

2 курс / Нормальная физиология / Общий_курс_физиологии_человека_и_животных_Том_1_Ноздрачев_А_Д_,
.pdfвозможны не только вегетативные реакции, но и целый ряд соматических эффектов, таких, как изменение позы или увеличение частоты дыхания.
Эти факты свидетельствуют о том, что гипоталамус вряд ли является статическим и локальным объединением эрготропной и трофотропной зон. Предпочтительнее представлять себе гипоталамус как важный интегративпый центр вегетативных, соматических и эндокринных функций, который отвечает за реализацию сложных гомеостатических реакций и входит в иерархически организованную систему отделов головного мозга, регулирующих висцеральные функции.
Системность гомеостатических реакций можно продемонстрировать опять-таки на примере регуляции гемодинамики. Известно, что саморегуляция сосудистого тонуса осуществляется за счет функции сосудистого центра продолговатого мозга, который работает как следящая система. Этот уровень интеграции достаточен для осуществления простых рефлекторных реакций в ответ на информацию от баро- и механорецепторов сосудистого русла. Вместе с тем более сложные вазомоторные реакции, связанные, например, с терморегуляцией или локомоторными актами, реализуются с участием гипоталамуса, который связан с сосудистым центром и с сосудодвигательными нейронами спинного мозга.
В то же время сам гипоталамус имеет связи с корой головного мозга, представляющей еще более высокий уровень интеграции. Такие связи обнаружены, например, между корой и латеральной областью гипоталамуса, отвечающей за приспособительные реакции сосудистой системы при физической нагрузке. Очевидно, по этим путям распространяются модулирующие влияния коры на деятельность гипоталамуса.
Таким образом, в регуляцию вегетативных реакций вовлекается целая система центров, представленных на всех уровнях головного мозга. Гипоталамус является одним из уровней данной системы, и это во многом определяет сложность и адаптивный характер регулируемых им вегетативных реакций.
3.7.5. Терморегуляционная функция гипоталамуса
Целый ряд экспериментальных и клинических данных указывает на то, что гипоталамус является интегративным центром терморегуляции (см. разд. 10.4.5). Так,
например, перерезка стволовой части мозга у кошки на уровне верхней границы среднего мозга нарушает терморегуляцию и превращает теплокровное животное в холоднокровное. Локальное раздражение задних отделов гипоталамуса показало, что здесь находятся структуры, регулирующие процесс теплопродукции. Эффект от раздражения проявляется в интенсификации обменных процессов, увеличении частоты сердечных сокращений и появлении дрожания скелетной мускулатуры. Разрушение задних отделов гипоталамуса вызывает подавление теплопродукции и снижение температуры тела.
Передние отделы гипоталамуса, в частности паравентрикулярные ядра, ответственны за интеграцию процессов теплоотдачи. Раздражение этой области вызывает расширение кожных сосудов, усиление потоотделения, интенсификацию дыхания. Повреждение переднего гипоталамуса сопровождается нарушением теплоотдачи и повышением температуры организма — гипертермией. При регистрации электрической активности одиночных клеток в переднем гипоталамусе были обнаружены так называемые тепловые нейроны, у которых локальное нагревание вызывает увеличение частоты импульсной активности. Как правило, усиление разрядов этих нейронов предшествует комплексу реакций, увеличивающих теплоотдачу.
Тепловые нейроны переднего гипоталамуса могут быть отнесены к категории внутренних терморецепторов, которые в естественных условиях реагируют на температуру крови и вместе с кожными терморецепторами рефлекторно возбуждают эфферентные нейроны гипоталамуса, контролирующие теплоотдачу на периферии.
3.7.6. Участие гипоталамуса в регуляции поведенческих реакций
Локальные электрические раздражения определенных зон гипоталамуса могут вызывать направленные на выживание особи поведенческие комплексы, которые включают моторные, вегетативные и гормональные компоненты. Так, в заднем гипоталамусе обнаружена область, электрическая стимуляция которой вызывает комплекс реакций, характерных для пищевого поведения: поиск пищи, обильное слюноотделение, усиленная моторика и кровоснабжение кишечника, снижение мышечного кровотока.
Б. Ананду и Д. Бробеку (1951) удалось идентифицировать в гипоталамусе взрослых крыс несколько зон, ответственных за пищевое поведение. Они показали, что повреждение небольшого участка в латеральном гипоталамусе ведет к полному отказу от пищи (афагия) и воды (адипсия), истощению и гибели животного. Эта зона была названа авторами «зоной голода».
В то же время разрушение вентромедиальных ядер гипоталамуса вызывает чрезмерное потребление пищи (гиперфагию) и ожирение. Здесь локализован «центр насыщения», нейроны которого, по всей вероятности, обладают прямой химической чувствительностью к составу крови, зависимому от периодичности питания. Согласно гипотезе Б. Ананду и Д. Бробека, нейроны центра насыщения по мере потребления пищи активируются и через внутригипоталамические тормозные связи подавляют функции центра голода, что, как следствие, вызывает устранение комплекса реакций, связанных с пищевым поведением.
Эта привлекательная гипотеза в целом еще не получила экспериментального подтверждения, так как тормозные взаимодействия между центрами не идентифицированы, да и само понятие центров голода и насыщения является скорее абстрактным, нежели соотнесенным с представлением о четко очерченных анатомических структурах.
Дело в том, что области гипоталамуса, связанные с конкретными поведенческими реакциями, широко перекрываются и зачастую их удаление вызывает содружественные нарушения общего поведения животных. Например, разрушение латерального гипоталамуса ведет помимо афагии еще и к снижению двигательной активности животных, угнетению эмоций, ослаблению устойчивости к стрессу. Эти факты объясняются тем, что при повреждении латерального гипоталамуса нарушаются функции не только одной изолированной системы. Как известно, через латеральный гипоталамус проходят волокна мощного медиального пучка переднего мозга, который связывает между собой базальные отделы переднего мозга, гипоталамус и покрышку среднего мозга. Сюда же от стволовой части мозга подходят восходящие дофаминэргические и норадренэргические пути, поражение которых вызывает комплекс нарушений, сходных с теми, которые возникают при повреждении латерального гипоталамуса.
Таким образом, латеральный гипоталамус является общим путем, в котором сходятся воздействия разнообразных интегративных систем мозга, и нет ничего удивительного, что его повреждение вызывает комплексные нарушения поведенческих реакций.
В этой связи следует отметить, что в медиальном пучке переднего мозга, так же как и в заднем гипоталамусе, обнаружены зоны, связанные с регуляцией полового поведения. В 1954 г. Дж. Олдс и П. Милнер впервые показали, что крысы с хронически вживленными в эти зоны электродами довольно быстро обучались нажимать на рычаг, который включает электрическое раздражение их собственного мозга. Такое производимое по собственному почину раздражение действует как «вознаграждение». Если животным предоставлялся свободный выбор, то голодные крысы гораздо быстрее устремлялись к рычагу самостимуляции, чем к лежащей рядом пище. Максимальная частота самораздражения достигала 5000 нажатий рычага за 1 ч.
Естественно, что о субъективных ощущениях животных мы ничего не знаем, и вопрос о возможности использовать эксперименты на животных для объяснения человеческих эмоций долгое время оставался спорным. Однако работы американских ученых С. Сен-
Джекобсона, А. Торкилдсена, X. Дельгадо, проведенные в условиях нейрохирургической клиники, показали, что раздражение аналогичных участков мозга у людей вызывало чувства радости, удовлетворения, сопровождавшиеся эротическими переживаниями.
Очевидно, эти положительные эмоции являются компонентами полового поведения, которое регулируется задним гипоталамусом. Вместе тем судить о локализации здесь «центра удовольствия» несколько опрометчиво, так как система регуляции полового поведения включает и другие отделы головного мозга, в том числе лимбическую кору.
Материалы клинических данных свидетельствуют о том, что гипоталамус определяет
правильную периодичность функций, связанных с размножением. Опухолевые процессы в области гипоталамуса могут вызывать быстрое половое созревание, нарушение менструального цикла, половую слабость и ряд других дисфункций.
Как показали опыты с локальным раздражением, гипоталамус участвует в регуляции агрессивного поведения животных. При раздражении передних отделов гипоталамуса кошка принимает угрожающую позу, оскаливает зубы, шипит и выпускает когти. Эта реакция сопровождается вегетативными компонентами — увеличением частоты сердечных сокращений и пиломоторным эффектом. В связи с тем, что данная реакция не имеет объекта агрессии, она называется ложной яростью.
Иная картина наблюдается при раздражении некоторых боковых отделов гипоталамуса. В данном случае агрессия имеет четкую направленность против контрольного животного, без лишних движений и агрессивных демонстраций. Эти факты говорят о том, что каждая форма поведения имеет свою эмоциональную окраску, в создании которой гипоталамус как интегративный центр играет существенную роль.
Как регуляторный орган гипоталамус принимает участие в чередовании состояний сна и бодрствования. В клинике описаны случаи перехода в состояние летаргического сна при повреждении гипоталамуса. Сноподобное состояние у животных можно вызвать при электрическом раздражении некоторых зон медиального гипоталамуса. Напротив, задний гипоталамус имеет решающее значение в поддержании состояния бодрствования. Переход от сна к бодрствованию и обратно сопровождается изменением соматических (мышечный тонус) и вегетативных (частота сердечных сокращений, перистальтика кишечника) процессов, интеграция которых осуществляется гипоталамусом.
В процессы регуляции гипоталамусом приспособительных поведенческих реакций входит и его участие в поддержании водного баланса организма. Отсутствие воды создает мощное побуждение (мотивацию), направленное на устранение дефицита жидкости. В создании этой мотивации, в появлении чувства жажды участвует гипоталамус, в передней области которого обнаружены нейроны с осморецепторной функцией. Эти нейроны возбуждаются при изменении осмотического давления крови и запускают целый комплекс соматических и эндокринных реакций, направленных на устранение данной мотивации.
Даже это краткое перечисление регуляторных функций гипоталамуса свидетельствует о его важной роли в организации поведения у животных.
Вместе с тем принцип нейронной организации гипоталамуса до сих пор мало исследован и остается загадкой, как такое маленькое образование способно управлять многими системными реакциями организма. Вероятно, исследование структуры и функции нейронных цепей гипоталамуса обеспечит дальнейший прогресс в нашем познавании этого отдела мозга.
3.7.7. Гипоталамо-гипофизарная система
Одной из важнейших функций гипоталамуса является регуляция деятельности гипофиза (см. разд. 6.2). Гипофиз — главная железа внутренней секреции — состоит из передней, задней и промежуточной долей. В процессе эмбриогенеза передняя доля, или аденогипофиз, развивается из выпячивания глотки, а задняя, или нейрогипофиз, формируется из промежуточного мозга. Как передняя, так и задняя доли гипофиза
находятся под контролем гипоталамуса, однако механизмы их регуляции различны. Нейрогипофиз является органом, депонирующим два гормона: антидиуретический
(АДГ, вазопрессин) и окситоцин. Как уже упоминалось выше, местом синтеза этих гормонов являются супраоптическое и паравентрикулярное ядра гипоталамуса. Формируясь в нейросекреторных клетках этих ядер, гормоны в виде гранул транспортируются по их аксонам, и после разрушения гранул выделяются в капиллярную сеть нейрогипофиза. Антидиуретический гормон регулирует обратное всасывание воды в почечных канальцах и воздействует на гладкую мускулатуру артериол, повышая, таким образом, артериальное давление. Окситоцин стимулирует сокращение гладкой мускулатуры матки и молочных желез.
Регуляция секреции обоих гормонов осуществляется по механизму нейрогуморального рефлекса, афферентное звено которого представлено нервными путями от осморецепторов или механорецепторов до гипоталамуса, а эфферентное звено — поступающим в кровяное русло гормоном. Кроме того, регуляция функций молочной железы может осуществляться на первых порах чисто гуморальным путем за счет изменения гормонального фона во время беременности.
Если гормоны задней доли гипофиза продуцируются нейросекреторными клетками гипоталамических ядер, то все гормоны передней доли секретируются клетками самого аденогипофиза. В зависимости от мишеней, на которые направлено их действие, гормоны аденогипофиза подразделяются на гландотропные, влияющие на другие периферические эндокринные железы, и эффекторные, воздействующие непосредственно на организм. К первой группе относятся: адренокортикотропный гормон (АКТГ) — регулирует секрецию глюкокортикоида кортизола в коре надпочечников; тиреотропный гормон (ТТГ) — стимулирует рост и секрецию щитовидной железы; гонадотропные гормоны — фолликулостимулирующий (ФСГ) и лютеинизирующий (ЛГ) — регулируют деятельность половых желез.
Эффекторные гормоны аденогипофиза представлены соматотропным гормоном (СТГ, гормон роста), регулирующим рост костей, и пролактином, который стимулирует рост молочных желез и секрецию молока. К числу эффекторных гормонов относятся также
меланоцитостимулирующий гормон промежуточной доли гипофиза (МСГ, интермедии),
являющийся регулятором кожной пигментации.
В 70-е гг. было установлено, что секреторная функция аденогипофиза находится под контролем гипоталамуса. Этот контроль осуществляется нейрогуморальным путем за счет гормонов, выделяемых гипофизотропной зоной гипоталамуса (в медиальном гипоталамусе). Оказалось, что секреция всех известных гормонов аденогипофиза регулируется гипофизотропными гормонами гипоталамуса, которые являются как бы гормонами гормонов. Гипофизотропные гормоны представляют собой пептиды с низкой молекулярной массой и подразделяются на стимулирующие, или так называемые рилизинг-факторы (либерины), и тормозящие секрецию ингибирующие факторы
(статины).
Высвобождаясь из нервных окончаний, гипофизотропные гормоны через сосуды гипоталамо-гипофизарной портальной системы попадают в аденогипофиз и там воздействуют на клетки, секретирующие тот или иной тропный гормон. Совсем недавно из гипоталамуса и гипофиза была выделена еще одна группа пептидов — энкефалины и эндорфины, которые оказывают на нервные клетки морфиноподобное действие и, повидимому, играют существенную роль в регуляции вегетативных процессов и поведения.
Секреция гипофизотропных гормонов гипоталамуса регулируется по принципу отрицательной обратной связи. Установлено, что при повышении содержания в плазме крови гормонов периферических желез внутренней секреции уменьшается поступление соответствующих рилизинг-факторов в портальную гипоталамо-гипофизарную систему и тем самым снижается секреция того или иного тропного гормона аденогипофиза.
Параллельно с этим механизмом деятельность гипоталамо-гипофизарной системы может регулироваться и за счет нервных влияний, приходящих от лимбической системы и среднего мозга через латеральный гипоталамус. Так, известно, что при сильных болевых или других стрессорных воздействиях у животных наблюдается усиленное выделение АКТГ и, наоборот, снижение секреции гонадотропных гормонов. Механизм этой реакции обусловлен изменениями в секреции соответствующих рилизинг-факторов, которые вызываются импульсацией, приходящей в гипофизотропную зону гипоталамуса из лимбической коры и среднего мозга. Существует точка зрения, что сигналы от этих центров поступают через дофамин- и норадренэргические пути.
Таким образом, за счет нервных влияний секреция аденогипофиза приводится в соответствие с мотивацией и биологической направленностью реакций организма.
3.8.Лимбическая система
3.8.1.Анатомические структуры лимбической системы
Гипоталамус как нервный, центр, участвующий в формировании биологических побуждений к действию или мотиваций, тесно связан с лимбической системой мозга. Под лимбической системой понимают морфофункциональное объединение, которое включает в себя филогенетически старые отделы коры переднего мозга, а также ряд подкорковых структур, которые регулируют функции внутренних органов, обусловливающих эмоциональную окраску поведения и его соответствие имеющемуся субъективному опыту
(см. разд. 5.5).
В состав лимбической коры входит древняя кора (палеокортекс), формирующая обонятельный мозг и состоящая из обонятельных луковиц, обонятельного бугорка, прозрачной перегородки и прилежащих областей коры (препериформная, периамигдалярная и диагональная области). Следующий компонент лимбической системы
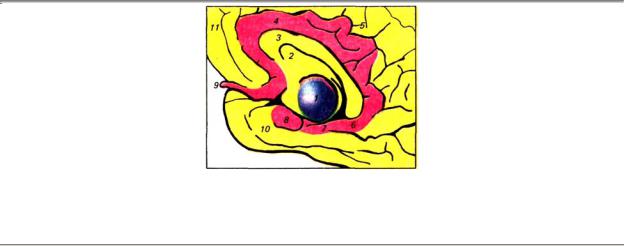
— старая кора (архикортекс), объединяющая сместившийся в процессе эволюции к височной доле гиппокамп (аммонов рог), зубчатую фасцию, основание гиппокампа (субикулум) и расположенную над мозолистым телом поясную извилину (рис. 3.14).
Рис. 3.14. Расположение отделов лимбической системы на внутренней поверхности полушария
(выделены красным): 1 — зрительный бугор (таламус), 2 — прозрачная перегородка, 3 — мозолистое тело, 4 — поясная извилина, 5 — поясная борозда, 6 — перешеек поясной извилины, 7 — парагиппокампальная извилина, 8 — крючок, 9 — обонятельная луковица, 10 — височная доля, 11 — лобная доля
Древняя и старая кора, которые обозначают как аллокортекс, граничат с пятислойной межуточной корой, или мезокортексом, переходящим непосредственно в новую шестислойную кору — неокортекс, или изокортекс. Мезокортекс формируется прилежащей к древней коре островковой, или инсулярной, зоной и граничащей со старой корой парагишюкамповой извилиной (энториальная область и предоснование гиппокампа, или пресубикулум), которые также включены в состав лимбической системы.
Из подкорковых структур в лимбическую систему входят расположенный в медиальной стенке височной доли миндалевидный комплекс и ядра мозговой перегородки.
Многие исследователи причисляют к лимбической системе также переднее таламическое ядро, мамиллярные тела и гипоталамус.
Все многочисленные формирования лимбической коры кольцеобразно охватывают основание переднего мозга и являются своеобразной границей между новой корой и стволовой частью мозга.
Лимбическая система характеризуется обилием двусторонних связей с другими отделами мозга и внутри самой системы. Так, например, установлено наличие мощных связей лимбической системы с гипоталамусом.
Через гипоталамус и мамиллярные тела лимбическая система соединена с центральным серым веществом и ретикулярной формацией среднего мозга.
К миндалине и гиппокампу идут пути от височной доли коры, передающие информацию от зрительной, слуховой и соматической сенсорных систем. Установлены связи лимбической системы с лобными долями коры переднего мозга. Наконец, в пределах лимбической системы идентифицированы сложные циклические связи, создающие условия для циркуляции возбуждения по сложным круговым путям. Примером такой циклической связи может служить так называемый круг Папеса, идущий от гиппокампа через свод — мамиллярное тело — переднее ядро таламуса — кору поясной извилины и пресубикулум обратно к гиппокампу.
Очевидно, сложность связей и внутренней организации лимбической системы свидетельствует об ее участии в интеграции функций новой коры и стволовых образований головного мозга.
3.8.2. Функции лимбической системы
Исследование функции лимбической системы у высших позвоночных началось сравнительно недавно. Длительное время в физиологии существовала точка зрения, что аллокортекс имеет только обонятельную функцию, и это нашло выражение в термине обонятельный мозг. Вместе с тем экспериментальные данные показывают, что помимо обонятельных афферентов в аллокортексе обнаруживаются проекции и других афферентных систем, конвергирующих к одним и тем же нервным структурам. Это и ряд других факторов заставили отказаться от традиционных взглядов и расширить наши представления о функциях древней и старой коры.
В 1937 г. американский невропатолог Д. В. Папес выдвинул гипотезу, согласно которой данные структуры мозга образуют единую систему (круг Папеса), ответственную за осуществление врожденных поведенческих актов и формирование эмоций. В 1952 г. другой американский исследователь, П. Д. Мак-Лин, развивая предположения Папеса, ввел понятие лимбическая система, представляя этим термином сложную функциональную систему, обеспечивающую постоянство внутренней среды и контроль видоспецифических реакций, направленных на сохранение вида. Теоретические и практические разработки Папеса и Мак-Лина послужили мощным толчком для дальнейших исследований в этой области.
Эти исследования показали, что локальное раздражение различных отделов лимбической системы вызывает разнообразные вегетативные эффекты и влияет на деятельность внутренних органов (см. разд. 5.5.4). Так, раздражение ядер миндалевидного комплекса приводит к изменениям частоты сердечного ритма, дыхательных движений, сосудистого тонуса. В ряде случаев раздражение миндалин влияет на деятельность пищеварительного тракта, изменяя перистальтику тонкого кишечника, стимулируя секрецию слюны, произвольное жевание и глотание. Описано влияние миндалин на сокращения мочевого пузыря, матки, пилоэрекцию и сокращение третьего века. Все эти разнообразные реакции могут иметь различный знак и характеризоваться активацией или угнетением висцеральных функций.
Существуют указания на то, что кортикальные и медиальные ядра миндалевидного
комплекса имеют отношение к регуляции пищевого поведения. Однако большей частью попытки связать тип реакции с определенным ядром миндалевидного комплекса не дали четких результатов. Факты указывают скорее на то, что функциональное представительство в миндалевидном комплексе, да и в других структурах лимбической системы (например, в гиппокампе) не дифференцировано. В некоторых случаях негативные результаты объясняются генерализованным, распространяющимся на соседние участки нервной ткани действием электрических стимулов.
Вместе с тем характер вегетативных реакций, вызванных раздражением различных отделов миндалины, идентичен эффектам при электрической стимуляции гипоталамуса, и
это позволяет думать, что регулирующие влияния лимбической системы опосредованы нижележащими вегетативными центрами.
Скорее всего, изменяя в ту или иную сторону возбудимость гипоталамических центров, лимбическая система определяет знак соответствующей вегетативной реакции. Так формируется многоэтажная, построенная по иерархическому принципу система управления вегетативной сферой, интегрирующая вегетативные и соматические реакции.
Как известно, эмоциональная окраска поведенческих реакций определяется не только вегетативными компонентами, но и соответствующими эндокринными сдвигами. В этом плане представляют интерес данные о влиянии лимбической системы на деятельность желез внутренней секреции. Установлено, что длительное (60 мин) раздражение ядер миндалевидного комплекса у обезьян вызывает повышение содержания кортикостероидов в плазме крови. Низкочастотное (12-36 имп/с) раздражение гиппокампа, напротив, уменьшает содержание кортикостероидов, которые выделяются надпочечниками при нанесении стрессорных раздражителей. Очевидно, нисходящие влияния этих структур лимбической системы на гипоталамус, а через него на гипофиз изменяют продукцию АКТГ, который регулирует секрецию кортикостероидов.
Таким образом, изменяя гормональный фон, лимбическая система в естественных условиях может участвовать в формировании побуждений к действию (мотиваций) и регулировать реализацию самих действий, направленных на устранение побуждения, усиливая или ослабляя эмоциональные факторы поведения.
3.8.3. Роль лимбической системы в формировании эмоций
Сведения об участии различных отделов лимбической системы в формировании эмоций неоднозначны. Наибольшее их количество относится к роли миндалевидного комплекса и поясной извилины в этих процессах. При локальном электрическом раздражении ядер миндалевидного комплекса могут быть получены эмоциональные реакции типа страха, гнева, ярости и агрессии. Двустороннее удаление височных долей вместе с миндалиной и гиппокампом вызывает у макак-резусов целый ряд сдвигов в эмоциональной сфере. Как правило, агрессивные обезьяны после этой операции становятся спокойными и доверчивыми. У животных наблюдается гиперорализм, когда все незнакомые предметы без разбора запихиваются в рот.
Удаление височных долей вызывает у обезьян гиперсексуальность, причем их половая активность может быть направлена даже на неодушевленные предметы. Наконец, послеоперационный синдром сопровождается так называемой психической слепотой. Животные утрачивают способность правильной оценки зрительной и слуховой информации, и эта информация никак не связывается с собственным эмоциональным настроем обезьян. Так, макаки без разбора исследуют все, даже опасные для себя предметы. Возникновение психической слепоты связывают с послеоперационным нарушением передачи сенсорной информации от височной доли к гипоталамусу. Эта точка зрения подкрепляется электрофизиологическими данными, свидетельствующими о том, что различные сенсорные раздражения изменяют частоту импульсной активности нейронов миндалевидного комплекса, который, очевидно, участвует в оценке
поступающей из внешней среды информации.
Согласно теории Папеса, сенсорные пути на уровне таламуса расходятся, причем один путь идет в проекционные зоны коры, где обеспечивается восприятие, а второй — в лимбическую систему. Вероятно, в этой системе происходит оценка поступающей информации, ее сопоставление с субъективным опытом и запуск соответствующих эмоциональных реакций через гипоталамические структуры. Информация об аффективном состоянии организма может поступать от стволовых структур в неокортекс либо прямо от миндалевидного комплекса, либо через поясную извилину, которая связана с лобными, теменными и височными долями коры полушарий большого мозга. Вероятно, эти пути связаны с восприятием эмоционального фона, однако детальные механизмы этого восприятия еще нуждаются в разработке.
Как уже отмечалось выше, лимбическая система принимает участие в запуске тех эмоциональных реакций, которые уже апробированы в ходе жизненного опыта. В этом плане привлекают внимание исследования, констатирующие участие лимбической системы в процессах сохранения памяти. Так, например, удаление гиппокампа вызывает у людей полное выпадение памяти на недавние события. Электрическое раздражение гиппокампальной извилины во время нейрохирургических операций может сопровождаться появлением мимолетных воспоминаний. Двустороннее удаление гиппокампа у обезьян и крыс приводит к нарушению способности выполнять ту или иную последовательность поведенческих актов. Описанные факты привели к заключению, что гиппокамп играет определенную роль в процессах памяти и послужили толчком для дальнейших исследований этой структуры.
Оказалось, что для строения гиппокампа характерно наличие четко отграниченных слоев с преимущественным расположением в них либо тел, либо отростков нервных клеток. Нейронные цепи гиппокампа представляют собой стереотипные микросети, которые состоят из возбуждающих волокон энториальной коры, клеток-зерен и пирамидных клеток с аксонами, идущими через свод к перегородке. Аксоны пирамидных клеток образуют коллатерали, направляющиеся к нейронам других частей гиппокампа.
Такая стереотипная слоистая структура гиппокампа делает его очень удобным объектом для изучения функции нейронных сетей на перфузируемых срезах мозга. Электрофизиологические исследования срезов и тотальных препаратов гиппокампа показали, что для этой структуры характерно периодическое возникновение низкочастотных (4—5 в 1 с) электрических колебаний тета-ритма. Эти колебания сопряжены с правильным чередованием возбуждающих и тормозных постсинаптических потенциалов пирамидных клеток гиппокампа и, вероятно, отражают синхронизацию деятельности его нейронных элементов.
Способность генерировать ритмическую активность, по всей видимости, зависит от упорядоченной слоистой структуры гиппокампа, которая создает условия для циркулирования возбуждения по нейронным цепям, лежащему в основе одного из нейронных механизмов памяти.
Об участии гиппокампа в процессах консолидации памяти свидетельствуют также изменения свойств его нейрональных синапсов после тетанической стимуляции. При высокочастотной стимуляции энториальной области коры кролика наблюдается длительное, достигающее нескольких часов, а иногда даже дней, возрастание амплитуды синаптических потенциалов клеток-зерен гиппокампа. В дальнейшем при микроэлектродном исследовании толстых срезов гиппокампа было установлено, что изменение эффективности синаптического проведения после тетанической стимуляции обусловлено повышением секреции медиатора из пресинаптического окончания. Параллельные электронно-микроскопические исследования показали, что после тетанической стимуляции наблюдалось увеличение числа шипиков на дендритах нейронов гиппокампа.
Эти факты убеждают в том, что пластичность нейронных цепей гиппокампа является
предпосылкой его участия в формировании нейронных механизмов памяти. Однако было бы упрощением считать гиппокамп единственным хранилищем следов памяти у позвоночных. Пластичность является весьма распространенным свойством нейронов, и поэтому большинство исследователей склоняются к тому, что функция памяти не является прерогативой какой-либо одной структуры, а обусловлена содружественными действиями многих центров головного мозга. Существенным звеном в этой системе являются связи гиппокампа с неокортексом.
Функциональная роль этих связей подтверждается физиологическими экспериментами. При одновременной регистрации электрической активности гиппокампа и неокортекса наблюдаются реципрокные взаимоотношения между ними. Когда в гиппокампе возникает медленноволновый тета-ритм, в неокортексе доминирует высокочастотная низкоамплитудная активность и, наоборот, медленноволновой активности энцефалограммы соответствует высокочастотная активность гиппокампа.
Наиболее выраженное усиление тета-ритма в гиппокампе обнаруживается на начальных стадиях выработки условного рефлекса и сочетается с состоянием настороженности и сосредоточения внимания при формировании ориентировочной реакции.
Удаление гиппокампа у животных нарушает процессы внутреннего торможения и снижает способность к угашению потерявших свое адаптивное значение условнорефлекторных реакций. Одновременно затрудняется упрочение условного рефлекса в связи с резким усилением ориентировочной реакции. Следовательно, гиппокамп, как, впрочем, и другие структуры лимбической системы, существенно влияет на функции неокортекса и на процессы научения. Это влияние осуществляется в первую очередь за счет создания эмоционального фона, который в значительной степени отражается на скорости образования любого условного рефлекса.
Таким образом, лимбическую систему как одно из наиболее древних образований мозга нельзя считать простым атавизмом. Это важный отдел головного мозга, функционально связанный с неокортексом и стволовыми структурами, образующими вместе систему координации висцеральных и соматических функций организма.
3.9. Базальные ганглии и их функции
Базальные ганглии, названные так гистологами прошлого века, представляют собой структуры ядерного типа, которые располагаются в толще белого вещества переднего мозга ближе к его основанию. У млекопитающих к базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро и заложенное в толще белого вещества чечевицеобразное ядро. Двумя белыми пластинками оно подразделяется на три части: наиболее крупную, лежащую латерально скорлупу и бледный шар, состоящий из внутреннего и внешнего отделов (рис. 3.15).

Рис. 3.15. Афферентные и эфферентные связи базальных ганглиев: 1 — хвостатое ядро, 2 — скорлупа, 3
— черная субстанция, 4 — таламус, 5, 5’— внутренний и внешний отделы бледного шара, 6 — субталамическое ядро, 7 — сенсомоторная кора, 8 — двигательная кора, 9 — пути к стволовой части мозга; синими стрелками обозначены афферентные пути, красными — эфферентные, черными — межъядерные взаимодействия
Эти анатомические образования формируют так называемую стриопаллидарную систему, которая по филогенетическим и функциональным критериям разделяется на древний палеостриатум и неостриатум. Палеостриатум представлен бледным шаром, а неостриатум, появляющийся впервые у рептилий, состоит из хвостатого ядра и скорлупы, которые объединяются под названием полосатого тела или стриатума. Хвостатое ядро и скорлупа связаны анатомически и характеризуются чередованием белого и серого вещества, что оправдывает возникновение термина полосатое тело.
К стриопаллидарной системе часто относят также субталамическое ядро (люисово тело) и черную субстанцию среднего мозга, которые образуют с базальными ганглиями функциональное единство. Полосатое тело состоит главным образом из мелких клеток, аксоны которых направляются к бледному шару и черной субстанции среднего мозга.
Полосатое тело является своеобразным коллектором афферентных входов, идущих к базальным ганглиям. Главными источниками этих входов служат новая кора (преимущественно сенсомоторная), неспецифические ядра таламуса и дофаминэргические пути от черной субстанции.
В противоположность полосатому телу бледный шар состоит из крупных нейронов и является сосредоточением выходных, эфферентных путей стриопаллидарной системы. Аксоны локализованных в бледном шаре нейронов подходят к различным ядрам промежуточного и среднего мозга, в том числе и к красному ядру, где начинается руброспиналышй тракт экстрапирамидной системы двигательной регуляции.
Другой важный эфферентный тракт идет от внутреннего отдела бледного шара к передневентральному и вентролатеральному ядрам таламуса, а оттуда продолжается к двигательным областям коры головного мозга. Наличие этого пути обусловливает многозвенную петлеобразную связь между сенсомоторными и двигательными областями коры, которая осуществляется через полосатое тело и бледный шар к таламусу. Примечательно, что в составе этого стриапаллидоталамокортикального пути базальные ганглии выполняют роль афферентного звена по отношению к моторным областям коры головного мозга. Многочисленные связи стриопаллидарной системы с различными отделами мозга свидетельствуют об ее участии в процессах интеграции, однако до настоящего времени в знании о функциях базальных ганглиев остается много невыясненного.
Базальные ганглии играют важную роль в регуляции движений и сенсомоторной координации. Известно, что при повреждении полосатого тела наблюдается атетоз — медленные червеобразные движения кистей и пальцев рук. Дегенерация клеток стриатума