирования мРНК
для синтеза конкретной полипептидной
цепи, ген характеризуется дозированностью
действия, т.е. количественной зависимостью
результата его экспрессии от дозы
соответствующего аллеля этого гена.
Примером может служить зависимость
степени нарушения транспортных свойств
гемоглобина у человека при
серповидно-клеточной анемии от дозы
аллеля НЬS. Наличие в генотипе человека
двойной дозы этого аллеля, приводящего
к изменению структуры β-глобиновых
цепей гемоглобина, сопровождается
грубым нарушением формы эритроцитов и
развитием клинически выраженной картины
анемии вплоть до гибели. У носителей
только одного аллеля НЬS при нормальном
втором аллеле лишь незначительно
изменяется форма эритроцитов и анемия
не развивается, а организм характеризуется
практически нормальной жизнеспособностью.