

7. Вилка репликации днк: ферменты и их свойства.
Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В каждом сайте может формироваться одна или две репликационные вилки в зависимости от того, является ли репликация одно- или двунаправленной. Более распространена двунаправленная репликация.
В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500—5000 — у эукариот
DnA-белок. Он узнает участок начала репликации.
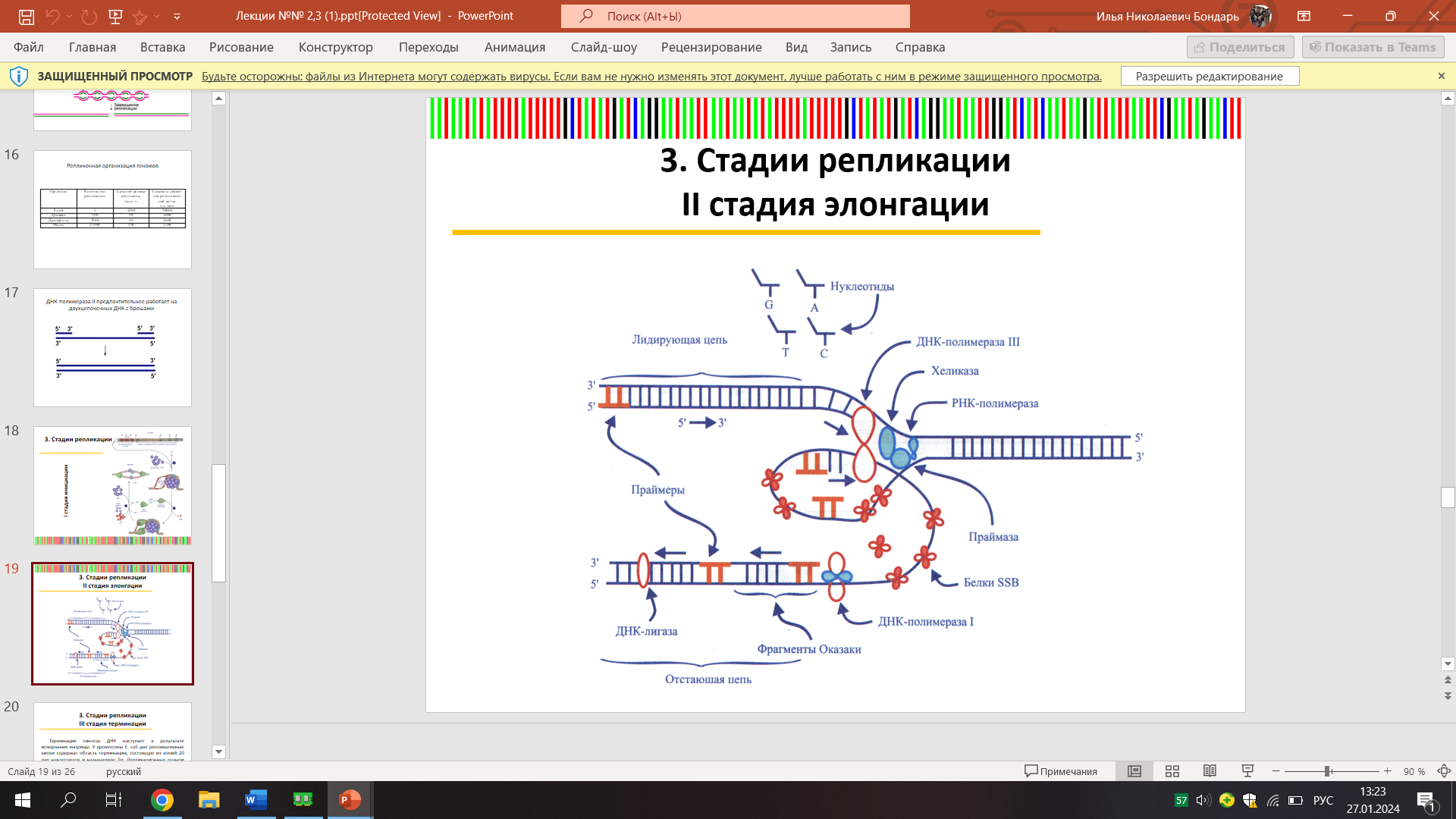
DnВ-белок (хеликаза). Данный фермент расплетает двойную спираль ДНК.
DnС-белок, необходимый для присоединения хеликазы к месту инициации синтеза ДНК.
ДНК-гираза (топоизомераза II) вводит отрицательные супервитки в ДНК и выполняет роль шарнира при продвижении репликационных вилок.
Праймаза (ДНК-зависимая РНК-полимераза или dnaG-белок), синтезирующая РНК-затравку (праймер).
ДНК-лигаза. Она соединяет концы фрагментов ДНК.
Dam метилаза обеспечивает метилирование последовательности (5)GАТЦ в oriC.
SSB-белки. Препятствуют образованию двойной спирали.
8. Стадии репликации.
Репликация ДНК начинается в определенном месте – в точке начала репликации. Репликация от точки начала репликации может происходить в одном или двух направлениях.
Инициация
Точка начала репликации -OriC
Для инициации нужно 3 составляющие:
Хеликаза (расплетает ДНК) образуется вилка
Топо-изомераза снимает напряжение при раскручивании, помогает раскручивать цепь
SSB-белки препятствуют обратному восставлению цепи
После подготовки матрицы образуется рипликационный пузырь, от него отходят 2 репликационные вилки.
Репликон-содержит точку начала репликации и способен к независимой репликации.
5’3’ - ведущая цепь
3’5’ - отстающая цепь
Инициация заканчивается синтезом праймера (катализирует ДНК-зависимая РНК-полимераза).
Элонгация
Ведущая цепь - непрерывный синтез
К 3’-ОН группе праймера присоединяется ДНК-полимераза 3, доноращивая цепь по принципу комплементарности в направлении от 5’3’.
Отстающая цепь
Фрагменты Оказаки осуществляют прерывистый синтез цепи в обратном направлении 3’5’ с помощью ДНК-полимеразы
Терминация
Удаление РНК-праймеров экзонуклеазой. ДНК-полимераза достраивает недостающие фрагменты. Лигаза связывает фрагменты между собой.
9. Механизм репликации у e.Coli.
В клетках E.coli присутствуют три ДНК-полимеразы.
ДНК-полимераза I
На одной полипептидной цепи ДНК-полимеразы I находятся 2 активных центра:
a) 1-й активный центр ответственен за полимеразную и 3’→5’ экзонуклеазную активности. Последняя обеспечивает удаление ошибочно встроенных нуклеотидов;
b) 2-й активный центр ответственен за 5’→ 3’-экзонуклеазную активность. Эта активность необходима для удаления РНКзатравки в процессе репликации.
ДНК-полимераза II
Этот фермент обладает полимеразной и 3’→ 5’-экзонуклеазной активностями, предпочтительнее работает на двухцепочечных ДНК с брешами. ДНК-полимераза II участвует в репарации ДНК.
ДНК-полимераза III
Этот фермент обладает полимеразной и 3’→5’-экзонуклеазной активностями, состоит из десяти типов субъединиц. Его основное назначение – репликация ДНК. Скорость синтеза – 500 нуклеотидов в секунду
Хромосома Е.coli имеет одну точку начала репликации (oriC), ее – размер 258 н.п. Белок Dna А узнает OriC и инициацирует репликацию. В результате АТФ-зависимая хеликаза начинает расплетать дуплекс ДНК. Топоизомераза, располагаясь впереди по ходу движения репликативной вилки, снимает напряжение, возникаюшее в результате расплетения двойной спирали в ДНК.
С образующимися одноцепочечными участками ДНК связывается SSB-белок. Праймаза осуществляет синтез затравки, которая необходима для проявления активности ДНКполимеразы. Затем в работу включается ДНК-полимераза III, которая последовательно присоединяет нуклеотиды к 3’-концу полинуклетидной цепи. Поскольку синтез ДНК осуществляется в направлении 5’→ 3’, одна цепь (ведущая) синтезируется непрерывно, вторая фрагментами (отстающая цепь) по 1000 – 2000 нуклеотидов (фрагменты Оказаки).
По окончанию синтеза фрагмента Оказаки ДНК-полимераза I за счет 5’→ 3’-экзонуклеазной активности удаляет затравку и заменяет ее ДНК. После действия этого фермента между фрагментами Оказаки остается разрыв, который сшивает ДНК-лигаза.
Терминация репликации происходит после удвоения кольцевой молекулы ДНК.