

2 курс / Нормальная физиология / ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
.pdfналы для ионов Na+ — натриевые, для ионов К+ — калиевые, для ионов
Cl -— хлорные, для ионов Са2+ — кальциевые и т.д.;
•неспецифические (неселективные) ионные каналы, через которые могут про-
ходить разные ионы. Например, в кардиомиоцитах есть каналы, которые пропускают как ионы натрия, так и ионы кальция. В основном, такие каналы расположены на постсинаптических мембранах синапсов.
По наличию ворот в канале:
•безворотные (каналы утечки), они всегда открыты, и через них пассивно (по концентрационному градиенту) проходят (утекают!) ионы (Рис. 7);
•воротные каналы.
Рис. 7. Безворотный ионный канал
В состоянии функционального покоя клетки они закрыты и открываются только под действием раздражителей.
21

Рис.8. Строение воротного ионного канала
Воротный канал имеет (Рис.8): устье; селективный (избирательный)
фильтр; ворота; механизм управления воротами.
Такие ионные каналы обладают свойством управляемости, т.е. они могут открываться под действием определенных факторов. В зависимости от меха-
низма управления воротами (т.е. на что открываются ворота канала), разли-
чают каналы:
•механочувствительные — реагируют на механическую деформацию мембра-
ны, например, при растяжении;
•хемозависимые — реагируют на химическое вещество, например, медиатор;
•потенциалзависимые — реагируют на изменение заряда на поверхности мем-
браны.
Например, потенциалзависимые Na-каналы имеют ворота 2-х типов — активационные (m-ворота) и инактивационные (h- ворота) и могут находиться в трех состояниях. В состоянии покоя активационные ворота закрыты, а инакти-
вационные открыты.
22

Рис.9. Схема работы потенциалзависимого Naионного канала
В ответ на действие раздражителя и изменение заряда на мембране про-
исходят изменения белковой молекулы канала: в результате активационные во-
рота открываются и ионы проходят через канал в клетку (состояние активации),
после чего начинают закрываться инактивационные ворота и происходит инак-
тивация каналов. Некоторое время спустя закрываются активационные и от-
крываются инактивационные ворота и канал снова приходит в состояние покоя
(Рис.9).
Мембранный потенциал или потенциал покоя
Через клеточную мембрану в клетку и из нее постоянно осуществляется транспорт веществ, в том числе ионов (K+, Na+, Ca2+, Cl-). В результате, каких-то ионов оказывается больше снаружи клетки, а каких-то внутри нее.
В состоянии покоя поверхностная мембрана возбудимой клетки поляри-
зована, то есть внутренняя ее поверхность заряжена отрицательно по отноше-
нию к наружной.
Разность потенциалов между наружной и внутренней поверхностями мембраны — это и есть мембранный потенциал (МП) или трансмембранная разность.
Для объяснения механизма возникновения и поддержания МП было раз-
работано несколько теорий. В 1949–1952 гг. Ходжкин, Хаксли и Кац модифи-
23
цировали существовавшие ранее теории и экспериментально обосновали ос-
новные положения мембранно-ионной теории. Согласно этой теории, проис-
хождение МП связано с двумя основными условиями, которые должны иметь место в состоянии покоя в любой возбудимой клетке. Это:
•наличие ионной асимметрии по обе стороны поверхностной клеточной мем-
браны;
•наличие разной проницаемости поверхностной клеточной мембраны для раз-
личных ионов.
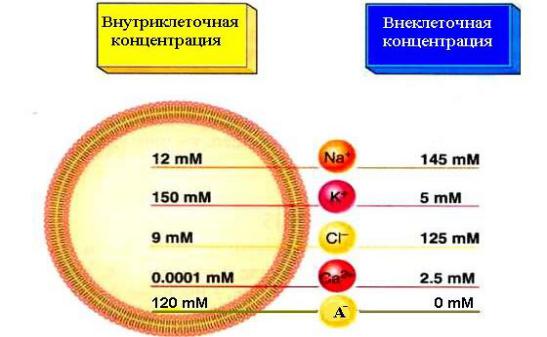
Ионная асимметрия — это разная концентрация ионов по обе стороны
поверхностной мембраны клетки (Рис.10). Так,
•ионов К+ находится в клетке в 30–50 раз больше, чем снаружи;
•ионов Na+ больше находится вне клетки, чем внутри нее в 8–10 раз;
•ионов Са2+ примерно в 20 раз больше вне клетки;
•ионов Cl- больше во внеклеточной жидкости, чем внутри клетки в 50 раз;
•внутри клетки находится больше органических анионов, по сравнению с наружной поверхностью.
Рис. 10. Ионная асимметрия клеточной мембраны
24

Таким образом, для этих ионов направление концентрационного градиен-
та (из большей концентрации к меньшей) различно: для К+ — из клетки, для Na+, Са2+ и Cl- — в клетку.
Вторая причина, обусловливающая поляризацию мембраны — это раз-
личная проницаемость мембраны для различных ионов.
Так, в состоянии покоя мембрана в 25 раз более проницаема для ионов К+, чем для Na+ за счет наличия каналов утечки для этих ионов. Кроме того, ка-
лиевых каналов на единицу площади мембраны намного больше, чем натрие-
вых.
Механизм возникновения мембранного потенциала (потенциала покоя)
Поскольку в покое внутри клетки содержится большое количество ионов К+ и А- , а мембрана высокопроницаема для ионов К+ , то К+. начинает пассивно выходить по градиенту концентрации из клетки через свои каналы утечки
(Рис. 11).
Рис.11. Ток калия через клеточную мембрану в покое
Ионы К+.выносят положительные заряды и группируются у наружной по-
верхности мембраны, поэтому снаружи!!! мембрана заряжается положитель-
но!!!, а потеря положительных зарядов клеткой ведет к появлению отрица-
тельного!!! заряда на внутренней!!! поверхности мембраны.
Ионы калия, находящиеся внутри клетки, связаны с органическими анио-
нами (Рис. 12 А). Когда ионы калия выходят из клетки по концентрационному градиенту, то органические анионы за счет сил электростатического взаимо-
действия «стремятся» выйти вслед за ними. Но их большой размер и отрица-
25

тельный заряд (а внутренние стенки ионных каналов тоже заряжены отрица-
тельно!) не позволяют им даже войти в канал. Поэтому анионы остаются на внутренней поверхности мембраны, заряжая мембрану изнутри отрицатель-
но!!!, и за счет электростатического взаимодействия удерживают ионы калия!!!
на наружной поверхности мембраны. В результате этого на мембране формиру-
ется разность потенциалов (рис. 12 Б).
А Б Рис. 12. Роль ионов калия в создании мембранного потенциала покоя
Ионы натрия также участвуют в создании МП, так как небольшие порции натрия по каналам утечки постоянно проникают в клетку, и тем самым на
15–20% уменьшают величину МП, созданного ионами калия. В создании МП принимают участие и другие ионы, но их роль незначительна.
Таким образом, калий является основным ионом, обеспечивающим фор-
мирование МП (трансмембранную разность)!!!
Величина МП в покое у разных клеток различна и колеблется, например,
для волокон поперечнополосатой мускулатуры от -60 до -90 мВ, для волокон гладкой мышечной ткани от -30 до -50 мВ, для нервных клеток от -60 до -70
мВ. Знак «-» указывает на преобладание отрицательного заряда внутри клетки.
Поддержание постоянства этого значения осуществляется специальным механизмом, который обеспечивает обратное перемещение ионов К+ и Na+ про-
тив градиента их концентрации, т.е. активный транспорт. Таким механизмом является Na+-К+ — обменный насос, который при затрате энергии 1 молекулы АТФ выкачивает 3 иона Na+ из клетки и одновременно закачивает 2 иона К+
внутрь клетки!!! (Рис.13).
26

Рис. 13. Механизм работы натрий-калиевого насоса
В результате, положительно-заряженных частиц выкачивается больше,
чем закачивается, и значение МП сохраняется со знаком «-». В тоже время натрий-калиевый насос поддерживает ионную асимметрию Na+ и К+ внутри клетки.
Таким образом, МП представляет собой разность потенциалов между наружной и внутренней поверхностью мембраны клетки. Он является результа-
том разделения зарядов относительно клеточной мембраны, которое возникает за счет движения заряженных ионов (калия, и частично натрия) по концентра-
ционным градиентам через ионные каналы, открытые в покое, и за счет работы
Na+ — К+ насоса.
Потенциал действия
Возбудимые ткани осуществляют переработку и передачу информации с помощью биопотенциалов, которые подразделяются на: распространяющие-
ся — потенциал действия (ПД); местные — локальный ответ (ЛО).
Местные и распространяющиеся потенциалы переходят друг в друга,
обеспечивая единый процесс обработки и передачи информации.
К распространяющемуся биопотенциалу относится потенциал действия
(нервный импульс, возбуждение), который возникает при действии на возбуди-
27

мую клетку порогового или надпорогового раздражителя. В результате проис-
ходит передача информации (т.е. возбуждения) по этой клетке.
Потенциал действия (ПД) — это быстрое колебание мембранного по-
тенциала клетки в ответ на ее раздражение, сопровождающееся изменением знака заряда на мембране, обусловленное изменением проницаемости клеточ-
ной мембраны и диффузией ионов в клетку и из нее.
Потенциал действия (процесс возбуждения) протекает в две фазы:
I.фаза деполяризации (уменьшение МП).
II.фаза реполяризации (восстановление МП).
Фаза деполяризации
Согласно мембранно-ионной теории при действии раздражителя порого-
вой силы на мембране открываются Na-каналы (Рис. 14).
Рис. 14. Ток ионов натрия через мембрану клетки при действии раздражителя
В результате начинается пассивное проникновение натрия внутрь клетки.
Натрий заносит в клетку положительный заряд, и уровень МП начинает посте-
пенно уменьшаться, т.е. на мембране начинается процесс деполяризации. Это в свою очередь способствует открытию еще большего количества управляемых натриевых каналов. Таким образом, деполяризация нарастает и достигает тако-
28

го уровня, когда одновременно открываются (!) все управляемые натриевые ка-
налы, и Na+ лавинообразно устремляется в клетку! Такой уровень деполяриза-
ции поверхностной мембраны называется «критическим уровнем деполяриза-
ции» (КУД).
При лавинообразном токе натрия в клетку деполяризация мембраны быстро нарастает, доводя трансмембранную разность сначала до 0, а затем мембрана внутри заряжается положительно, а снаружи — отрицательно
(Рис. 15).
Рс.15. Перезарядка мембраны в результате процесса деполяризации
Такое состояние мембраны называется перезарядка, или реверсия, или ин-
версия, или овершут. Таким образом, деполяризация мембраны завершается ее перезарядкой!!! (Рис.16).
Но поступление ионов натрия в клетку не может быть бесконечным. В
момент достижения деполяризации пикового потенциала (спайка) наступает
инактивация натриевых каналов!!! Кроме того, проникшие в клетку ионы натрия создают электрическое поле, препятствующее дальнейшему поступле-
нию натрия. С этого момента начинается фаза реполяризации.
29

Рис.16. Фазы потенциала действия
Фаза реполяризации — это восстановление исходной степени поляриза-
ции мембраны после ее деполяризации.
Реполяризация осуществляется 2-мя механизмами:
1)выходом калия из клетки;
2)работой натрий-калиевого насоса.
В ответ на возникшую перезарядку клеточной мембраны одновременно с закрытием (инактивацией) натриевых каналов открываются потенциалзависмые калиевые каналы. За счет выхода калия из клетки выносится избыток положи-
тельного заряда, привнесенного натрием, и происходит восстановление исход-
ной степени поляризации мембраны. Кроме этого в фазу реполяризации макси-
мально активизируется натрий-калиевый насос, который начинает активно вы-
качивать из клетки натрий, зашедший в фазу деполяризации.
Фаза реполяризации ПД обычно заканчивается следовыми явлениями.
Следовые явления или потенциалы — это восстановительные процессы,
медленно развивающимися на мембране после ее возбуждения. Так, если про-
30
Рекомендовано к изучению сайтом МедУнивер - https://meduniver.com/