

37) Перечислите противоречия современной науки.
Миг наибольшего торжества науки, свидетельствовавший о ее мощи, был в то же время началом ее кризиса, потому что создание и применение атомного оружия вело к разрушению и уничтожению. Затем возникла экологическая проблема. Виновны в ней не столько сама наука, сколько цели, которые перед ней ставились, а также нормы, методы и средства, в соответствии с которыми она развивалась. Еще древние философы делили все утверждения на знание и мнение. Знание, или наука, по Аристотелю, может быть двух родов — либо демонстративным, либо интуитивным. Демонстративное знание представляет собой знание причин. Оно состоит из утверждений, которые могут быть доказательствами, т. е. демонстративное знание — это заключения силлогистическими доказательствами, или демонстрациями. Интуитивное знание состоит в схватывании «неделимой формы», сущности или сущностной природы вещи. Интуитивное знание является первоначальным источником всей науки, поскольку оно формирует первоначальные «базисные посылки» для всех доказательств (демонстраций). «Для всего без исключения доказательства быть может, ведь иначе приходилось бы идти в бесконечность», — писал Аристотель «Метафизике». Современные методологи науки принимают это положение и соглашаются идти в бесконечность. «Другими словами, мы знаем, что наши научные теории навсегда должны остаться только гипотезами, но во многих возможных случаях мы можем выяснить, новая гипотеза лучше старой или нет. Дело в том, что если они различны, то они должны вести к различным предсказаниям, которые, как правило, можно проверить экспериментально. На основе такого решающего эксперимента иногда можно обнаружить, что новая теория приводит к удовлетворительным результатам там, где старая оказалась несостоятельной. В результате можно сказать, что в поиске истины мы заменили научную достоверность научным прогрессом. Дело в том, что наука развивается не путем постепенного накопления энциклопедической информации, как думал Аристотель, а движется значительно более революционным путем. Она прогрессирует благодаря смелым идеям, выдвижению новых, все более странных теорий (таких, как теория, по которой Земля не плоская, и «метрическое пространство» не является плоским) и ниспровержению прежних теорий. Однако такой подход к научному методу означает, что в науке нет «знания» в том смысле, в котором понимали это слово Платон и Аристотель, т. е. в том смысле, в котором оно влечет за собой окончательность. В науке мы никогда не имеем достаточных оснований для уверенности в том, что мы уже достигли истины. То, что мы называем «научным знанием», как правило, не является знанием в платоновско-аристотелевском смысле, а, скорее, представляют собой информацию, касающуюся различных соперничающих гипотез и способа, при помощи которого они выдерживают разнообразные проверки. Это, если использовать язык Платона и Аристотеля, информация, касающаяся самого последнего и наилучшим образом проверенного научного «мнения». Такое воззрение означает также, что в науке не существует доказательств (за исключением, конечно, чистой математики и логики). В эмпирических науках, а только они и могут снабжать нас информацией о мире, в котором мы живем, вообще нет доказательств, если под «доказательством» имеется в виду аргументация, которая раз и навсегда устанавливает истинность теории. «А вот что здесь есть, так это опровержения научных теорий» (Поппер К. Открытое общество и его враги. Т. II, М., 1992, с. 20-21). К этому добавляются еще и противоречия, имеющие место внутри самого процесса познания. Природа едина, а науки разделены на отдельные дисциплины. В природе все связано со всем, каждая наука занимает свою полочку. «Существуют отдельные науки., а не наука вообще как наука о действительном, однако каждая из них входит в мир, беспредельный, но все-таки единый в калейдоскопе связей» (Ясперс К. Смысл и назначение истории. М., 1994, с. 102-103). Объекты действительности функционируют как целостные образования, а наука развивается путем абстрагирования некоторых свойств этих объектов, принимаемых за наиболее важные. Основой структуры научного познания (что особенно характерно для наиболее развитых отраслей естествознания) является анализ предмета исследования, т. е. выделение абстрактных элементарных объектов и последующий синтез из этих абстрактных элементов единого целого в форме теоретической системы. По мнению Б. Рассела, «научный прогресс осуществляется благодаря анализу и искусственной изоляции. «Возможно, как считает квантовая теория, что существуют границы правомерности этого процесса, но, если бы он не был обычно правильным, хотя бы приблизительно, научное познание было бы невозможно» (Рассел Б. Человеческое познание. Его сфера и границы. М., 1957, с. 71). Ситуация в области исследования экологической проблемы в практическом плане, как и ситуация в квантовой механике в теоретической, ставит под вопрос правомерность абсолютизации процесса искусственной изоляции и анализа, и многие ученые именно эти черты науки считают ответственными за экологические трудности. С критикой аналитической направленности науки в последнее время приходится сталкиваться все чаще. Эта ее черта признана фундаментальной и оценивалась по большей части положительно в истории науки, хотя известна и другая ее оценка. Критиковали аналитическую направленность науки Гете, Монтень и другие писатели, ученые, философы. С аналитического расчленения Универсума начинается наука. Как пишет В. Вайскопф, «наука стала развиваться, когда люди начали удерживать себя от общих вопросов, таких как: «Из чего состоит материя? Как возникла Вселенная? В чем сущность жизни?» Они стали задавать вопросы частного характера, например: «Как падает камень? Как вода течет по трубе?» и т. д.» (Вайскопф В. Физика в двадцатом столетии. М., 1977, с. 256). В областях, которые наиболее доступны аналитическому расчленению, как, например, физика, наука, достигает наибольшего успеха, и эти области становятся как бы эталонами знания. Мечтой Т. Гоббса было свести все науки к физике, а Ф. Бэкон называл физику «матерью наук». В ХХ веке эти мечты воплотились в методологической концепции «единой науки», которая возникла бы на базе физики (физикализм). Программа сведения всего научного познания к физическому, получившая название редукционизма, не могла быть воплощена в жизнь, поскольку каждая область реальности обладает своей спецификой и не может быть сведена ни к какой другой. Здесь уместно отметить, что аналитизм, лежащий в самом фундаменте научного подхода к действительности, вполне отвечает стремлению человека практически овладеть предметным миром, поскольку сама преобразовательная деятельность в своей сущности также преимущественно аналитична. С этой точки зрения вполне понятно восхищение аналитическим методом (и физикой, в которой этот метод наиболее полно воплотился), которое испытывал Ф. Бэкон. Конечно, делать отсюда вывод, что с помощью науки нельзя познать действительность или что наука ничего не дает для решения фундаментальных проблем человеческого существования, значит впадать в крайность. Выигрыш в четкости познания деталей в общем случае не обязательно должен вести к проигрышу в точности познания целостной картины мира. Но не следует забывать об упоминавшемся относительном характере научных истин, находящем свое выражение в следующем парадоксе познания: знание в наиболее четкой и логичной форме достигается через науку и в более общем плане — через рациональное мышление, но оно в определенной мере и ответственно за разрушение (по крайней мере, идеальное) мира. Итак, один из гносеологических корней экологического кризиса — чрезмерный аналитизм научного мышления, который в стремлении все дальше проникнуть в глубь вещей таит в себе опасность отхода от реальности, от целостного взгляда на природу. Искусственная изоляция какого-либо фрагмента реальности дает возможность его углубленного изучения, однако при этом не учитываются связи этого фрагмента с его средой, и данное обстоятельство, которое может оказаться малосущественным в рамках конкретного исследования, влечет за собой важные негативные последствия, когда результаты подобного исследования вовлекаются в практику человеческой преобразовательной деятельности. Аналитизм внутри конкретных научных дисциплин находит свое продолжение в аналитической направленности развития науки в целом как особой формы постижения мира. Фундаментальной особенностью структуры научной деятельности, вытекающей из ее преимущественно аналитического характера, является разделенность науки на обособленные друг от друга дисциплины. Это, конечно, имеет свои положительные стороны, поскольку дает возможность изучать отдельные фрагменты реальности, но при этом упускаются из виду связи между отдельными фрагментами, а в природе, как известно, «все связано со всем». А каждый акт изменения человеком природной среды не ограничивается какой-либо одной ее областью, а имеет, как правило, большие отдаленные последствия. Разобщенность наук особенно мешает сейчас, в эпоху быстротекущей дифференциации научного знания, выявилась необходимость комплексных интегративных исследований. Чрезмерная специализация так не может помешать эволюции науки, как чрезмерная специализация животных приводит к созданию тупиковых направлений в биологической эволюции.
Тема4.
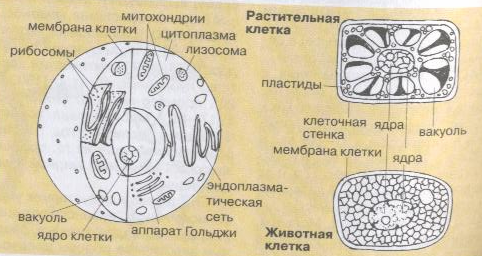
1.Какие основные элементы и вещества слагают живую клетку?
Клетка – самая маленькая организованная единица организмов растений и животных микроскопической величины; состоит из цитоплазмы, в которой находятся клеточные органоиды, и ядра; покрыта оболочкой, у растений клеточной стенкой.
2.Физико-химическая структура ДНК
Дезоксирибонуклеи́новая кислота́ (ДНК) — один из двух типов нуклеиновых кислот, обеспечивающих хранение, передачу из поколения в поколение и реализациюгенетическойпрограммы развития и функционированияживых организмов. Основная роль ДНК вклетках— долговременное хранениеинформациио структуреРНКибелков.
В клетках эукариот(например,животныхилирастений) ДНК находится вядре клеткив составехромосом, а также в некоторых клеточных органоидах (митохондрияхипластидах). В клеткахпрокариотических организмов(бактерийиархей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например,дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемыеплазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовыватьгеномДНК-содержащихвирусов.
С химической точки зрения, ДНК — это длинная полимернаямолекула, состоящая из повторяющихся блоков,нуклеотидов. Каждый нуклеотид состоит изазотистого основания, сахара (дезоксирибозы) ифосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы и фосфатной группы. В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула спирализована. В целом структура молекулы ДНК получила название «двойной спирали».
В ДНК встречается четыре вида азотистых оснований (аденин,гуанин,тиминицитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепиводородными связямисогласнопринципу комплементарности: аденин соединяется только с тимином, гуанин — только с цитозином. Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счет копирования последовательности ДНК в последовательность РНК, синтезируемой в процессетранскрипциии принимают участие в биосинтезе белков (процессетрансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции. Кроме того, в геноме эукариот часто встречаются участки, принадлежащие "генетическим паразитам", например,транспозонам.
Расшифровка структуры ДНК (1953 г.) стала одним из поворотных моментов в истории биологии. За выдающийся вклад в это открытие Фрэнсису Крику,Джеймсу Уотсону,Морису Уилкинсубыла присужденаНобелевская премия по физиологии и медицине1962 г.
3.Свойства генетического кода.
Генетический код— соответствие между триплетами в ДНК (или РНК) и аминокислотами белков.
Свойства генетического кода
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно.
Дискретность — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов.
Специфичность — определённый кодон соответствует только одной аминокислоте.
Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусовдочеловека(на этом основаны методыгенной инженерии)
4.Объясните что такое: репликация, транскрипция, трансляция.
Транскри́пция — процесс синтеза РНКс использованиемДНКв качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Транскрипция катализируетсяферментомДНК-зависимой РНК-полимеразой. Процесс синтеза РНК протекает в направлении от 5'- к 3'- концу, то есть по матричной цепи ДНКРНК-полимеразадвижется в направлении 3'->5'[1]
Транскрипция состоит из стадий инициации, элонгации и терминации.
Инициация транскрипции — сложный процесс, зависящий от последовательности ДНК вблизи транскрибируемой последовательности (а у эукариоттакже и от более далеких участков генома — энхансеров и сайленсеров) и от наличия или отсутствия различныхбелковых факторов.
Момент перехода РНК-полимеразы от инициации транскрипции к элонгации точно не определен. Три основных биохимических события характеризуют этот переход в случае РНК-полимеразы кишечной палочки: отделение сигма-фактора, перваятранслокациямолекулыферментавдоль матрицы и сильная стабилизация транскрипционного комплекса, который кроме РНК-полимеразы включает растущую цепь РНК и транскрибируемую ДНК. Эти же явления характерны и для РНК-полимераз эукариот. Переход от инициации к элонгации сопровождается разрывом связей между ферментом,промотором, факторами инициации транскрипции, а в ряде случаев — переходом РНК-полимеразы в состояние компетентности в отношении элонгации (например,фосфорилированиеCTD-доменау РНК-полимеразы II ). Фаза элонгации заканчивается после освобождения растущего транскрипта идиссоциациифермента от матрицы (терминация).
На стадии элонгации в ДНКрасплетено примерно 18 парнуклеотидов. Примерно 12 нуклеотидов матричной нити ДНК образует гибридную спираль с растущим концом цепи РНК. По мере движения РНК-полимеразы по матрице впереди нее происходит расплетание, а позади — восстановление двойной спирали ДНК. Одновременно освобождается очередное звено растущей цепи РНК из комплекса с матрицей и РНК-полимеразой. Эти перемещения должны сопровождаться относительным вращением РНК-полимеразы и ДНК. Трудно себе представить, как это может происходить в клетке, особенно при транскрипциихроматина. Поэтому не исключено, что для предотвращения такого вращения двигающуюся по ДНК РНК-полимеразу сопровождаюттопоизомеразы.
Элонгация осуществляется с помощью основных элонгирующих факторов , необходимых, чтобы процесс не останавливался преждевременно [2].
В последнее время появились данные, показывающие, что регуляторные факторы также могут регулировать элонгацию. РНК-полимераза в процессе элонгации делает паузы на определенных участках гена. Особенно четко это видно при низких концентрацияхсубстратов. В некоторых участках матрицы длительные задержки в продвижении РНК-полимеразы, т.н. паузы, наблюдаются даже при оптимальныхконцентрацияхсубстратов. Продолжительность этих пауз может контролироваться факторами элонгации
У бактерии бактерий есть два механизма терминации транскрипции:
ро-зависимый механизм, при котором белок Rho (ро), который дестабилизирует водородные связи между матрицей ДНКимРНК, высвобождая молекулу РНК.
ро-независимый, при котором транскрипция останавливается, когда только что синтезированная молекула РНК формирует стебель-петлю, за которой расположено несколькоурацилов(...УУУУ), что приводит к отсоединению молекулы РНК от матрицы ДНК.
Терминация транскрипции у эукариот менее изучена. Она завершается разрезанием РНК, после чего к её 3' концу фермент добавляет несколько аденинов(...АААА), от числа которых зависит стабильность данного транскрипта[3].
Существует ряд экспериментальных данных, свидетельствующих о том, что транскрипция осуществляется в так называемых транскрипционных фабриках: огромных, по некоторым оценкам, до 10 МДа комплексах, которые содержат около 8 РНК-полимераз II и компоненты последующего процессинга и сплайсинга, а также пруф-ридинга новосинтезированного транскрипта [4]. В ядре клетки происходит постоянный обмен между пулами растворимой и задействованной РНК-полимеразы. Активная РНК-полимераза задействована в таком комплексе, который в свою очередь является структурной организовывающей компактизацию хроматина единицей. Последние данные[5]свидетельствуют о том, что транскрипционные фабрики существуют и в отсутствие транскрипции, они фиксированы в клетке (пока не ясно, взаимодействуют ли они с матриксом клетки или нет) и представляют собой независимый ядерный субкомпартмент. Попытки выделить белковый функциональный комплекс транскрипционной фабрики пока не привели к успеху ввиду его огромных размеров и низкой растворимости.
Трансляцией называют осуществляемый рибосомойсинтезбелкаизаминокислотна матрице информационной (или матричной)РНК(иРНК илимРНК). Трансляция является финальной реакциейреализации генетической информации.
Синтез белка является основой жизнедеятельности клетки. Для осуществления этого процесса в клетках всех без исключения организмов имеются специальные органеллы —рибосомы. Рибосомы представляют собойрибонуклеопротеидныекомплексы, построенные из 2 субъединиц: большой и малой. Функция рибосом заключается в узнавании трехбуквенных (трехнуклеотидных)кодоновмРНК, сопоставлении им соответствующих аминокислот и присоединении этих аминокислот к растущей белковой цепи. Двигаясь вдоль молекулы мРНК рибосома распознает кодон за кодоном и синтезирует белок в соответствии с информацией, заложенной в молекуле мРНК.
Для узнавания аминокислот в клетке имеются специальные «адаптеры», молекулы транспортной РНК (тРНК). Эти молекулы, имеющие форму клеверного листа, имеют участок (антикодон), комплементарный кодону мРНК, а также другой участок, к которому присоединяется аминокислота, соответствующая этому кодону. Присоединение аминокислот к тРНК осущетвляется в энегро-зависимой реакции ферментами аминоацил-тРНК-синтетазами, а получившаяся молекула называется аминоацил-тРНК. Таким образом, специфичность трансляции определяется взаимодействием между кодоном мРНК и антикодоном тРНК, а также специфичностью аминоацил-тРНК-синтеназ, присоединяющих аминокислоты строго к соответствующим им тРНК (например, кодону GGU будет соответствовать тРНК, содержащая антикодон CCA, а к этой тРНК будет присоединяться только аминокислотаглицин).
Механизмытрансляциипрокариотиэукариотсущественно отличаются, поэтому многие вещества, подавляющие прокариотическую трансляцию в значительно меньшей степени действуют на трансляцию высших организмов, что позволяет использовать их в медицинской практике как антибактериальные средства безопасные для организма млекопитающих.
Процесс трансляции разделяют на инициацию — узнавание рибосомой стартового кодонаи начало синтеза, элонгацию — собственно синтез белка и терминацию — узнавание терминирующего кодона (стоп-кодона) и отделение продукта.
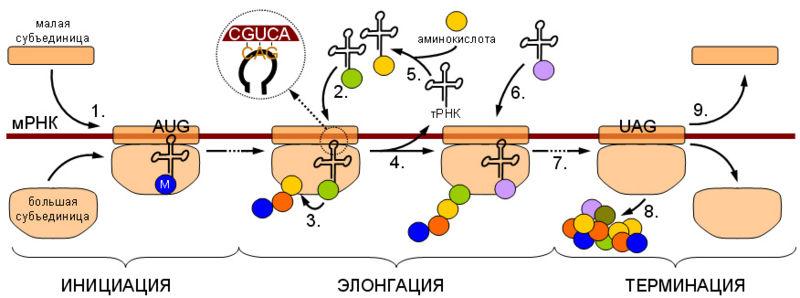
Общая схема трансляции. Инициация. 1. Узнавание стартового кодона (AUG), сопровождается присодинением тРНК аминоацилированной метионином (М) и сборкой рибосомы из большой и малой субъединиц. Элонгация. 2. Узнавание текущего кодона соответствующей ему аминоацил-тРНК (комплементарное взаимодействие кодона мРНК и антикодона тРНК увеличено). 3. Присоединение аминокислоты, принесенной тРНК, к концу растущей полипептидной цепи. 4. Продвижение рибосомы вдоль матрицы, сопровождающееся высвобождением молекулы тРНК. 5. Аминоацилирование высвободившейся молекулы тРНК соответствующей ей аминоацил-тРНК-синтетазой. 6. Присоединение следующей молекулы аминоацил-тРНК, аналогично стадии (2). 7. Движение рибосомы по молекуле мРНК до стоп-кодона (в данном случае UAG). Терминация. Узнавание рибосомой стоп-кодона сопровождается (8) отсоединением новосинтезированного белка и в некоторых случаях (9) диссоциацией рибосомы.
Поскольку каждый кодон содержит три нуклеотида, один и тот же генетический текст можно прочитать тремя разными способами (начиная с первого, второго и третьего нуклеотидов), то есть в трех разных рамках считывания. За некоторыми интересными исключениями, значимой является информация закодированная только в одной рамке считывания. По этой причине крайне важным для синтеза белка рибосомой является ее правильное позиционирование на стартовом AUG-кодоне — инициация трансляции.
Синтез белка в большинстве случаев начинается с AUG-кодона, кодирующегометионин. Этот кодон обычно называют стартовым или инициаторным. Инициация трансляции предусматривает узнаваниерибосомойэтого кодона и привлечение инициаторной аминоацил-тРНК. Для инициации трансляции необходимо также наличие определенных нуклеотидных последовательностей в районе стартового кодона. Существование последовательности, отличающей стартовый AUG от внутренних совершенно необходимо, так как в противном случае инициация синтеза белка происходила бы хаотично на всех AUG-кодонах.
Процесс инициации обеспечивается специальными белками — факторами инициации(англ. initiation factors, сокращенно IF; эукариотические инициаторные факторы обозначают eIF, от англ. eukaryotes).
Механизмы инициации трансляции у про-иэукариотсущественно отличаются: прокариотические рибосомы потенциально способны находить стартовый AUG и инициировать синтез на на любых участках мРНК.
У эукариот существуют два механизма нахождения рибосомой стартового AUG. Наиболее распространен, так называемый сканирующий механизм, при котором рибосома, двигаясь вдоль молекулы мРНК от ее 5'-конца «сканирует» один кодон за другим, пока не наткнется на инициаторный AUG. Для привлечения рибосомы к 5'-концу мРНК требуется специальная структура, кэп — 7-метилгуанин, прикрепленный к 5'-концевому нуклеотиду мРНК. Второй механизм эукариотической инициации трансляции не требует наличия кэп-структуры и позволяет инициировать трансляцию с внутреннего участка мРНК, называется IRES-зависимым механизмом. IRES(от англ. Internal Ribosomal Entry Site, участок внутренней посадки рибосомы) — участок мРНК, обладающий выраженной вторичной стуктурой, позволяющей ему направлять рибосомы на стартовый AUG. По IRES-зависимому механизму инициируется синтез лишь на небольшой части клеточных мРНК, а также на РНК некоторыхвирусов.