17 Заказ 552
ные воды. Выносливость их, по-видимому, обусловливается внутренними физиологическими перестройками в^ клетках, структурой и химическим составом цитоплазмы.
Водоросли снега и льда
Другая группировка, приспособленная к жизни в крайних температурных условиях (при 0°С и ниже), — криофитон, или криофильны е водоросли. Они развиваются на поверхности снега и льда и в период интенсивного размножения нередко придают субстрату ту или иную окраску.
Наиболее известно явление так называемого «красного снега». Чаще всего оно вызывается СЫатуйотопаз пшаНз, в клетках которой содержится большое количество красного пигмента астаксантина. Другой вид СН. ^^аVО-V^^епз, встречающийся в Высоких Татрах, окрашивает снег в желто-зеленый цвет. Позеленение снега, отмеченное в Гренландии, в Альпах и ряде других высокогорий, вызывалось кроме видов СЫатуйотопаз видами РирЫйопета и некоторыми десмидие- выми. Вместе с ними присутствовали также синезеленые водоросли, С1епо(Итит разсНеп из БторЬусеае и др. Буровато-желтую окраску обычно придают снегу диатомовые.
«Снежные» водоросли развиваются, как правило, не на свежевы- павшем, а на старом снегу, оставшемся лежать в холодных ущельях или на снежных полях высоко в горах. Рост и размножение водорослей начинается после того, как под действием солнечной радиации снег частично растает. Площадь окрашенного снега может занимать до нескольких квадратных километров.
В настоящее время список «снежных» водорослей насчитывает свыше 100 названий. Среди них особенно распространены зеленые, диатомовые и синезеленые водоросли. Ограниченное число видов имеют золотистые и пирофитовые водоросли. Окраска снега в зависимости от преобладания в нем того или иного вида (а также стадии развития) может быть красной, зеленой, желтой, бурой, почти черной.
Во льдах арктических и антарктических бассейнов сильно разрастаются диатомеи. Первоначально они развиваются на нижней, погруженной в воду поверхности льда, а затем в толще его, распространяясь по тонким канальцам, которые пронизывают лед в разных направлениях. В случае массового развития диатомовые водоросли вызывают интенсивное буровато-желтое окрашивание льда на больших пространствах. При этом продукция диатомей весьма значительна: с 1 м3 льда собирают около 1 кг (сырой массы) водорослей.
Водоросли соленых водоемов
Своеобразную группировку организмов представляют собой водоросли соленых озер Концентрация солей в континентальных водоемах может быть очень высокой: она доходит до 285 г на .1 л воды, если в ней преобладает хлористый натрий, и до 347 г на 1 л при преобладании сульфата натрия. Солевыносливых водорослей довольно много, известны даже виды, живущие в насыщенных растворах. Альгофлора засоленных водоемов не может считаться морской, так как типичные обитатели моря здесь отсутствуют. По происхождению это, скорее, пресноводные формы, приспособившиеся к повышенной концентрации солей.
Организмы, населяющие воды с хлористыми солями, делятся на две группы: эвгалофиты и олигогалофиты. К первой относятся формы, живущие только в присутствии солей натрия и магния (из них полигалобионты населяют воды с высокой их концентрацией, близкой к морской, а иногда и большей, а мезогалобионты — с уменьшенным содержанием солей, например во внутренних морях и в опресненных морских бухтах). Вторая группа включает формы, которые живут в воде либо без хлоридов (галофобы), либо с незначительным их содержанием, не более 5 %о.
Типичный полигалобионт — ОипаНеИа заИпа — одна из наиболее распространенных водорослей соленых водоемов, сходная по строению с хламидомонадой." В клетках этого вида дуналиеллы содержится красный пигмент р-каротин. В периоды массового развития пигмент из отмерших клеток водорослей переходит в солевой раствор (рапу), окрашивая его и соль, выпадающую в виде кристаллов, в разные оттенки красного цвета
Другой обычный обитатель соленых озер — синезеленая водоросль СН1ого§1оеа загстоШез, встречающаяся в СССР на юге, вблизи Евпатории. Мощные слизистые скопления этой водоросли образуют пласты в береговой части некоторых озер. Йл, остающийся после отмирания хлороглеи, используется в лечебных целях.
Известковые водоросли
К этой группе относятся интересные «сверлящие» водоросли, всегда связанные в своей жизни с известковым субстратом. Эти микро- с» тес кие формы особенно распространены среди синезеленых, но имеются и в других группах. Число их видов небольшое, около 20, встречаются же они довольно широко в пресных и морских водах, на вз? ;тковых скалах, раковинах, кораллах. Выделяя органические кис- д" . водоросли постепенно растворяют известь, в результате чего субстрат оказывается пронизанным тонкими и глубокими канальца- мл — ходами.
Противоположную группу известковых водорослей образуют фор- жы. способные выделять углекислый кальций1. Количество извести, об- -»зуемое разными видами, неодинаково: у одних оно небольшое, у дру- - настолько обильное, что клетки и нити водорослей как бы заключаются в своеобразные футляры, у третьих организм полностью погружается в субстрат, в котором он постепенно отмирает. Жизнеспособными в таких случаях остаются лишь водоросли, расположенные ближе к поверхности субстрата.
Сожительство водорослей с другими организмами
Водоросли не всегда ведут свободный образ жизни. Очень часто можно наблюдать сожительство — симбиоз их с другими организмами: бактериями, грибами, другими водорослями, высшими растениями и животными. Взаимоотношения между организмами, составляющими такие сочетания или комплексы, степень влияния одного компонента на другой могут быть самыми разными и не всегда носят характер положительных взаимодействий (когда один из видов извлекает для себя пользу из сожительства с другим видом и не причиняет ему никакого вреда).
Обычно различают несколько типов сожительства водорослей с другими организмами: эпифитизм, эндофитизм, паразитизм, мутуализм. Эпифитизм представляет такой тип сожительства, при котором имеет место тесный наружный контакт между водорослью и другим организмом, на котором она поселяется, при автономности их питания. Эндофитизму при сохранении самостоятельности питания взаимосуществующих организмов свойствен уже внутренний контакт между ними (водоросль живет внутри другого организма, не нанося ему видимого вреда). Паразитизмом называют такие взаимоотношения, когда внедрившаяся в тело хозяина водоросль питается за его счет и в конечном итоге приводит к отмиранию отдельных частей организма хозяина или иногда к полной его гибели. Мутуализм, наоборот, представляет собой такой тип взаимоотношений, когда совместно существующие виды (один организм внутри другого) извлекают из своего сожительства множество преимуществ и иногда не в состоянии жить раздельно. В природе такие случаи встречаются, правда, очень редко. Следует подчеркнуть, что резко разграничить перечисленные типы взаимоотношений, обнаружить их в чистом виде на практике удается далеко не всегда, скорее можно наблюдать переходные формы.
Эпифитизм среди водорослей — очень частое явление. Он заключается в том, что водоросли-эпибионты используют другие растения как место поселения и, не проникая внутрь их тела, питаются вполне самостоятельно. При этом один и тот же вид водоросли может прикрепляться не только к разным растениям, но даже к неживым субстратам Ч В пресных водоемах эпифитами, например, являются диатомеи, поселяющиеся на талломе одной из распространенных зеленых водорослей — кладофоры, а в морях — многие бурые и красные водоросли, прикрепляющиеся к многолетним черешкам ламинарий, основаниям фукусов и других крупных форм. Все отделы водорослей имеют представителей, ведущих эпифитиый образ жизни1. Приспособления к прикреплению у эпифитов разной систематической принадлежности часто носят конвергентный характер. Пример тому — удивительно похожие морфологически, но относящиеся к разным отделам коккоидные водоросли. Отношения между эпибионтом и хозяином не всегда складываются безразлично для организма, служащего опорой. Это особенно выявляется при массовом развитии эпифитов, когда они почти сплошным слоем обрастают растение. Та же кладофора или аквариумные высшие растения, плотно покрытые талломами эпифитных водорослей, испытывают явное угнетение. Причиной, по-видимому, служит избыточное затенение, нарушающее фотосинтез этих растений.
Более специализированным и сложным типом сожительства двух организмов оказывается эндофитизм (эндосимбиоз), когда водоросль живет полностью или частично внутри другого организма. При этом хозяин не обнаруживает признаков какого-нибудь подавления, а эндофит, используя его тело как жизненное пространство и сохраняя способность самостоятельно питаться, не теряет своей независимости. Эндофит поселяется либо в толще клеточной стенки другой водоросли, либо проникает между клетками, раздвигая их. К первому типу_ относится Со1еосНае(е пИеИагит, таллом которой помещается в стенках междоузлий нителлы, и Епйойегта, поселяющаяся в стенках кладофоры. Ко второму типу принадлежит описанный уже выше хло- рохитриум, виды АпаЬаепа и N08(00, живущие в воздушных камерах мхов Ап(Носего8 и В1ав1а, АпаЬаепа агоЫае, непременно присутствующая в полостях на (нижней стороне листьев водного папоротника Аго1- 1а атепсапа, а также N05(00 рипсЩогте, обитающий в межклетниках С у саз и 2,апйа.
С эндофитизмом тесными переходами связан паразитизм. В качестве примеров таких переходов можно взять близкие к хлорохигриуму роды РНуИоЫит и ННойосНу(пшп. Филлобиум поселяется вблизи сосудистых пучков некоторых высших растений (из разных семейств) и проявляет определенные паразитические тенденции. Своеобразные мешковидные выросты, образуемые им на теле хозяина, могут быть истолкованы как приспособления для восприятия питательных материалов из окружающих тканей. Родохитриум, встречающийся на сложноцветных,— уже настоящий паразит, живущий исключительно за счет хозяина, так как из-за отсутствия хлорофилла утерял способность к автотрофному питанию. Другой пример — Тгеп(ероНИа и СерНа1еигоз (рис. 182, А). Трентеполия рассматривается одновременно и как эпифит, и как эндофит, поскольку глубоко проникает внутрь пробковой ткани коры деревьев. Цефалеурос выступает как промежуточная форма между эндофитами и паразитами. Разрастаясь внутри живых листьев тропических растений, он вызывает отмирание прилегающих к нему тканей растения-хозяина. При этом питание цефалеуроса не исключительно паразитное, так кате он сохраняет хлорофилл.
Б
«Рис. 182. А — СерНакигов влисте высшего растения; Б — ИагоеуеИа пМгаЬШв на %ко- л с1оте1а(мелкоклеточные нити паразита внедряются в ткань хозяина)
Из красных водорослей наиболее совершенный пример паразитизма представляет лишенная хлорофилла НагьеуеИа тьгаЬШз (рис. 182, В), развивающаяся на Я.Нойоте1а (из красных же водорослей).
В отличие от вышеперечисленных примеров паразитизма эндосим- биозы могут характеризоваться и взаимовыгодными отношениями партнеров, наиболее тесно и длительно связанных друг с другом в едином комплексе. Классическим примером межклеточного эндосимбиоза служат лишайники, представляющие собой настолько прочное и гармоничное биологическое единство, что образуется целостный организм нового типа. В такой кооперации гриб защищает водоросль, а водоросль его кормит, так как сохраняет хлорофилл и способность к фотосинтезу. Правда, не всегда взаимоотношения между грибом и водорослью в лишайнике столь просты и безобидны, особенно для водоросли. Большая часть выгод от такого сожительства находится все-таки на стороне гриба, доминирующего в лишайниковых талломах.
Внутриклеточный эндосимбиоз распространен значительно шире.
„■ В качестве симбионтов водоросли могут соединяться с животными — одноклеточными и многоклеточными (инфузориями, радиоляриями, гидрами, губками, некоторыми червями и др.). Сами они принадлежат чаще всего к простейшим зеленым и пирофитовым водорослям и носят специальное название зоохлорелл (2оосН1оге11а) и зооксантелл (%оо- хап1Не11а).
л
Рис.
183. Внутриклеточный симбиоз.
А — С1аисо- еу&Ш; Б — Сео&рНоп
я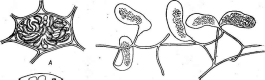

верительного "тракта клетки Еи§1епа цгасШз, ресничный червь конволют (СопюоШа гозсоЦепзьз) с картерией (Сайепа) и др.
Следующий этап развития симбиотических отношений характеризуется еще более прочными и, главное, продолжительными связями между двумя организмами, Автотрофный компонент в таких комплексах содержится постоянно и при размножении животных передается из поколения в поколение, сохраняясь как вид. В подобных вариантах водоросль приспосабливается к сожительству только с определенными видами животных. Пример такого рода симбиозов — зеленая гидра, обитающая в пресных водоемах. В ней автотроф находит подходящие условия существования, о чем свидетельствует его интенсивное размножение.
Особенно интересны случаи внутриклеточного сожительства у синезеленых „ водорослей. Они вступают с простейшими и некоторыми другими организмами в своеобразные симбиотические отношения, обозначенные А. Пашером специальным термином синцианоз, при этом комплекс из двух организмов называют цйаномом, а синезеленые водоросли в нем — цианеллами.
Наглядными примерами могут быть СуапорНога ратайоха и 01аи- сосузИз поз1осЫпеагит (рис. 183, А). Первый организм был описан как новый вид водоросли в 1924 г., а второй известен с конца прошлого века как одноклеточная форма неопределенного систематического положения. Позднее при детальном исследовании этих организмов обнаружили, что они представляют собой симбиозы: в первом случае—> одноклеточной водоросли криптомонады и цианеллы из рода СНгоо- соссиз, во втором—одноклеточной обесцветившейся формы (близкой к роду ОосузИз) и палочковидной синезеленой водоросли, которую не удается точно идентифицировать. Окрашенные в гсиний цвет внутриклеточные включения, описанные ранее как «хроматофоры», оказались, таким образом, симбиотическими синезелеными водорослями.
Электронно-микроскопическое изучение цианелл (на примере глаукоцистиса) показало, что по своей тонкой организации они мало отличаются от свободноживущих синезеленых водорослей. Однако своеобразие условий жизни внутри другого организма не могло совсем не отразиться на строении симбионтов. Изменения эти касаются клеточных стенок (у внутриклеточного симбионта глаукоцистиса типичных клеточных стенок нет, клетка окружена лишь тонкой мембраной — плаз- малеммой) и запасных продуктов (цианеллы их не накапливают, так как необходимое питание получают из клетки хозяина). Относительно функционирования цианелл в составе комплексного организма следует сказать, что они берут на себя роль хлоропластов — «поставщиков» органического вещества, крахмала, откладывающегося, правда, не в них, а в цитоплазме бесцветных клеток хозяина.
Во всем остальном они ведут себя как вполне самостоятельные, автономные организмы. Делятся цианеллы перетяжкой пополам, причем время их деления не связано с периодом размножения хозяина. При делении в каждую автоспору водоросли попадает некоторое количество цианелл, что обеспечивает непрерывность симбиоза. При изоляции водорослей, составляющих цианом, они довольно легко растут и размножаются на искусственных средах.
Аналогичные симбионты имеются у ОеозьрНоп и Я.Ыго8о1ета. Разница с предыдущими примерами в том, что здесь синезеленые водоросли свой облик заметно не меняют и их легко узнать. Это нитчатые формы, близкие к Ыоз(ос и АпаЬаепа. Геосифон встречается на влажной почве и внешне напоминает маленьких ботридиумов (рис. 183, Б). Таллом его состоит из стелющихся разветвленных нитей (без перегородок) и отходящих от них вверх пузыревидных вздутий, содержащих нити ностока. В таком цианоме, как и в вышеописанных, отсутствие хроматофоров в растении-хозяине компенсируется за счет фотосинтеза синезеленой водоросли. Некоторые виды ризосолении, содержащие в цитоплазме нити анабены, в отличие от геосифона не утрачивают собственных хроматофоров
ЗНАЧЕНИЕ ВОДОРОСЛЕЙ В ПРИРОДЕ И ЖИЗНИ ЧЕЛОВЕКА
Благодаря широкому распространению водоросли имеют большое значение в жизни отдельных биогеоценозов, в круговороте веществ в природе, а также в хозяйственной деятельности человека.
Составляя основную часть растительности водной среды, эти фотосинтетики являются одними из главных поставщиков органического вещества в водоемах. Они представляют собой начальное звено в цепи питания различных гидробионтов, включая и наиболее важных из них— рыб. При этом роль планктонных форм, называемых «микроскопическими лабораториями органического вещества», просто незаменима. Питательная ценность планктона по содержанию белков и жиров не уступает ценности многих пищевых растений..
Обилие в водоемах мельчайших растительных организмов в первую очередь определяет количество его животного населения. Учитывая это обстоятельство, для искусственного рыборазведение выбирают водоемы, богатые фитопланктоном. В случае же недостаточного его развития, например в олиготрофных прудах, применяют органические и минеральные удобрения. При этом продукция планктона обычно сильно возрастает.
В морях, где" имеется такое же соотношение между растительным и животным населением, по степени развития фитопланктона нередко ставят прогнозы для рыбного промысла.
Однако чрезмерное изобилие микроскопических растений в водоемах отрицательно сказывается на качестве воды, приобретающей неприятный привкус и запах. Накопление же в йей прижизненных Выделений водорослей и ядовитых веществ, образующихся при распаде их клеток, губительно действует на- животные организмы. Причиной появления токсичных вод могут быть многие водоросли, вызывающие «цветение» водоемов. Наиболее часты из них виды Аркатготепоп, М'югосуШв (синезеленые водоросли), Ргутпезшт ратит (золотистые) и т. д. ' .
При массовом развитии водорослей-обрастателей возникают 'трудности в водоснабжении, в эксплуатации водного транспорта и различных гидротехнических сооружений.
С другой стороны, водоросли (вольвоксовые, эвгленовые, желто- зеленые, пирофитовые, диатомеи) вместе с гетеротрофными организмами как активные санитары осуществляют процессы естественного самоочищения сточных и загрязненных вод. Многие виды их являются индикаторами биологического загрязнения и засоления.
Наряду с образованием биомассы постоянно идет обратный процесс— отмирание водорослей. В водоемах оседающий планктон входит в состав детрита, который служит пищей для осмотрофов: бактерий, грибов, актиномицетов, окончательно разрушающих мертвое органическое вещество. В случае развития, например, диатомового планктона детрит идет не только на питание организмов; ему принадлежит также ведущая роль в осадконакоплениях. Диатомовые, зеленые, золотистые водоросли формируют илы, сапропели и некоторые породы. Так, диатомит на 50—80 % состоит из панцирей диатомей, некоторые известняки сложены в основном из остатков золотистых и харовых водорослей.
Водоросли, живущие в почвах, повышают их плодородие. Особенно ценны в этом отношении азотфиксирукяцие синезеленые. Те же растения, которые осваивают свежеобнаженные субстраты, скалы, песчаники, участвуют в создании первичного гумуса, в результате чего такие места становятся пригодными для жизни других организмов. На бесплодных участках земной поверхности, как правило, первыми селятся осцилляториевые из синезеленых водорослей. Вместе с бактериями, грибами и лишайниками им принадлежит роль пионеров растительности.
При определении пригодности свежеобнаженных грунтов промышленных отвалов для последующей биологической рекультивации водоросли используются в качестве биоиндикаторов. Они служат удобными тест-объектами и в случае изучения токсичности почв после внесения гербицидов или попадания других ядов.
Что касается прямого использования водорослей для нужд человека, то на первое место следует поставить красные (Рогркуга, ННойу- •тета и др.) и бурые (Ьаттапа, А1аг'ш, 1)пйапа и др.). Морские макрофиты с древних времен используют в пищу и как корм для -скота. Приготовление салатов, супов, острых приправ и кондитерских изделий из этих водорослей особенно популярно на Востоке (Японии м Китае). В этих странах в широких масштабах занимаются искусственным выращиванием морских макрофитов. В приморских районах большинства европейских государств водоросли обычно употребляют для кормления домашних животных.
Водоросли, используемые в пищевых целях, не только питательны, но богаты витаминами и солями иода, брома. В связи с этим добавление их в пищу или применение в виде лекарственных препаратов рекомендуется с целью лечения и профилактики ряда заболеваний. Например, морская капуста назначается при склерозе, расстройстве деятельности щитовидной железы и как слабое слабительное средство.
В гинекологической практике- и при лечении ран успешно применяют специально приготовленные тампоны из черешков ламинарии (Ьаттапа <Н@На1а и Ь. с1ои81оп1). В этом случае используется свойство сухих водорослей сильно набухать при увлажнении.
Из пресноводных водорослей в пищу идут крупные колонии носто- ков, а также ЗрииНпа рШепз'18, издавна употребляемая некоторыми африканскими народностями. В настоящее время в ряде стран налажено массовое культивирование этой водоросли.
Морские водоросли служат сырьем для многих отраслей промышленности. Наиболее важные продукты, получаемые при переработке красных и бурых макрофитов,— агар-агар, или кантен, и альгин. Агар образуется при вываривании талломов красных водорослей и состоит из смеси различных полисахаридов. При комнатной температуре расплавленный агар легко застывает и превращается в плотный студень наподобие животного желатина. Агар выпускается промышленностью в виде тонко размолотого порошка, лент или плиток. Он чрезвычайно широко применяется в пищевой, бумажной, фармацевтической, текстильной и других отраслях. Совершенно незаменим агар в микробиологической практике при культивировании микроорганизмов. В СССР получают агар из анфельции, добываемой в Белом и дальневосточных морях, и агароид (агар низкого качества) из филлофоры, большие запасы которой имеются в Черном море.
Альгин и альгинаты, извлекаемые из бурых водорослей, обладают превосходными клеящими свойствами. Эти вещества добавляют в пищевые продукты, в таблетки при изготовлении лекарственных препаратов, используют при выделке кож, при производстве бумаги и тканей, синтетических волокон, пластмасс, строительных материалов. Иэ этих соединений делают растворимые нити, используемые в хирургии.
В настоящее время из морских водорослей промышленным путем получают зольные элементы: калий и натрий (соли их используют для удобрений), а также некоторые органические кислоты, маннит, ацетон и ряд других веществ.
В последнее десятилетие большое внимание уделяется вопросам промышленного применения микроскопических водорослей. Особенно важны работы по освоению массовых культур этих растений в условиях искусственного выращивания либо под открытым небом, либо в замкнутых аппаратах. Биомасса микроскопических водорослей используется как дополнительный источник белка и витаминов, для регенерации воздуха в замкнутых экологических системах, например в космических кораблях, а также для получения ценных для промышленности веществ.
Культуры водорослей необходимы и в научно-исследовательской работе. Значение водорослей как объектов исследования в области физиологии, биохимии, генетики, биофизики и общей биологии трудно переоценить. Можно без преувеличения сказать, что многие открытия в биологии в последние десятилетия связаны с изучением именно низших автотрофных растений. Широкую известность среди них получили микроскопические хлорококковые водоросли (СМогеИа, Зсепейевтиз), а также водоросли с крупными талломами, очень удобными для экспериментирования: харовые (СНага, ЫНе11а) и бриопсидовые (Асе1аЬи- 1апа).
ОТДЕЛ СЛИЗЕВИКИ — МУХОМУСОТА
Отдел объединяет свыше 450 видов бесхлорофилльных организмов, довольно разнообразных по строению и образу жизни. Среди них есть как свободноживущие в почве, на навозе, на разлагающихся растительных остатках и тому подобных субстратах, так и паразиты водорослей, водных грибов и высших водных и наземных растений.
Некоторые из них имеют микроскопически малые размеры и представлены одноядерной или многоядерной амебоидной клеткой, у других вегетативное тело а виде крупного многоядерного цитоплазменно- го образования, так называемого плазмодия, достигающего в отдельных случаях даже нескольких десятков сантиметров. У некоторых форм, относимых в этот отдел, развивается не настоящий плазмодий, а псевдоплазмодий, т. е. скопление амеб, не теряющих своей индивидуальности, но ведущих себя как единое целое.
В вегетативной стадии большинство слизевиков, представленных амебоидными клетками, размножается делением этих клеток. Для некоторых слизевиков с плазмодиями показано также, что в течение вегетативной жизни от плазмодиев могут отделяться небольшие участки, далее существующие в виде двужгутиковых зооспор.
В жизненном цикле разных групп слизевиков имеются некоторые особенности. Так, у слизевиков с вегетативными телами в виде сво- бодноживущих плазмодиев в определенный момент развития, зависящий от состояния плазмодия и от условий среды, формируются плодовые тела, содержащие споры, одетые твердой оболочкой. У паразитов таких плодовых тел не образуется и плазмодий распадается на споры в клетках хозяина, служащих им вместилищем. /
Зрелые споры в благоприятных условиях прорастают, т. е. из нх оболочки выходят (в зависимости от степени влажности субстрата) или зооспоры с двумя гладкими жгутиками неравной длины, или амебы. Те и другие могут размножаться делением, увеличиваясь в количестве, а затем клетки попарно сливаются. Возникает диплоидная клетка, развивающаяся затем в плазмодий. У слизевиков с псевдоплазмодиями полового процесса нет, а псевдоплазмодий образуется в результате агрегации амеб.
Отдел включает следующие классы, различающиеся по степени организации и особенностям циклов развития своих представител. протостелиевые (Рго1о51еНотусе1е5), миксогастровые, или собственно слизевики (Мухоеа51егошусе1ез), плазмодиофоровые (Р1азтос1юрЬого- шусе1е5), акразиевые, или клеточные слизевики (Асгазютусе1е8).
КЛАСС ПРОТОСТЕЛИЕВЫЕ — РКОТОЗТЕЬЮМУСЕТЕЗ
Большинство представителей класса обнаружено и описано только в середине нашего столетия, хотя и представляет собой широко распространенные организмы. Они могут быть выделены из отмершей
Рис.
184. Схема цикла развития протостелид: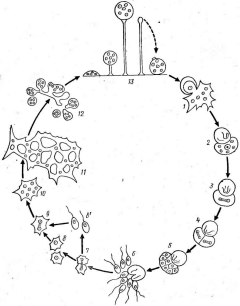
/ — прорастающая спора, 2— образование цисты, 3—I — два деления единственного сохранившегося ядра, 5 — третий мнтоз, 6— образование зооспор, 7—9— деление клеток с образованием зооспор (6') или амебоидов (8)2 10—И— образование из амебоида сетчатого плазмодия, 12— образование
предспоровых клеток, 13— спорогенеэ
Рис.
185.А — СегаИо- туха
{гиИси1о&а~угх. [1е- я хио&а:
А1— внешний вид спороно- шения,
А2— спорангии
иа
иожках,
сидящие на рожковидном выросте;
Б
—С.
1гиИси1овауаг.ропоЫев,внешний вид , спороношения

древесины, коры деревьев, из почвы, особенно богатой гумусом, из навоза, из покровов отмерших, нр не отделившихся плодов (коробочек, стручков, ягод и т, п.).
В настоящее время известно около десятка родов и немногим более видов этого класса. Большинство представителей относительно просты по морфологии и характеризуются микроскопически малыми размерами вегетативных тел — амебоидов. Лишь у немногих вегетативное тело представляет собой многоядерный сетчатый плазмодий.
Из амеб (или частей плазмодия у некоторых протостелиевых) образуются плодовые тела — спорокарпы непосредственно на субстрате, а у других — на рожкообразных выростах. Они представляют собой сидящую на тонкой полой ножке головку .с одной, двумя или большим числом спор. Зрелая головка отделяется от ножки, и в подходящих условиях споры освобождаются и прорастают четырьмя — восемью одноядерными подвижными клетками — зооспорами, превращающимися в амебоиды, или непосредственно амебоидами, с которых начинается новый цикл. У видов, имеющих плазмодий, последний развивается из разрастающихся амебоидных клеток (рис. 184).
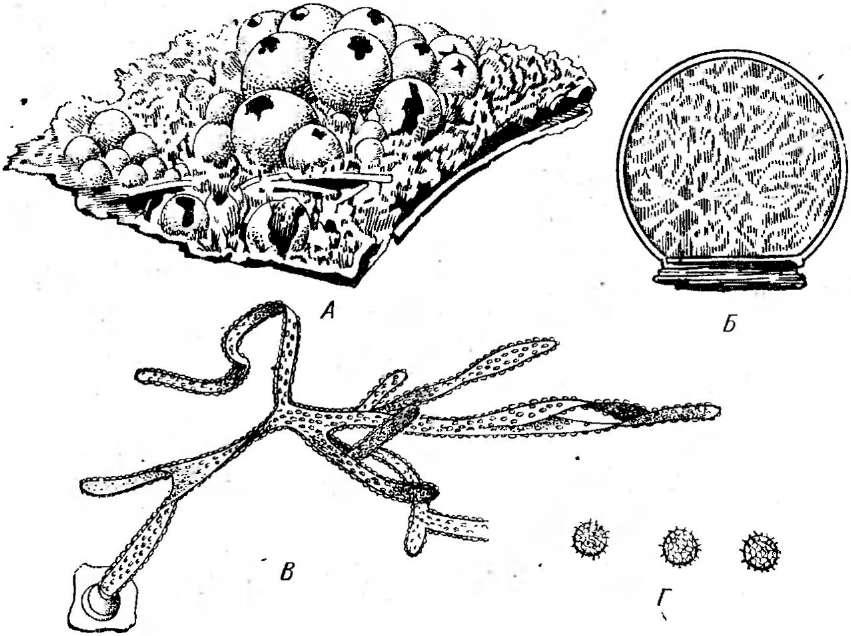
Наиболее широко известный представитель класса — церациомикса кустарничковая (СегаЫотуха 1гиИси1оза), отличающаяся макроскопическими размерами и высокой степенью морфологического развития. В СССР встречается в двух формах. Плазмодий и спороношения этого слизевика летом можно найти повсеместно, чаще на гнилой древесине, иногда на опавших листьях или на лесной подстилке. Плазмодий имеет вид прозрачной стекловидной, почти белой или желтоватой слизистой массы, иногда с оттенками розового, абрикосового или зеленоватого цвета. Он преобразуется или в пучки поднимающихся вверх и аиа- стомозирующих выростов у одной из форм, или в более компактные образования, напоминающие пчелиные соты у другой. По всей поверхности выростов или йчеек «сот» на тонких полых ножках сидят одно- споровые бесцветные спорангии (рис. 185).
По-видИмому, протостелиевые в своем происхождении связаны со свободноживущими жгутиковыми (Р1а^е11а1ае), на что указывает существование жгутиковой стадии у некоторых представителей. У бесцветных и окрашенных жгутиковых также наблюдается образование плазмодиев и оно, безусловно, облегчало выход на сушу древним жгутиковым. Развитие спороношений было дальнейшим шагом приспособления к наземной жизни.
КЛАСС МИКСОГАСТРОВЫЕ,
ИЛИ СОБСТВЕННО СЛИЗЕВИКИ,—
МУХОСА8ТЕРОМУСЕТЕ5
Центральный и самый крупный класс отдела, насчитывающий более 400 видов, обитающих главным образом в лесах в глубине гнилых пней, под корой разлагающихся колод, под опавшими перегнивающими листьями и тому подобных влажных и темных местах, богатых органическими веществами.
Вегетативное тело этих слизевиков представляет собой плазменную массу с большим количеством диплоидных ядер — плазмодий. Размеры плазмодия у разных видов могут быть от нескольких миллиметров (или даже долей миллиметра) до 15—20 см и более. В состав плазмодия входит вода, до 30 % белков, гликоген, масло, пигменты различной природы, окрашивающие плазмодии многих слизевиков в яркие цвета, ферменты, витамины, стеролы и другие органические вещества; некоторые содержат до 30 % извести. Плазмодий поглощает из субстрата растворенные, в том числе органические вещества, активно захватывает амеб, бактерий, мелких беспозвоночных животных, споры грибов, различные твердые пищевые частицы, перемещаясь в направлении к источникам пищи и току воды (положительные трофо-, гидротаксисы и др.1). Он быстро увеличивается в размерах и в определенный момент развития выползает на свет: на поверхность пней, листьев и т. п., т. е. отрицательный фототаксис сменяется положительным. Здесь плазмодий преобразуется в спороношения, очень разнообразные у слизевиков.
Самый простой тип спороношения представляет собой так называемый плазмодиокарп, развивающийся таким образом, что плазмодий, не меняя формы, покрывается перепончатой или хрящеватой' оболочкой (перидием), а находящаяся внутри цитоплазма распадается на споры, одетые твердой оболочкой. У некоторых слизевиков известен только такой тип спороношения, у большинства же (иногда наряду с ним) формируются многочисленные одиночные спорокар- п ы, одетые перидием, сидячие или на ножках. У некоторых слизевиков такие спороношения (спорокарпы) сливаются на ранних стадиях развития, одеваясь общей оболочкой. Эти образования называются эталия ми. Тип спороношения часто зависит от внешних условий: виды со спорокарпами на ножках могут развивать сидячие спорокарпы или, наоборот, ножка сильно удлиняется; виды, формирующие отдельные спорокарпы, могут дать плазмодиокарпы и т. д.
Перидий зрелого спороношения вскрывается, и споры свободна рассеиваются. У большинства слизевиков этому опособствуют находящиеся в спороношении особые нити — капиллиций, несущие на поверхности утолщения в виде спиралей, колец, полуколец, шипиков в т. п. и потому способные к гигроскопическим движениям.
При благоприятных условиях спора прорастает, образуя или от одной до нескольких зооспор с двумя гладкими, но разными по величине жгутиками на переднем конце, или миксамебы, которые в присутствии капельножидкой влаги могут вырабатывать жгутики и превращаться в зооспоры. При развитии спор из плазмодия или при прорастании спор ядра редукционно делятся. Количество зооспор и микс- амеб может увеличиваться за счет их продольного деления. Зооспоры или миксамебы попарно копулируют, образуя диплоидную миксамебу. Количество ядер в ней увеличивается за счет серии митотических делений диплоидного ядра, таким образом снова возникает плазмодий. Известно также, что диплоидные плазмодии могут сливаться друг с другом, как и с гаплоидными миксамебами (плазмогамия). Это обеспечивает разнокачественность ядер (гетерокариоз) и возможность парасексуального процесса.
Поразительная черта слизевиков этой группы — их пластичность и приспособленность к неблагоприятным условиях среды. Так, при наступлении больших морозов, отсутствии пищи, высоком осмотическом давлении, низких значениях рН, сублетальных дозах тяжелых металлов и т. п. плазмодий преобразуется в утолщенную твердеющую массу— склероций, который сохраняет жизнеспособность* в течение не
одного десятка лет. Так же долго могут сохраняться, не прорастая, споры. Если условия жизни стали неподходящими на стадии зооспор или миксамеб, то они могут инцистироваться, а затем снова прорастать.
Около 40 представителей этого класса удается культивировать и в условиях лаборатории наблюдать полный цикл их развития. Поэтому миксогастровые успешно используются как удобные объекты или модели для биохимических, биофизических, физиологических, цитологических, генетических и других исследований.
Класс миксогастровые примыкает к классу протостелиевых и может рассматриваться как дальнейшая ступень их прогрессивной эволюции. На это указывают в первую очередь одинаковые особенности зооспор (наличие двух гладких жгутиков неравной длинн) и, кроме того, сходство вегетативных тел и спороношений, хотя и более совершенных у истинных слизевиков.