

Билет1. 1)клеточные включения- необязательные компоненты клетки-делятся на запасные и экскреторные в-ва. Запасные,т.е. временно выключенные из обмена. К ним относятся а)зап-ыебелки,б)зап-ыежиры,в)зап-ые углеводы. а)-осажденные белки образуют в вакуолях алейроновые зерна. Алейроновые зерна бывают простыми(сост из аморфного белка) и сложными(помимо аморфного белка есть кристаллоид и глобоид). Кристаллоид-кристаллоподобнаяструктура,глобоид-блестящее бесцветное тельце. Типы алейроновых зерен:1)зерна с глобоидами,2)зерна с глобоидами и кристаллоидами,3)зерна с кристаллами оксалата кальция. б)-обычно распол в гиалоплазме в виде капель. Основной тип питательных веществ,ими богаты семена и плоды. Жиры-наиболее калорийное зап-ое вещ-во. Реактивом на жиры явл-сясудан 3-окраш в оранж цвет. в)-дел-ся на растворимые сахара(глюкоза,фруктоза,сахароза) и нерастворимые полисахариды(целлюлоза,крахмал). Углеводы играют роль источника энергии для реакций обмена веществ. Главным зап. углеводом является крахмал. Он бывает первичный(ассимиляционный,обр-ся в хлоропластах) и вторичный(откладывается в виде зерен в амилопластах). Зерна бывают простые концетрические(1 центр крахмалообразования в центре амилопласта) и простые эксцентрические(центр смещен),а также сложные(несколько крахмалообразующих центров),полусложные(несколько крахмалообрцентров,покрытые общими слоями). Экскреторные в-ва-продукты вторичного обмена. К ним относятся а)кристаллы оксалата кальция,б)эфирные масла,в)смолы. а)-бывают одиночные кристаллы,рафиды(игольчатые кристаллы),друзы(сростки кристаллов),кристаллический песок(скопление множества мелких кристаллов). Цистолиты-кристаллы карбоната кальция или кремнезема,откладыв в клеточной стенке. Кристаллы оксалата кальция явл-ся конечным продуктом метаболизма протопласта,накапливаются в листьях,цветках,семенах и плодах,т.к. они периодически сбрасываются,тем самым выводя излишки кальция. б)-концентрируются влистьях,цветках,семенах и плодах. Не принимают участия в обмене веществ. Скапливаются в железках,лизигенныхвместилищах,железистых волосках. в)-комплексные соединения,обр-ся в процессе норм жизн-ти или в рез-те разруш тканей. Благодаря антисепт свойствам повыш сопротивляемость растения к болезням. Нерастворимы в воде,непроницаемы для микроорганизмов. Образуются эпител-ми клетками,выстилающими смольные ходы,как побочный продукт обмена веществ. Накапл в клетсоке,цитоплазме,вместилищах.
2)Различают 4 зоны корня:а)деления,б)растяжения,в)всасывания,г)проведения. а)-верхушка конуса нарастания,где происходит митоз. Верхушечная меристема откладывает наружу клетки корневого чехлика,а внутрь-ткани ост части корня. Эта зона сост из тонкостенных паренхимныхклеток,прикрытых корневым чехликом. Корн чехлик выполн защитную функцию,его клетки пост разруш-ся,выделяя слизь. Слизь предохр зону деления при продвижении вглубь почвы. б)-клетки меристемы увел в размерах(в силу оводнения),вытянут в длину и деление постеп прекращается. Из-за вытяг клеток в длину осущ рост корня. Зоны а) и б) можно объед в зону роста,т.к. в них еще сохран меристем-я активность. На границе зоны б) и в) начин формирризодермаперв-ой коры и ЦОЦ. в)-хар-сяналич корневых волосков,являющихся выростами ризодермы. Корневые волоски увел всасывающую поверхность,обесп всасывание воды и р-ов солей. г)-в этой зоне идет интенсивное ветвление корня и появл боковые корни. Составляет большую часть корня. Тянется до корневой шейки.
3) подкласс ламииды(lamiidae). Объед 52 сем-ва. Жизн формы: деренвья,кустарники,полукуст,травы. Листья простые,безприлистников,листораспсупротивное.эволюц произошли от представ подкласса розид. Эволюц цветка шла в направ от актиноморфного к зигоморфному,околоцветник станов сростнолепестным. Гинецей ценокарпный,сост из 2 плодолистиков,завязьверхняя,полунижняя или нижняя. Порядок (Gentianales) включ 13 семейств. В их числе горечавковые,вахтовые,кутроовые,мареновые,логаниевые,ластовневые. Семейство(menyanthaceae). 5 родов,590 видов. Цветок - *Са(5)Со(5)А(2)G(2)-вахта трехлистная(menianthestrifoliata). Распростр повсеместно. Жизн форма: многолетн корневищные травы. Листья простые ,очередные,безприлист. Соцветия цимозные,в закрытых кистях или одиночные цветки. Плод-коробочка. Завязь верхняя,одногнездная,семена с эндоспермом. Вахтовые содержат горькие гликозиды и алкалоиды. Водные извлечения из листьев вахты применяются в медиц в кач-вежелчегон средства и для увелсекрецжелуд сока. Лек-ые виды: чилибуха,хинноедерево,ипекакуана,марена,горечавка желтая, золототысячник красный,вахтатрехлистная,строфант,обвойникгреческий,красавка.
4)орган раст-стебель двудол,покровная ткань-эпидерма,свп-открытые коллатеральные,тканиЦОЦ:запаспаренхима,склеренхима,камбий,флоэма,ксилема
Билет №2. 1)Макроэлементы-калий,кальций,магний,железо – металлы и азот,сера,фосфор-органогены,входящие в состав белков.Все макроэлементы абсолютно необходимы растению,каждый из них играет особую роль в жизни растения и поэтому не может быть заменен на какой-либо другой элемент.Наиболее ясна роль органогенов,они входят в состав белков.
Азот входит в состав аминогруппы во все аминокислоты.Сера входит в состав аминокислот:цистина,цистеина,метионина.Фосфор поглащается в виде иона.Он входит в состав нуклеиновых кислот ядра(ДНК и РНК),а также в состав АДФ и АТФ.Играет огромную роль в процессах дыхания,брожения и фотосинтеза.
Микроэлементы-бор,медь,цинк,молибден и др.Микроэлементы необходимы для большинства растений,т.к. влияют на физико-химическое состояние коллоидов цитоплазмы,влияют на синтез многих ферментов,хлорофилла,общий обмен веществ.Для некоторых растений список микроэлементов может быть расширен:мышьяк,йод,фтор,никель,кобольт.При отсутствии микроэлементов нарушается обмен веществ,что проявляется задержкой роста и развития,заболеваниями растений.
2)1. корень – это один из двух основных вегетативных органов;
2. корень обладает радиальной симметрией, причем характер симметрии более резко выражен в первичном строении;
3. корень – открытая система роста, он, как и стебель, продолжительное время нарастает в длину за счет деятельности апикальной (верхушечной) меристемы, но
4. в отличие от стебля точка роста корня прикрыта чехликом, поэтому апикальная меристема работает на две стороны: наращивая корень и пополняя чехлик;
5. как и стебель, корень способен ветвиться, но боковые корни эндогенны, они закладываются внутри корня в особом слое – перицикле, в то время, как боковые выросты стебля имеют экзогенное происхождение;
6. корень совершенно безлистен, на нем никогда не развиваются листья, ни простые, ни метаморфизированные;
7. корень имеет особый всасывающий слой, состоящий из корневых волосков;
8. корень имеет особую опробковевшую внутреннюю кору - эндодерму;
9. проводящие и механические ткани в корне сосредоточены в центре, а в стебле - вынесены на периферию;
10. на поперечном срезе через корень первичного строения большую часть его объема занимает первичная кора, а меньшую часть стела (центральный осевой цилиндр); в стебле же все наоборот;
11. корень нарастает только верхушкой, в то время как для побега характерен еще и вставочный (интеркалярный) рост.
3) Дуб обыкновенный —Один из самых морозостойких дубов, спокойно переносящий морозы до -30 и ниже. Характерная лесообразующая порода в зоне широколиственных лесов и лесостепи. Дуб довольно светолюбив и не очень требователен к механическому составу и химизму почвы при условиях достаточного дренажа. В благоприятных местообитаниях растет довольно быстро. Сырьем является кора, которую собирают с молодых ветвей. Кора содержит дубильные вещества, свободные галловую и эллаговую кислоты, флавоноид кверцетин и пектиновые кислоты. В виде отвара применяется как вяжущее и противовоспалительное средство для полосканий при гингивитах, стоматитах и других воспалительных заболеваниях полости рта, глотки и гортани. Может использоваться наружно при лечении ожогов. Допускается к употреблению также кора дуба скального, произрастающего на Кавказе.
Маковые (Papaveraceae) , чистотел (Chelidónium) Формула цветка:
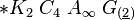 (
(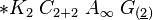 )Плод —
многосемянная стручковидная коробочка.
)Плод —
многосемянная стручковидная коробочка.
Билет №3. 1)Клеточная стенка состоит из целлюлозных микрофибрилл, погруженных в матрикс.Функции:обеспечивает сохранение формы клетки,ее механическую опору и защиту протопласта. Может расти только в контакте с протопластом.
Химический состав: Микрофибриллы(которые состоят из целлюлозы).Целлюлозный каркас заполнен молекулами матрикса(в составе матрикса:полисахариды,пектиновые вещества,гликопротеиды).
Видоизменения(происходят в процессе мацерации(разрушение срединной пластинк в мякоти созревших плодов):
-одревеснение(за счет внедрения лигнина,вызывающего жесткость,реактив:Cl-Zn-I,сине-фиолетовое окрашивание)
-опробковение(за счет суберина,реактив:Судан-3,оранжевое окрашивание)
-ослизнение(за счет углеводов-слизи,камеди;реактив-вода,набухание)
-кутинизация(за счет кутина,реактив:судан-3,оранж.окрашивание)
-минерализация(кремнезем,соли Са,Мg;реактив-сжигание,минеральный остаток)
2)Ткани первичной коры стебля:
Однодольных:ассимиляционная паренхима(осуществляется фотосинтез)
Двудольных:уголковая колленхима,ассимиляционная паренхима и эндодерма
Корня:экзодерма,мезодерма и эндодерма(однодольные-с подковообразными утолщениями,двудольные-с пятнами каспари)
3)Подкласс ламииды(Lamiidae),Семейство яснотковые или губоцветные(Lamiaceae ,Labiatae)
Жизненная форма-травы,полукустарники и кустарнички. Характерен четырехгранный стебель и супротивное листорасположение. Листья простые цельные ,без прилистников.Цветки обоеполые,зигоморфные,околоцветник двойной.Чашечка сростнолистная,пятизубчатая,правильная или неправильная. Венчик сростнолепестной. Андроцей обычно из четырех тычинок,прикрепленных тычиночными нитями к трубке венчика.(в шалфее 2 тычинки).Внутри трубки венчика имеется волосистое кольцо(приспособление для защиты нектара). Гинецей ценокарпный,из двух плодолистиков. Завязь верхняя,четырехгнездная и четырехлопастная. Плод-ценобий,распадается на 4 орешковидные доли(эремы).Семена без эндосперма.
Лекарственные виды:
-Мята перечная(Menta piperita)-содержит метнол,входящий в состав лекарственных препаратов.
-Шалфей лекарственный(Salvia officinalis) его эфирное масло обладает бактерицидным действием
-Пустырник сердечный(Leonurus cardiaca) получают препараты седативного действия и понижающие кровяное давление.
-корни Шлемника байкальского(Scutellaria baicalensis) обладают гипотензивным действием
-цветки Зайцегуба опьяняющего(Lagochilus inebrians) обладают кровоостанавливающим действием
4) Сем-во Розоцветные; *Ca5Co5A~G1 Плод- сочная или сухая костянка.
Билет №4.1.Запасные и экскреторные вещества растительной клетки.
Запасные в-ва:1.Запасные белки,относятся к простым белкам(протеинам), чаще всего откладываются в семенах.
Осаждающие белки в вакуолях образуют зерна округлой формы -алейроновые. Если алейроновые зерна не имею внутренней структуры и состоят и аморфного белка ,их называют простыми. Сложные зерна состоят из кристаллогидрата аморфного белка и глобоида. В зависимости от строения можно выелить несколько типов алейроновых зерен:
а).зерна с глобоидами (бобовые)
б).зерна с глобоидами и кристаллоидами(семян льна и клещевины)
в).зерна с кристаллами оксалата кальция(зонтичные)
2.Запасные липиды обычно встречаются в гиалоплазме в виде капель.они встречаются почти во всех растительных клетках. Наиболее богаты им семена и плоды. Реактивом на жиропадобные вещества являются судан 3,окрашивающий их в оранжевый цвет .
3.Углеводы в виде раств. В воде сахаров и нераств. полисахаридов ,входят в состав каждой клетки.
Главнейшим и наиболее запасным углеводом является полисахарид крахмал. Первичный ассимиляционный крахмал образуется в хлоропластах, а зерна вторичного крахмала образуется в особых типах лейкопластов -амилопласты.
Экскреторные включения: Растения в отличие от животных не имеют специальных выделительных органов и нередко накапливают конечные продукты жизнедеятельности протопласта в виде солей оксалата или карбоната кальция . Кристаллические включения в значительных количествах накапливаются в тканях и органах, которые растения периодически сбрасывают (листья, кора). Они откладываются исключительно в вакуолях . Форма этих включений достаточно разнообразна: одиночные многогранники - стилоиды (палочковидные кристаллы), игольчатые кристаллы - рафиды , скопления множества мелких кристаллов - кристаллический песок, сростки кристаллов - друзы ( рис. 20 ). Форма кристаллов нередко специфична для определенных таксонов и иногда используется для их микродиагностики.
К кристаллическим включениям близки цистолиты . Они чаще всего состоят из карбоната кальция или кремнезема и представляют собой гроздевидные образования, возникающие на выступах клеточной оболочки, вдающейся внутрь клетки. Цистолиты характерны для растений семейств крапивных , тутовых и др.
К экскреторным в-вам относясяи эфирные масла .они концентрируются в листьях,цветках,плодах и семенах растений.
Смолы-это комплексные соединения ,образ. В процессе жизнедеятельности. образуются эпителиальными клетками выстилающие смоляные ходы. Могут накапливаться в клеточном соке,в цитоплазме в виде капель.
2.Принцип деления на простые и сложные листья. Сложные листья и их
классификация. Анатомическое строение дорзовентрального листа.
По тому, как листовые пластинки разделены, могут быть описаны две основные формы листьев.Простой лист состоит из единственной листовой пластинки и одного черешка. Хотя он может состоять из нескольких лопастей, промежутки между этими лопастями не достигают основной жилки листа. Простой лист всегда опадает целиком. Если выемки по краю простого листа не достигают четверти полуширины листовой пластины, то такой простой лист называется цельным. Сложный лист состоит из нескольких листочков, расположенных на общем черешке (который называется рахис). Листочки, помимо своей листовой пластинки, могут иметь и свой черешок (который называется черешочек, или вторичный черешок). В сложном листе каждая пластинка опадает отдельно. Так как каждый листочек сложного листа можно рассматривать как отдельный лист, при идентификации растения очень важно определить местонахождение черешка. Сложные листья являются характерными для некоторых высших растений, таких как бобовые.
Сложные листья бывают тройчатосложные ,состоящие из 3-х листочков(земляника)и пальчатосложные(состоящие из множества листочков -каштан).Также бывают парноперисто сложные(горох посевной)и непарноперистосложные (рябина обыкнов), дважды-триждыперстосложные .
Дорзовентральный лист с обеих сторон покрыт эпидермой.Верхняя эпидерма отличается от нижней
1-более крупными клетками
2- количество устьиц меньше или отсутствует.
Под эпидермой располагается мезофил(фотосинтез и транспирация)К эпидерме верхней стороны листа примыкают клетки палисадной паренхимы. Это наиболее высокоспециализированная ткань, приспособленная к выполнению функции фотосинтеза. Число рядов составляющих ее клеток один—три и более. Это длинные цилиндрические клетки, расположенные перпендикулярно поверхности листа и соответственно параллельно солнечным лучам. Между ними есть межклетники для газообмена и транспирации. Вследствие этого мезофилл имеет очень большую поверхность — она называется внутренней поверхностью листа — и во много раз превышает его наружную поверхность. Стенка клеток тонкая, целлюлозная .Под эпидермой нижней части листа располагается губчатая паренхима. Это рыхлая ткань, состоящая из двух — семи слоев клеток разнообразной неправильной (лопастной) формы с крупными межклетниками, подходящими к устьицам эпидермы. Эта ткань выполняет функции газообмена и транспирации хотя участвует и в фотосинтезе. Число хлоропластов в ее клетках в 2.. .6 раз меньше, чем в клетках палисадной ткани. С этим часто связана разная окраска листа: сверху он ярко-зеленый, нижняя сторона вследствие содержания меньшего числа хлоропластов и обилия воздухоносных межклетников бледно-зеленая.
Мезофил листа пронизан жилками ,представляющими сосудисто волокнистые пучки.Со всех сторон пучок окружен склеренхимной обкладкой а над пучком расположена колленхима.
3.Отдел моховидные. Класс листостебельные мхи. Подкласс бриевые (зеленые) мхи,
их характеристика. Цикл развития на примере кукушкина льна.
Значение мхов в медицине.
Моховидные — это древняя, самая примитивная группа высших растений, ныне живущих на Земле. Древнейшие ископаемые формы мхов известны с каменноугольного периода
Характерные прзнаки 1-отсутствие корней(ризоиды) 2- преоблодание гаметофазы 3-гаметофит представлен слоевищем (у примит) у остальных расчленен на стебель и листья.4-половое и безполое поколение существуют вместе.
КЛАСС ЛИСТОСТЕБЕЛЬНЫЕ МХИ - BRYOPSIDA, MUSCI
В отличие от печеночных мхов все представители листостебельных имеют ветвящийся или неветвящийся стебель и расположенные на нем листья. Протонема лиственных мхов чаще нитевидная, реже — в виде пластинки. У наиболее развитых представителей этого типа высших растений развивается перистом, способствующий рассеиванию спор.
Подкласс Бриевые, или Зеленые Мхи (Bryidae).
Подкласс бриевых, или зеленых, мхов — наиболее многочисленная группа мохообразных. Протонема этих мхов обычно имеет вид сильноразветвленной, стелющейся по поверхности субстрата нити, от которой в почву отходят незеленые ризоиды
Представитель кукушкин лен. Кукушкин лен — многолетнее растение. Его коричневато-зеленые стебли неветвящиеся, густо покрыты узкими листьями, иногда достигают в высоту 30 см Нижняя часть стебля темно-коричневая. Корней у кукушкина льна, как и у остальных мхов, нет. На нижних частях стеблей имеются нитевидные выросты покровной ткани — ризоиды.
Питаются зеленые мхи так же, как другие зеленые растения. Из почвы ризоиды всасывают воду и минеральные вещества. Листья мхов, в клетках которых содержится хлорофилл, поглощают углекислый газ. В хлоропластах происходит фотосинтез.
Размножение кукушкина льна: Кукушкин лен — двудомное растение. В начале лета на кукушкином льне имеются своеобразные красно-желтые верхушки некоторых стебельков. Здесь между листочками находятся удлиненные мешочки из большого количества клеток. Это мужские половые органы, в которых образуются подвижные мужские гаметы — сперматозоиды.
На верхушках женских экземпляров, которые не имеют ярко окрашенных листьев, образуются женские половые органы. Они похожи на колбочки с длинной шейкой. Их наружные стенки состоят из одного слоя клеток. В расширенной части колбочки помещается женская гамета — яйцеклетка.
Мхи растут густыми дернинками, а значит, мужские и женские растения оказываются рядом. Дождевая вода помогает попасть сперматозоидам на верхушки женских растений. Передвигаясь в воде, сперматозоиды проникают к яйцеклеткам. Содержимое сперматозоида сливается с содержимым яйцеклетки. Происходит оплодотворение, и образуется зигота.
На следующий год из зиготы развивается коробочка со спорами на длинной ножке. Чем выше расположена коробочка, тем дальше могут разлетаться споры, тем лучше будет расселяться мох. Отсюда понятно значение ножки. Коробочка снабжена крышечкой и сверху прикрыта колпачком. Внутри коробочки находится спорангий со спорами. Когда споры созреют, в сухую погоду колпачок, а за ним и крышечка опадают. Тонкая ножка раскачивается даже от слабого дуновения ветра, и споры высыпаются.
Спора попадает на влажную почву и прорастает, образуя тонкую зеленую нить-протонему. Нить ветвится; на ней появляются почки, из которых вырастают побеги мха.
Мхи способны впитывать и удерживать большое количество воды, благодаря чему играют значительную роль в регулировании водного баланса ландшафтов.
Кроме того, они обеспечивают равномерный перевод поверхностного стока вод в подземный, предохраняя тем самым почвы от эрозии.
Экономическое значение имеют виды мхов, произрастающих в болотистой местности:
Остатки сфагнума или торфяного мха накапливаются под водой и образуют торф, который можно использовать в качестве топлива и как упаковочный материал при транспортировке. Способность его хорошо удерживать влагу, важна при удобрении полей.
Торф также является сырьем для химической промышленности при получении спиртов, фенола, смол, пластиков и других материалов.
Используют мхи в строительстве как утеплительный и изоляционный материал
Находят применение в медицине благодаря своим антибиотическим (антимикробным) свойствам
4) стебель однодольного строения
покровная ткань: эпидерма
СВП: закрытые коллатеральные пучки
ткани, входящие в ЦОЦ: склеренхима, паренхима, флоэма, ксилема
Билет №5. 1.Тропизмы (от греч. ?????? — поворот, направление) — реакция ориентирования клетки то есть направление роста или движения клеток относительно раздражителя (химического, светового и др.).Если растение под влиянием раздражителя изгибается к источнику раздражителя, то это положительный тропизм, а если оно изгибается в противоположную сторону от раздражителя, то это отрицательный тропизм.
Ортотропизм — расположение органа растения вдоль градиента раздражителя.
Диатропизм — расположение под прямым углом к градиенту раздражителя.
Плагиотропизм — ориентация под любыми другими углами.
В основе тропизма лежит одно из свойств цитоплазмы клетки — её раздражимость, как ответной реакции на различные факторы внешней среды.
Ответные реакции растений на различные односторонние воздействия раздражителей внешней среды (свет, земное притяжение, химические вещества и др.) заключаются в направленных ростовых и сократительных движениях (изгибах) органов растения, приводящих к изменению его ориентации в пространстве. Ростовые движения зависят от вида раздражителя, механизм действия которого на растения сложен. Эти движения могут возникать в растущих частях растений, как следствие более быстрого роста клеток, расположенных на одной стороне органа растения (стебле, корне, листе). В органах растения возникают растяжения, связанные с асимметричным распределением в них фитогормонов роста растений — ауксина и абсцизовой кислоты и др.
Тропизмы различают в зависимости от вида раздражителя.
Геотропизм связан с воздействием на растения силы тяжести Земли. При положительном геотропизме рост главного корня направлен строго вниз по направлению к центру Земли, что связано не только с деятельностью гормонов, но и с особыми крахмальными зёрнами в корневом чехлике, выполняющим роль статолита. Отрицательный геотропизм характерен для главного стебля.
Фототропизм вызывает направленный изгиб растения к источнику света. Этот изгиб имеет химическую природу. Под влиянием фитогормона ауксина на теневой стороне деление и рост клеток интенсивнее по сравнению со световой стороной, где ауксина меньше и рост клеток замедлен. В связи с этим растение изгибается в сторону клеток медленно растущих, то есть к свету. У стеблей наблюдается положительный фототропизм, корней — отрицательный, листьев — поперечный. Примером поперечного гелиотропизма, который свойствен, скажем, листьям растений, живущих в засушливых зонах, например, листьям эвкалиптовых деревьев. В солнечный день эти листья поворачиваются ребром и пропускают солнечные лучи мимо себя так, что найти тень в эвкалиптовой роще является нелегкой задачей. Такие деревья демонстрируют, так сказать, «обратный эффект жалюзи».
Благодаря положительному фототропизму растения образуют листовую мозаику, то есть листья в пространстве располагаются так, чтобы максимально использовать свет.
Хемотропизм вызывает движение растений под влиянием химических соединений. Наиболее яркий пример хемотропизма — рост корней в сторону больших концентраций питательных веществ в почве.
2.Выделительные системы в жизнедеятельности растений играют важную, разнообразную роль. В процессе их жизнедеятельности образуется ряд веществ с разнообразной химической природой, не участвующих в дальнейшем метаболизме.
Выделительные структуры растений характеризует,то что клетки выделительных тканей паренхимные,тонкостенные.Их ультраструктура связана с секретирующим веществом. В тканях, где синтезируются эфирные масла,смолы, каучук,имеется хорошо развитый агранулярный ЭР, слизи - аппарат Гольджи.
Выделительные ткани подразделяются на ткани внешней(наружной) и внутренней секреции.
Наружные выделительные структуры(внешняя секреция).Связаны эволюционно с покровными тканями. Они представлены следующими структурами:железистые волоски, желёзки,переваривающие желёзки, нектарники,осмофоры, гидатоды.
Железистые волоски и желёзки представляют собой трихомы эпидермы. Они состоят из живых клеток,обычно имеют удлиненную ножку из одной или нескольких клеток и одно-или многоклеточную головку. Клетки головки выделяют секрет под кутикулу. При разрыве кутикулы вещество изливается наружу, после чего может образовываться новая кутикула и накопиться новая капля секрета. Железистые волоски цветков герани, листьев и цветков душистого табака выделяют эфирные масла;сидячие головчатые волоски, образующие мучнистый налёт на листьях мари и лебеды, - воду и соли. Желёзки отличаются от волосков короткой ножкой из несекретирующих клеток и многоклеточной головкой. Они характерны у мяты, лаванды,полыни, черной смородины.
Нектарники обычно образуются на частях цветка,но могут встречаться и на других надземных органах растения.Они могут быть представлены отдельными поверхностными железистыми клетками или находиться в ямках (лютик),желобках, шпорцах(живокость),возвышаться в виде бугорков,подушечек(тыква, ива, яснотка белая). Выделительные клетки нектарников отличаются густой цитоплазмой и высокой активностью обмена веществ.К нектарнику может подходить проводящий пучок.
Осмофоры представляют собой или специализированные клетки эпидермы,или особые желёзки, где секретируются ароматические вещества. Выделение летучего секрета происходит в течение короткого времени и связано с использованием запасных веществ.Аромат цветка создается секрецией сложной смеси органических соединений,главным образом эфирных масел.
Гидатоды выделяют капельно-жидкую воду и растворенные в ней соли. При избытке воды и ослаблении транспирации через гидатоды происходит гуттация –выделение капель воды из внутренних частей листа на его поверхность.Специальной секреторной ткани здесь,как правило нет. Гидатоды могут иметь вид многоклеточных волосков, устьиц,потерявших способность регулировать величину своей щели, и, наконец представлять собой специальное образование из большого числа клеток,расположенных под водным устьицем.
Гидатоды в виде многоклеточных волосков имются у фасоли огненно-красной,а в виде водных устьиц встречается у манжетки,настурции.Наиболее сложно устроенные гидатоды находятся в зубцах листьев камнеломки,земляники,шиповника, чая.Здесь под водными устьицами образуется многоклеточная ткань эпитема,через которую вода передается от водопроводящих элементов к отверстиям устьиц.
Преваривющие желёзки на листьях насекомоядных растений, например росянки, венериной мухоловки и др., выделяют жидкость, содержащую пищеарительные ферменты и кислоты.
Внутренние выделительные структуры(внутренняя секреция).Вырабатывают и накапливают вещества, остающиеся внутри растения.Это могут быть отдельные секреторные клетки, рассеяные среди других тканей, как идиобласты.Они содержат различные вещества, особенно часто оксалат натрия в виде одиночных кристаллов,друз или рафид,бальзамы, танины,слизи и др.
Секреторные вместилища разнообразны по форме и происхождению.Схизогенные вместилища образуются вследствие расхождения клеток и формирования межклетника,выстланного живыми эпителиальными клетками и заполненного выделенными веществами.К ним относятся смоляные ходы хвойных растений.Лизигенные вместилища возникают в результате растворения группы клеток с продуктами секреции. Такие вместилища видны в кожуре плодов цитрусовых(апельсина,лимона, мандарина).
Млечники –особый тип выделительной ткани. Это живые клетки (нечленистые млечники) или ряды слившихся клеток (членистые млечники),пронизывающие всё растение.В зрелом млечнике протопласт занимает постенное положение,полость млечника занята млечным соком – латексом,млечные клетки неодревесневающие,эластичные.Латекс представляет собой эмульсию белого, реже оранжевого или красного цвета. Жидкая основа латекса– клеточный сок, в котором растворены или взвешены углеводы (крахмальные зерна у молочайных,сахар у астровых),белки (у фикуса),жиры танины,слизи, эфирные масла, каучук(более чем у 12 500 растений). Среди каучуконосов промышленное значение имеет тропическая гевея – Hevea (семейство Молочайные),в млечном соке которой содержится40-50% каучука.
3.Ламииды происходят, по-видимому, от древних представителей подкласса розид и представляют мощную эволюционную ветвь, характермзующуюся высокой специализацией сростно лепестного, обычно сросшегося околоцветника. Они объединяют 11 порядков, 51 семейство, около 2400 родов и почти 40000 видов. Для цветка представителей ламиид характерно небольшое фиксированное число частей и "наростание" зигоморфии. Гинецей всегда ценокарпный и состоит большей чатью из 2 плодолистиков, образующих верхнюю, полунижнюю или нижнию завязь. Наряду с деревьями и кустарниками в подклассе очень много травянистых, иногда довольно специализированных форм.
Порядок Губоцветные – Lamiales
Многолетние и однолетние травы, полукустарники, реже кустарники и деревья. Листья большей частью простые, без прилистников, супротивные или мутовчатые. Цветки зигоморфные со сростнолистной чашечкой и спайнолепестным пятичленным венчиком; тычинок 4-2 (очень редко 5 или 1). Гинецей из 2 плодолистиков, завязь верхняя. Плод из 4 орешков (мерикарпиев), редко костянковидный.
Семейство Губоцветные, или Яснотковые – Lamiaceae, или Labiatae
Травы или полукустарники, редко кустарники с супротивными цельными, часто зубчатыми, пильчатыми или лопастными листьями без прилистников. Цветки обоеполые, зигоморфные, редко почти правильные, часто в тирсоидных соцветиях из двойных завитков. Околоцветник 5-членный. Чашечка сростнолистная, лопастная, зубчатая или 2-губая. Венчик, как правило, 2-губый, с 3-лопастной нижней губой и более или менее длинной трубкой, 4- или 2-лопастный диск. Тычинок 4, из них 2 более длинные, реже их 2. Завязь верхняя, 2– 4-гнездная и часто 4-лопастная, с 4 суббазальными анатропными семязачатками с 1 интегументом. Столбик гинобазический, с 2 рыльцами. Плод распадается на 4 эрема. Семена без эндосперма. Характерны различные ароматические вещества: спирты, фенолы, альдегиды, кетоны и др.
Губоцветные легко отличаются от похожих на них по цветкам семейств (Вербеновых, Норичниковых, Акантовых и др.) прежде всего 4-лопастной завязью с гинобазическим столбиком и плодами, распадающимися на 4 орешковидных эрема (как у Бурачниковых). Модель цветка Губоцветных (резко 2-губый венчик с трубкой и пыльниками, расположенными как бы в 2 яруса под верхней губой, нектароносный диск), несомненно, устанавливалась в связи с эволюцией перепончатокрылых – основных опылителей Губоцветных. Впрочем, цветки Губоцветных посещаются и другими насекомыми, а также колибри. В качестве особо специализированных цветков можно рассмотреть цветки рода шалфей (Salvia). У шалфеев всего 2 тычинки, причем у каждой остается фертильной лишь половина пыльника, другая же половина стерилизуется, превращается в своеобразное рычажное устройство. Стерильная и фертильная половины пыльника соединены длинными нитевидными связниками, а стерильные половины обоих пыльников срастаются. Сами же тычиночные нити очень короткие. Когда шмель наступает на площадку из сросшихся стерильных гнезд, рычажное устройство приводится в действие и пыльца из фертильных гнезд попадает на спинку насекомого, а вскоре – на рыльце пестика другого цветка. Хотя у большинства представителей семейства венчики 2-губые, они довольно разнообразны в деталях. Впрочем, у некоторых родов цветки почти правильные, например, у мяты (Mentha). Иногда развита лишь нижняя губа венчика, как у видов рода живучка (Ajuga).
Хотя Губоцветные довольно многочисленны и в тропиках, и в умеренно холодных областях, их настоящее царство – так называемая область Древнего Средиземья, от Центральной Азии до Пиренейского полуострова. Здесь они представлены не только множеством родов и видов, но обладают высокой численностью, часто доминируя в растительных группировках. Во многих сухих горных районах воздух буквально пропитан запахами Губоцветных. В семействе выделяют несколько подсемейств, из них два наиболее крупные.
Подсемейство шлемниковые (Scutellarioideae).
Цветки с двугубой (губы цельнокрайние) чашечкой и двугубым венчиком, верхняя губа которого шлемовидная. Тычинок 4.
Род шлемник (Scutellaria). Многолетние травы или полукустарники, редко кустарнички с опушением из простых волосков или лишь с примесью железистых (без запаха).
Подсемейство Чистецовые (Stachyoideae).
Самое крупное подсемейство, к которому относится большинство родов Губоцветных, представленных во флоре России. Цветки с чашечкой различной формы (если двугубой, то губы не цельнокрайние). Венчик от почти правильного до двугубого. Тычинок 4-2.
Очень крупный род, в России представлен многочисленными видами род котовник (Nepeta). К этому семейству также относятся роды – змееголовник (Dracocephalum), мята (Mentha), шалфей (Salvia), тимьян (Thymus), зопник (Phlomis), пустырник (Leonurus), яснотка (Lamium), чистец (Stachys) и др. (рис.26).
Основное практическое значение семейства как раз и связано с их ароматическими веществами. Как сырье для парфюмерной промышленности культивируются розмарин (Rosmarinus officinalis), лаванда (Lavandula spica) и целый ряд других видов. Плантации лаванды имеются, например, в Северном Крыму. Из различных видов мяты получают лекарственные мятные капли и ментол.
Шалфей луговой (Salvia pratensis): 1 – цветущее растение; 2 – схематический продольный разрез цветка с насекомым, нажимающим на сросшиеся стерильные гнезда пыльников (с.г.п). Шалфей лекарственный (Salvia officinalis): 3 – плод; 4 – диаграмма цветка. Мята (Mentha longifolia): 5 – верхняя часть цветущего растения; 6 – цветок. Дубровник (Teucrium chamaedrys): 7 – цветок. Глухая крапива крапчатая (Lamium maculatum): 8 – нижняя часть цветка в продольном разрезе. Глухая крапива белая (Lamium album): 9 – диаграмма цветка.
Как лекарственные растения особенно хорошо известны виды рода пустырник (Leonurus), обитающие и в средней полосе. Препараты пустырника употребляют как успокаивающие и понижающие кровяное давление. Наконец, многие губоцветные – прекрасные медоносы, например мелисса (Melissa officinalis) и душица (Origanum vulgare). Мед губоцветных отличается особым вкусом и ароматом. Приправа из листьев базилика (Ocimum basilicum) – непременная принадлежность кавказской кухни. Многие введены в культуру (мелисса, лаванда, иссоп и др.).
Виды, внесенные в Красную книгу Республики Бурятия
Змееголовник Стеллера – Dragocephalum stellerianum Hiltebr.
Лагопсис мохнатоколосый – Lagopsis eriostachya (Benth.) Ik.-Gal. ex Knorr.
Тимьян еравнинский – Thymus eravinensis Serg.
4) орган раст-стебель двудол,покровная ткань-эпидерма,свп-открытые коллатеральные,ткани ЦОЦ: запас. паренхима,склеренхима,камбий,флоэма,ксилема
Билет №6. 1.Пластиды и их пигменты
Пластиды это органеллы,характерные исключительно для растительных клеток. В них происходит первичный и вторичный синтез углеводов.Формы, размеры,строение и функции пластид различны. По окраске (наличию или отсутствию пигментов)Различают три типа пластид:
Зеленые хлоропласты – это органеллы фотосинтеза, относительно крупные структуры (5-10 мкм. в длину при диаметре 2-4 мкм.) овальной или дисковидной формы. Содержимое пластид называют стромой (матрикс). В строме содержатся белки, липиды, ДНК (кольцевая молекула), РНК, рибосомы и запасные вещества (липиды, крахмальные и белковые зерна). Кроме того, в строме содержатся также ферменты, участвующие в фиксации диоксида углерода. Наружная мембрана гладкая. Внутренняя – образует пластинчатые впячивания – тилакоиды, большая часть которых укладывается в виде стопки монет и образует граны. Хлоропласты образуются из пропластид – мельчайших сферических недеференцированных телец, котрые содержатся в растущих частях растения (в клетках зародыша, образовательной ткани) и имеют зеленый цвет, обусловленный присутствием фотосинтезирующим пигмента хлорофилла. Существует несколько видов хлорофилла.
Наиболее распространен хлорофилл а(найден у всех зеленых растений и цианобактерий)имеет голубовато-зеленый цвет
Хлорофилл б – желтовато-зеленый.Его молекула содержит на один атом кислорода больше и на два атома водорода меньше.
В процессе фотосинтеза хлорофиллу принадлежит ведущая роль.Он может поглощать солнечную энергию, запасать ее или передавать ее другим молекулам.
Кроме того в состав хлоропластов входят желтые пигменты –каротиноиды.Каротиноиды представляют собой высокомолекулярные углеводороды:оранжевый каротин С40Н56и желтый ксантофилл С40Н56О2.Каротиноиды хлоропластов,а также синие,красные, бурые пигменты хроматофоров водорослей называют дополнительными,вспомогательными пигментами,поскольку энергия, поглощенная ими, может передаватться на хлорофилл а. Хлорофилл использует энергию красной части спектра,каротиноиды– синей. По своему строению каротиноиды являются либо углеводородами(каротины), либо окисленными углеводородами,т. е. Кислородсодержащими(каротинолы или ксантофиллы).Кроме того каротиноиды могут защищать молекулы хлорофилла от чрезмерного фотоокисления на ярком свету.
Лейкопласты – бесцветные округлые пластиды, в которых обычно накапливаются запасные питательные вещества, в основном крахмал. По строению лейкопласты мало отличаются от протопластид, из которых он образуется: двумембранная оболочка окружает бесструктурную строму. Внутренняя мембрана, врастая в строму, образует немногочисленные тилакоиды. В лейкопластах имеются ДНК, рибосомы, а также ферменты, осуществляющие синтез и гидролиз запасных веществ, в первую очередь крахмала. Лейкопласты, в которых синтезируется и накапливается запасной крахмал, называют амилопластами, белки – протеинопластами, масла – элайопластами. В одено лейкопласте могут накапливаться разные вещества. Запасной белок может откладываться форме кристаллов или аморфных включений, масла – в виде пластоглобул. Однако белки и масла в лейкопластах встречаются довольно редко. В амилопластах в связи с тилакоидами в строме возникают образовательные центры, вокруг которых в виде зерен откладывается вторичный запасной крахмал из растворимых углеводов, образовавшихся в хлоропластах в процессе фотосинтеза. Много амилопластов в клетках клубней картофеля, зерновок ржи, пшеницы и других органах растений где откладываются запасные вещества. В секреторных клетках они в комплексе с агранулярным ретикуломом участвую в синтезе эфирных масел.
Хромопласты – пластиды оранжево-красного и желтого цвета, образующиеся из лейкопластов и хлоропластов в результате накопления в их строме каротиноидов. Они встречаются в клетках лепестков (лютик, нарцисс, тюльпан, одуванчик), зрелых плодов (томат, тыква, арбуз, апельсин), редко – корнеплодов (морковь, кормовая свекла), а также в осенних листьях.
Хромопласты– конечный этап в развитии пластид. По форме накопления каротиноидов различают следующие типы хромопластов:глобулярный пигменты растворены в липидных пластоглобулах;фибриллярный– пигменты накапливаются в белковых нитях; кристалический– пигменты откладываются в виде кристалов.
Косвенное биологическое значение хромопластов в том, что ярко окрашенные плоды успешнее распространяются птицами и животными,а выделяющиеся яркой желто-красной окраской цветки привлекают насекомых –опылителей.
В филогенезе первичным исходным типом пластид являются хлоропласты,из которых в связи со специализацией органов произошли лейко- и хромопласты.В онтогенезе взаимопревращения пластид происходит иными путями.Наиболее часто хлоропласты превращаются в хромопласты при осеннем пожелтении листьев или созревании плодов. В природе этот процесс необратим.Лейкопласты могут превращаться в хлоропласты(позеленение верхней части корнеплода моркови, оказавшейся на поверхности почвы) или хромопласты.Хлоропласты могут при помещении растения в темноту превратиться в лейкопласты.Процесс этот обратимимый.
2 Ко?рень (лат. radix) — осевой, обычно подземный вегетативный орган высших растений (сосудистых растений), обладающий неограниченным ростом в длину и положительным геотропизмом. Корень осуществляет закрепление растения в почве и обеспечивает поглощение и проведение воды с растворёнными минеральными веществами к стеблю и листьям.
На корне нет листьев, в клетках корня нет хлоропластов.
Кроме основного корня, многие растения имеют боковые и придаточные корни. Совокупность всех корней растения называют корневой системой. В случае, когда главный корень незначительно выражен, а придаточные корни выражены значительно, корневая система называется мочковатой. Если главный корень выражен значительно, корневая система называется стержневой.
Некоторые растения откладывают в корне запасные питательные вещества, такие образования называют корнеплодами.
Части корня
Точка роста. Участок непосредственно за кончиком корня, где клетки делятся, обеспечивая новый рост.
Зона растяжения. Участок новых клеток, образованных в точке роста и расположенных непосредственно за ней. Клетки, пока их клеточные стенки не станут жёсткими, растягиваются в длину при всасывании воды. Это растяжение толкает кончик корня дальше в почву.
Слой корневых волосков. Наиболее молодая часть эпидермиса или внешней кожицы корня. Это зона, где образуются корневые волоски. Она расположена непосредственно за зоной растяжения. Как только стенки вытянувшихся клеток становятся жёстче, самые крайние из них превращаются в слой корневых волосков. Более старая часть этого слоя (выше по корню) медленно отделяется, замещаясь слоем жёстких клеток, входящих в экзодерму (самый наружный слой коры).
Корневые волоски. Длинные выросты клеток слоя корневых волосков. Они всасывают воду и минеральные вещества.
Корневой чехлик. Слой клеток, защищающих кончик корня при росте.
Ризодерма — покровная ткань, которой снаружи покрыты молодые корневые окончания. Она содержит корневые волоски и участвует в процессах всасывания. В зоне всасывания ризодерма пассивно или активно поглощает элементы минерального питания, затрачивая в последнем случае энергию. В связи с этим клетки ризодермы богаты митохондриями.
Экзодерма — опробковевшая покровная ткань, приходящая на смену отмирающей ризодерме.
Мезодера- основная масса первичной коры.
Эндодерма- самый внутренний однорядный слой первичной коры. Выполняет роль барьера.у онодольних эндодерма с подковообразными утолщением.
Осевой цилиндр — представляет собой сложный комплекс из проводящей, образовательной и основной тканей.
Корневи?ще — видоизменённый побег, обычно подземный, с чешуевидными, недоразвитыми или рано отмирающими листьями, верхушечными или пазушными почками, придаточными корнями. Корневище выполняет функцию запасания питательных веществ, вегетативного возобновления и размножения. У дерева корневищем называют основной корень (продолжение ствола), а также часть корней, выступающую на поверхности земли.[1]
Корневище может быть:
простым или ветвистым — по степени разветвлённости;
горизонтальным, вертикальным или косым — по направлению роста;
длинным, коротким или сильно укороченным (со сближенными узлами) — по длине;
толстым или тонким — по толщине.
Эпигенное корневище
Гипогенное
у однодольного состоит из покровной ткани(эпидерма),запасающая паренхима,эндодерма с подковообразными утолщениями,перицикл закрытые колатеральные пучки паренхима запасающая.
3.Папоротниковидные, или папоротники, возникли в девонский период, а уже в карбоне их древовидные формы наравне с другими высшими споровыми растениями составляли обширные влажные леса, остатки которых образовали залежи каменного угля, К настоящему времени сохранилось примерно 12 тыс. видов папоротников, распространенных преимущественно в тропиках и субтропиках, а также в умеренных областях земного шара, включая пустынные районы. Папоротники умеренной зоны Северного полушария растут в сырых тенистых лесах, по кустарникам, лесным оврагам, сырым лугам, некоторые виды встречаются в сухих сосновых лесах (например, орляк обыкновенный).
Большинство современных представителей этого отдела — наземные многолетние растения, однако есть и водные формы (например, реликтовый вид однолетняя сальвиния плавающая, занесенная в Красную книгу Республики Беларусь).
В тропических лесах встречаются древовидные папоротники высотой 20—25 м и диаметром ствола до 0,5 м, а также немало лиановидных форм и эпифитов, растущих на стволах и ветвях деревьев.
Листья папоротников (часто их называют вайями) крупные с перисто-рассеченной листовой пластинкой и хорошо развитой проводящей системой. Общий черешок листа прикрепляется к подземному стеблю, представляющему собой корневище. Корни у папоротников придаточные, а листья, возникшие в результате уплощения крупных ветвей, растут, своей верхушкой образуя характерную разворачивающуюся «улитку». Длина листьев может колебаться от 1—2мм до 10м.
У некоторых папоротников (например, у страусника) листья дифференцированы на стерильные (фотосинтезирующие) и фертильные (несущие спорангии). Однако у большинства представителей листья выполняют не только функцию фотосинтеза, но и спорообразования. На их нижней стороне летом образуются спорангии, расположенные одиночно или группами — сорусами. Сорусы, имеющие вид буроватых бугорков, прикрыты особым выростом листа — индузием, или покрывальцем.
При созревании спорангии вскрываются, и споры высыпаются, подхватываются и переносятся ветром на огромные расстояния от материнского организма. В благоприятных условиях они прорастают в заростки (гаметофиты).
Большинство папоротников — равноспоровые растения; из одинаковых спор развиваются обоеполые гаметофиты (заростки). Они зеленые, мелкие, не более 0,5 см в поперечнике, разные по форме (пластинчатые, сердцевидные, нитчатые и др.), питаются самостоятельно. К почве гаметофит прикреплен ризоидами. На нижней стороне заростка образуются архегонии (с яйцеклеткой) и антеридии, в которых формируются сперматозоиды. Оплодотворение происходит только при наличии воды, обеспечивающей активное движение сперматозоидов к архегонию. Оплодотворенная яйцеклетка (зигота) дает начало диплоидному зародышу, развивающемуся во взрослое растение
На нижней стороне вайи образуются сорусы(собрание спорангиев).В спорангиях мейозом образуютс споры .При их созревании внешние стенки кольца клеток усыхает,стенка спорангия разрывается поперек в области устья и споры высыпаются.из спор образуется гаплоидный заросток (сердцевидная пластинка с ризоидами )между ризоидами образуются антеридии а в области сердцевидной выемки -архегонии (брюшко погружено в тело заростка)Во время дождя антеридии раскрываются и штопорообразно извитые сперматазоиды проникают в архегоний и происходит оплодотворение.из зигоы образуется зародыш который в дальнейшем переходит в самостоятельный спорофит.
4)Сем-во Яснотковые
(губоцветные) – Lamiaceae стрелочка Ca(5) Co(2/3) A4 G(2)_; Плод – ценокарпий: ценобий, распадающийся на 4 орешка.
Билет 7
1)Красные водоросли, как и бурые, почти все обитают в морях. Это многоклеточные растения, достигающие значительной величины (до 2 м), их слоевище обычно так расчленено, что напоминает стебель и листья. Красные водоросли в своих мелких хроматофорах, кроме хлорофилла, содержат пигмент красного цвета — фикоэритрин и другие пигменты. Окраска этих пигментов обусловливает окраску водорослей в розовый, красный, синеватый и другие цвета. Известно около 4 000 видов. Размножаются вегетативно, спорами и половым путем. Наиболее известными являются порфира, радимения.
Практическое значение багрянок для человека велико. Одну из живущих в Северном море водоросль — хондрус — употребляют как лекарство при заболевании дыхательных путей. Красные водоросли используются для добывания растительного студия агар-агара, применяемого в кондитерском производстве и в микробиологии — для получения чистых культур микробов. Природные запасы багрянок в нашей стране незначительны, поэтому ученые занимаются разработкой научных основ выращивания красных водорослей в Черном море
Половой процесс всегда оогамный. После оплодотворения, образовавшаяся зигота претерпевает сложное развитие прямо на гаметофите и дает начало особым спорам, которые называются карпоспорами, образующиеся в карпоспорангиях, тогда как у многих других водорослей зигота развивается в спорофит, давая начало новой форме развития растений.
2) Занимают наибольший объем. Снаружи они защищены покрывными тканями, а изнутри пронизаны проводящими и механическими пучками. Образующие их клетки отличаются обычно паренхимным строением, из-за этого основные ткани нередко называют Основной паренхимой. Основные ткани являются питающими тканями, хотя могут выполнять многие функции, начиная от ассимиляционной, кончая выделительной. В зависимости от специализации, основную паренхиму делят наассимиляционную, вентиляционную, всасывающую, запасающую и выделительную.Ассимиляционные (фотосинтези-рующие) ткани они имеют зеленую окраску – Хлоренхима. Ассим. ткань состоит из однородных тонкостенных клеток. Доступ углекислоты к клеткам облегчается тем, что в ней имеется развитая система межклетников, сообщающаяся с атмосферой. Наличие межклетников является характернейшей особенностью ассимиляционной ткани. Межклетники обеспечивают газообмен с окружающей средой. Хлоренхима располагается в местах, наиболее доступных свету: в надземной части растений непосредственно под кожицей листьев и стеблей. Нередко хлоренхима дифференцирована наСтолбчатую (палисадную) и Губчатую ткань. Палисадная ткань содержит большое количество хлоропластов, здесь происходят световые реакции фотосинтеза. Губчатая ткань - протекает газообмен и темновая стадия фотосинтеза.Вентиляционная ткань (Аэренхима) во всех тканях имеются межклетники, образующие единую систему. Межклетники через проходные отверстия или устьица сообщаются с атмосферой. Однако газовый состав в межклетниках сильно отличается от газового состава атмосферы. Запасающие ткани Поглощенные растением синтезированные вещества могут откладываться в виде запасов: накопление воды, пластические вещества. сахар, инулин, аминокислоты, белки, крахмал.
3) Маковые (лат. Papaveraceae) — семейство двудольных растений порядкаЛютикоцветные.
Однолетние и многолетние травы, содержащие нередко млечный сок; листьяочередные, изредка противоположные. Деревянистые растения встречаются лишь в родах боккония и дендромекон. Цветы или одинокие, или образуют кистеобразные соцветия. Они обоеполые, правильные или неправильные; в чашечке по большей части 2 листика, в венчике 4 лепестка, но бывают уклонения; тычинки, которых много, расположены в 2 или 4 кружка (не без уклонений), реже только 4 или 2; в последнем случае они разветвляются с самого основания. Гинецей содержит от 2 до 16 пестиков, сросшихся до верхушки и образующих одну цельную завязь, по большей части одногнёздую.Плод сухой, коробчатый, одногнёздый или разделен на неполные гнезда, редко ореховидный. Семена с маслянистым белком и маленьким зародышем, который у шишконосных снабжён только одной семядолей вместо двух. Опыление происходит преимущественно помощью насекомых.
Маковые делятся на 2 подсемейства: маковые (Papaveroideae) и гипекоумовые (Hypecoideae). Ранее в состав семейства маковых включалось также подсемейство дымянковые (Fumarioideae), выделенное ныне в отдельное семейство. Подсемейство маковых, состоящее из 26 родов и более 450 видов, — самое большое по объему. Из его представителей наиболее широко распространенным и самым известным родом является мак (Papaver). Представители семейства маковых содержат многочисленные и разнообразные алкалоиды — производные изохинолина. Названия этих веществ — папаверин, адлюмин, бикукулин, глауцин, фумарин и т. д.— ясно указывают источники сырья. Лекарственные свойства маковых были известны издавна. Лекарственными свойствами, широко используемыми в народной медицине, обладает такой представитель данного семейства, как чистотел.
Сильно ядовитый сок этого растения содержит 6-7 алкалоидов: хелидонин, гомохелидонин, протонин, сангуинарин и др. Его применяют для выведения бородавок, отсюда и русское его название.Все маковые очень декоративны. Многие из них давно введены в садовую культуру. Дикорастущие представители семейства маковых благодаря декоративности усиленно истребляются. Несколько видов из этого семейства, произрастающие на территории России, внесены в "Красную книгу".
4)корень, тип покровной ткани- пробка, входящая в состав перидермы, типы возможных механических тканей - склеренхима,
БИЛЕТ 8 1)Биологическая роль элементов минерального питания.
Калий поддержанием гомеостаза внутриклеточной и тканевой среды Калий, как активатор ферментных систем.
Кальций. Накопление, формы соединений, особенности поступления и перемещения Са2+ по растению, возможность его реутилизации Посредничество Са2+ в управлении комплексными физиологическими процессами: устьичные движения; рост кончиком (пыльцевая трубка и корневой волосок); гравиотропизм; фототропизм; фотоморфогенез; адаптация к стрессу и др...
Магний. Содержание и соединения магния в тканях растений. Значение связи Mg2+ с аденозинфосфатами и фосфорилированными сахарами. Роль в поддержании рН-стата и дальнем транспорте веществ. Запасные формы Mg2+ , его реутилизация и перераспределение в растении. Функции магния в фотосинтезе. Магний - как активатор ферментных систем; роль в образовании функциональных рибосом.
Азот..
Фосфор. Доступность фосфора в почве, особенности транспорта в корни и зависимость поглощения Pi от условий среды. Формы Р-соединений (органические и неорганические) и их распределение в клеточных структурах, тканях и органах. Запасание фосфора. Особенности транспорта соединений фосфора. Метаболическая компартментация его соединений в клетке. Функции роль фосфора в запасании и превращении энергии, структурной организации мембран, кодировании и реализации наследственной информации, обмене углеводов. Фосфор и метаболитная регуляция процессов. 1,4,5-фосфоинозитол, как вторичный мессенжер. Контроль ферментной активности через обратимое фосфорилирование белков.
Сера. Цикл серы в природе, соединения, используемые растением. Пути восстановления серы в растении, связь с фотосинтезом. Соединения серы в тканях. Значение сульфидной и дисульфидной связи; серосодержащие ферменты. Роль серы в энергетическом обмене. Значение глутатиона и фитохелатинов.
Микроэлементы.
Железо: доступность в почве, валентность поглощаемой формы, роль микоризы. Особенности поступления железа у двудольных и однодольных растений. Соединения железа; распределение по компартментам клетки и в растении. Комплексы железа в белках редокс-цепей и других ферментах. Железо и синтез хлорофилла. Медь: Содержание и распределение в клетке и тканях. Участие в окислительно-восстановительных процессах дыхания и фотосинтеза. Функции цитозольных оксидаз (аскорбат-, фенол- и диаминоксидаз).
Марганец: Активируемые им ферментные системы, его специфичность, как кофактора. Роль Mn2+ в функционировании ФС-2.
Молибден: Потребность в элементе; его значение для процессов утилизации азота среды. Мо-птерин и функционирование нитрогеназы и нитратредуктазы.
Цинк: Его структурная роль в поддержании ферментной активности и при синтезе белка. Метаболические функции ферментов, содержащих цинк.
Микроэлементы и изоформы супероксиддисмутазы, их локализация в клетке и роль в защите тканей от активных форм кислорода.
Бор: компартментация в клетке; формы соединений. Механизмы участия в регуляции физиологических процессов и метаболизма.
Нарушения в метаболизме растений при недостатке микроэлементов. Токсичность тяжелых металлов для растений их накопление в тканях. Механизмы защиты. Видоспецифичность в чувствительности и устойчивости растений к избытку и недостатку тяжелых металлов в среде. Фиторемедиация.
Регуляция процессов поступления, накопления и распределения элементов минерального питания в системе целого растения
2) Соцветия и их типы, Соцветие – это годичный побег растения (называемый «главная ось»), на котором вырастает группа цветков. Простые соцветия характерны тем, что цветки расположены только на единственной главной оси. Они могут вырастать непосредственно на побеге (в таком случае их называют сидячими), или на веточках различной длины – цветоножках. Всложных соцветиях главная ось ветвится на вторичные побеги (оси), на которых уже образуются соцветия второго порядка — их называют частные, или парциальные (также встречаются частные соцветия третьего и более порядка).
Простые соцветия Сложные соцветия
3) Ботаническое описание Многолетние травы, кустарники или деревца.
Листья большей частью очерёдные, реже супротивные или мутовчатые.
Цветки в соцветиях или одиночные, обоеполые, большей частью неправильные.Чашечка обычно 4—5, реже двураздельная или зубчатая, остающаяся при плодах. Венчик сростнолепестной, колесовидный, колокольчатый, с 4—5-лопастным отгибом или двугубый. Тычинок пять, четыре или две, приросших к трубке венчика. Пыльники двугнёздные или вследствие слияния гнёзд одногнёздные, почковидные. Завязь верхняя, обыкновенно двугнёздная, с семяносцами на перегородке, реже одногнёздная с двумя постенными семяносцами. Столбик простой, с цельным или двулопастным рыльцем.
Формула
цветка: ![]()
![]() .
(реже почти
.
(реже почти ![]() )[1].
Плод —
большей частью коробочка,
раскрывающаяся 2—5 створками или же
путём образования отверстий. Семена обыкновенно
многочисленные, белковые, с прямым зародышем.
)[1].
Плод —
большей частью коробочка,
раскрывающаяся 2—5 створками или же
путём образования отверстий. Семена обыкновенно
многочисленные, белковые, с прямым зародышем.
Подорожниковые (лат. Plantaginaceae) — семейство двудольных растений порядка Ясноткоцветные.Представители семейства — однолетние или многолетние травы, распространённые по всей планете.
Стебли у одних представителей слабо развиты, так что листья, не представляющие большей частью резкого расчленения на пластинку и черешок, бывают собраны в прикорневую розетку; у других
представителей стебли развитые, ветвистые и густо усаженные узкими листьями. Прилистников нет.
Цветки мелкие, не резко заметные, редко одиночные (у Litorella), у большинства же собраны в густые колосья или головки, поднимающиеся на более или менее длинной стрелке над розеткою листьев, у видов же с ветвистым стеблем соцветия занимают верхушку ветвей. Цветок правильного строения, обоеполый, редко (у Litorella, Bougueria) однополый. Околоцветник двойной, состоящий из четырёхраздельной чашечки и четырёхзубчатого венчика; тычинок одна, две или четыре; нити у них очень длинные, выносящие далеко из венчика подвижные пыльники; пестик один с верхнею одно-, дву- или четырёхгнездою завязью и с длинным нитевидным перистым столбиком; в каждом гнезде завязи на центральном семеноносце по нескольку семяпочек.
Плод — орешек (у Litorella, Bougueria) или коробочка, вскрывающаяся поперечною трещиною (у Plantago). Семена мелкие, белковые.
4)орган раст-стебель двудол,покровная ткань-эпидерма,свп-открытые коллатеральные,тканиЦОЦ:запаспаренхима,склеренхима,камбий,флоэма,ксилема
Билет № 10. 1) Флора — исторически сложившаяся совокупность видов растений, распространённых на конкретной территории («флора России») или на территории с определёнными условиями.
Растительность — совокупность фитоценозов определённой территории или всей Земли в целом. В отличие от флоры, которая характеризуется только видовым составом, растительность характеризуется и видовым составом, и численностью особей (причём как в отдельных растительных таксонах, так и в целом для рассматриваемой территории), и особенностями сочетания представителей различных растительных таксонов, и экологическими связями между ними.
Растительность — предмет изучения двух наук: геоботаники и экологии.
Ареа́л (от лат. area: область, площадь, пространство) — область распространения таксона, например, вида. Одно из основных понятий в биологических дисциплинах, изучающих географическое распространение организмов, — географии растений и зоогеографии. Иногда к слову ареал ошибочно добавляется слово обитания (Ареал обитания), что приводит к плеоназму (как, например, сосновый бор).
Ареал вида представляет собой сочетание видовых требований с определённой суммой экологических условий на обширной территории или акватории. Однако эта эколого-географическая сопряжённость видов осложняется изменениями физико-географических условий в прошлом — климата, растительного покрова, формы земной поверхности и пр. Поэтому ареал вида является суммированным эффектом современных и предшествовавших условий. В результате, в географическом распределении видов и их комплексов — флор для растений и фаун для животных — наблюдается ряд неравномерностей и свои закономерности.
Для большинства видов ареал является географически сплошным (то есть таксон встречается на его протяжении более или менее равномерно, без явных обширных пробелов), но у многих видов он разорван (дизъюнктивен), часто из-за глобальных естественных процессов, коренным образом поменявших условия среды в отдельных местах их исходного обитания. Одной из важнейших причин образования разорванных ареалов, например, в Евразии, явились ледниковый период и ксеротермические межледниковые периоды; в результате ряд северных форм оказался занесён далеко на юг, а некоторые южные формы проникли на север. В этих оторванных от сплошного ареала частях такие виды являются реликтами, в приведённом примере — ледниковыми и ксеротермическими. Поэтому с ними связывают особые типы разорванных ареалов:
Рассечённый — ареал, не образующий единой целостности. Характерен, как правило, для реликтовых таксонов
Реликтовый — ареал, имеющий тенденцию к сокращению. Признаком реликтовости является неспособность к возобновлению на территории, ранее занимаемой таксоном, после катастрофического исчезновения (вследствие пожаров и др.)
Нередко эволюция вида к современному периоду завершалась его дифференциацией на пару или несколько очень близких, фенотипически сходных видов, которые при более тщательном изучении оказываются разными. Такие виды-двойники могут быть либо иметь неперекрывающиеся ареалы, то есть быть аллопатрическими, либо могут быть симпатрическими (при частичном пересечении ареалов либо полном их совпадении.
В настоящее время ареалы многих видов изменены в результате их интродукции человеком на новые территории.
Вследствие межвидовой конкуренции ареал, которые вид может занять (аутэкологический) и который занимает фактически (синэкологический), различаются. То же самое можно говорить про экологические ниши — фундаментальную или потенциальную — которую вид может занять в принципе, и фактическую — которую вид занимает при наличии межвидовой конкуренции.
ЭНДЕМИКИ (от греч. endemos - местный) - виды (роды, семейства и т. д.) растений и животных, ограниченные в своем распространении относительно небольшой территорией.
КОСМОПОЛИТЫ - в биологии - виды (роды, семейства и т. п.) растений и животных, обитающие по всему (или почти по всему) земному шару. Космополитами являются, напр., злаки, воробьиные птицы.
2) МЕРИСТЕМЫ ИЛИ ОБРАЗОВАТЕ ЛЬНЫЕ ТКАНИ
Меристемы или образовательные ткани, обладают способностью к делению и образованию новых клеток. За счет меристем формируются все прочие ткани и осуществляется длительный (в течение всей жизни) рост растения. У животных меристемы отсутствуют,. Клетки меристем отличаются высокой метаболической активностью. Одни клетки меристем, получившие название инициальных, задерживаются на эмбриональной стадии развития в течение всей жизни растения, другие постепенно дифференцируются и превращаются в клетки различных постоянных тканей. Инициальная клетка меристемы принципиально может дать начало любой клетке организма. Тело наземных растений — производное относительно немногих инициальных клеток.
Первичные меристемы обладают меристематической активностью, т. е. способны к делению изначально. В ряде случаев способность к активному делению может вновь возникнуть и у клеток, уже почти утративших это свойство. Такие «вновь» возникшие меристемы называют вторичными.
По положению в растении выделяют верхушечные, или апикальные ,боковые, или латеральные и интеркалярные меристемы.
Апикальные меристемы располагаются на верхушках осевых органов растения и обеспечивают рост тела в длину, а латеральные — преимущественно рост в толщину. Каждый побег и корень, а также зародышевый корешок, почечка зародыша имеют апикальную меристему. Апикальные меристемы первичны и образуют конусы нарастания корня и побега (рис. 1).
Латеральные меристемы располагаются параллельно боковым поверхностям осевых органов, образуя своего рода цилиндры, на поперечных срезах имеющие вид колец. Часть из них относится к первичным. Первичными меристемами являются прокамбий и перицикл, вторичными — камбий и феллоген.
Интеркалярные, или вставочные, меристемы чаще первичны и сохраняются в виде отдельных участков в зонах активного роста (например, у оснований междоузлии, в основаниях черешков листьев).
Существуют также раневые меристемы. Они образуются в местах повреждения тканей и органов и дают начало каллюсу — особой ткани, состоящей из однородных паренхимных клеток, прикрывающие место поражения Каллюсо-образовательная способность растений используется в практике садоводства при размножении их черенками и прививками. Чем интенсивнее каллюсообразование, тем больше гарантия срастания подвоя с привоем и укоренения черенков. Образование каллюса— необходимое условие культуры тканей растения на искусственных средах.
Клетки апикальных меристем более или менее изодиаметричны по размерам и многогранны по форме. Межклетников между ними нет, оболочки тонкие, содержат мало целлюлозы. Полость клетки заполнена густой цитоплазмой с относительно крупным ядром, занимающим центральное положение. Вакуоли многочисленные, мелкие, но под световым микроскопом обычно не заметны. Эргастические вещества, как правило, отсутствуют. Пластид и митохондрий мало и они мелки.
Клетки боковых меристем различны по величине и форме. Они примерно соответствуют клеткам тех постоянных тканей, которые из них в дальнейшем возникают. Так, в камбии встречаются как паренхимные, так и прозенхимные инициали. Из паренхимных инициалей образуется паренхима проводящих тканей, а из прозенхимных — проводящие элементы.
3) Водоросли – Thallobionta, или Algae
Большинство из них свободно живут в толще воды (фитопланктон), некоторые прикрепляются ко дну водоемов или подводным предметам, или просто лежат на дне (фитобентос ). Часть водорослей живет на почве, в почве или на стволах деревьев.Водоросли бывают одноклеточными, колониальными или многоклеточными.
Тело водорослей – таллом (слоевище), не дифференцировано на органы и ткани, может быть очень примитивно устроено или сложно расчленено. Ризоиды служат для прикрепления к грунту и лишь у некоторых паразитных форм поглощают питательные вещества. Форма, размер и строение клеток водорослей разнообразны, но все они имеют плотную целлюлозную или пектиновую стенку. Последняя может быть всегда покрыта слизью или покрывается ею в определенные периоды жизни. У большинства водорослей протопласт состоит из цитоплазмы, одного или нескольких ядер и пластид, называемых хроматофорами, имеющими форму лент, зернышек или пластинок и содержащими пигменты разнообразной окраски. У некоторых водорослей в хроматофорах имеются бесцветные округлые тельца пиреноиды, около которых сосредоточивается крахмал.
Размножение:. -Вегетативное размножение-почкованием, делением клетки надвое, обрывками нитей (у многоклеточных нитчатых), распадением колоний и «клубеньками» (харовые водоросли).
-Бесполое спорами: подвижными (зооспорами) или неподвижными (апланоспорами). Споры одноклеточные, образуются в специальных органах – спорангиях или внутри вегетативных клеток.
-Половое размножение, осуществляется гаметами, которые образуются в специальных одноклеточных органах полового размножения: оогониях (женские гаметангии) и антеридиях (мужские гаметангии). Сущность полового размножения заключается в слиянии гамет и образовании зиготы. После периода покоя зигота прорастает, образуя зооспоры или новое растение. В цикле развития водорослей соотношение диплоидной и гаплоидной фаз различно. Если мейоз происходит при прорастании зиготы, то водоросль гаплоидна в течение всей жизни, а диплоидная фаза представлена зиготой. У некоторых редукционное деление происходит в гаметангиях перед образованием гамет, тогда в их жизненном цикле преобладает диплоидная фаза, а гаплоидная представлена только гаметами. Такую смену гаплоидного и диплоидного поколений называют сменой ядерных фаз.
Крупные водоросли используются в пищу, как корм для сельскохозяйственных животных, в медицине служат сырьем для получения агара, альгинатов, йода. Некоторые представители используются в процессе биологической очистки сточных вод и как биоиндикаторы загрязнения водоемов. В зависимости от биохимических особенностей (набор пигментов, состав клеточной стенки, тип запасных веществ) различают следующие отделы водорослей: Красные (Багрянки), Диатомовые, Бурые, Зеленые. Отдел Красные водоросли (багрянки) – Rhodophycota преимущественно обитатели морей, незначительная часть встречается в пресных водоемах и в почве. Среди багрянок есть неподвижные одноклеточные водоросли, нитчатые и пластинчатые. Размеры от нескольких сантиметров до 1-2 м (порфира). Таллом красных водорослей сильно расчленен и имеет вид разветвленных многоклеточных нитей, прикрепленных к субстрату с помощью ризоидов. Хроматофоры в виде зерен или пластинок содержат пигменты: красный (фикоэритрин), синий (фикоциан),зеленый (хлорофилл ). Фикоэритрин преобладает у глубоководных водорослей. Он маскирует другие пигменты и придает ярко-красный цвет. Мелководные формы имеют голубоватую окраску, так как у них преобладают фикоцианины. Продуктом фотосинтеза является полисахарид – багрянковый крахмал , откладывающийся в цитоплазме. Клеточные стенки, кроме целлюлозы, пропитаны большим количеством пектиновых веществ, которые сильно ослизняются и набухают, образуя мягкий или хрящеватый общий каркас, в который как бы погружены протопласты клеток. У некоторых багрянок стенки пропитаны углекислым кальцием и магнием. Размножаются красные водоросли бесполым и половым путем. Подвижные (жгутиковые) стадии при размножении полностью отсутствуют. При бесполом размножении образуются спорангии, содержащие 1-4 пассивные споры. Половой процесс оогамный. Женский половой орган - карпогон имеет брюшко (расширенная базальная часть), в котором находится яйцеклетка и отросток – трихогину. Мужской половой орган антеридий – это мелкая бесцветная клетка, которая несет гаплоидные половые клетки – спермации. Последние, не имея жгутиков, пассивно переносятся токами воды и прилипают к трихогине. В месте контакта наблюдается растворение стенок спермация и трихогины, проникновение ядра спермация в брюшную полость карпогона и слияние с женским ядром. После оплодотворения образуется зигота, а затем диплоидные карпоспоры. Из карпоспор развивается диплоидный таллом, на котором в результате редукционного деления образуются гаплоидные тетраспоры . На гаплоидном талломе, развивающемся из тетраспор, вновь образуются половые органы, и цикл повторяется. Таким образом, жизненный цикл красных водорослей сложен и необычен. В нем чередуются: гаплоидный гаметофит, диплоидный полупаразитический карпоспорофит, сидящий на гаметофите, и диплоидный тетраспорофит, живущий самостоятельно. Спорофиты и гаметофиты имеют сходное или отличное друг от друга строение. Багрянки – лучшие агароносы (анфельция – Ahnfeltia); имеющие сильно ослизняющиеся пектиново-целлюлозные стенки. Порфиру (Porphyra) используют в пищу (в Японии развито ее промышленное культивирование). В некоторых странах багрянки используют на корм сельскохозяйственным животным. Отдел Диатомовые водоросли – Diatomophycota Насчитывает более 10000 видов, распространенных в планктоне морей, океанов и пресных водах, в верхних слоях почвы, в горячих источниках, на снегу. Это одноклеточные и колониальные организмы. Протопласт клеток дифференцирован на цитоплазму, ядро, вакуоли. Запасной продукт углеводной природы – хризоламинарин. Хроматофоры крупные, пластинчатые или зернистые, желтовато-бурые, содержат хлорофиллы, фукоксантин и другие ксантофиллы (диатомовые относятся к группе буроокрашенных водорослей). Вместо клеточной стенки образуется тонкий двустворчатый панцирь, состоящий из кремнезема. Большая створка (эпитека) надета на меньшую створку (гипотека), как крышка на коробку. Под панцирем находится пектиновая стенка. По форме клеток все диатомеи делятся на две группы: центрические (радиально-симметричные) – преимущественно планктонные водоросли морей и океанов и пеннатные (двусторонне-симметричные) – чаще пресноводные бентосные и напочвенные водоросли. Размножаются диатомовые продольным, чаще всего вегетативным делением клеток надвое с последующим достраиванием одной створки кремниевого панциря - гипотеки. Половой процесс – изогамия и оогамия. Диатомовые водоросли живут в диплоидном состоянии (т.е. ядра содержат двойной набор хромосом), и только их гаметы гаплоидны. Роль диатомовых водорослей в природе велика: они служат основным кормом для значительного количества водных организмов (обладают высокой питательной ценностью); принимают первостепенное участие в осадконакоплении (огромное количество панцирей мертвых диатомей образуют на дне водоемов диатомовые илы). Отдел Бурые водоросли – Fucophycota Содержит около 1500 видов, преимущественно растущих во всех морях. Размер этих водорослей от нескольких миллиметров до 20-100 м. Таллом имеет вид либо ветвистых одноядерных и многоядерных нитей, либо у него сложное строение (он как бы расчленен на стеблевидную и листовидную части и имеет зачатки примитивной ткани – ассимиляционную, запасающую, механическую и проводящую) Основными пигментами бурых водорослей являются: хлорофиллы а и с, каротин, ксантофилл. Последний пигмент придает водорослям характерную бурую окраску. Клеточные стенки состоят из двух слоев: наружного ослизняющегося, содержащего альгиновую кислоту, и внутреннего, образованного разновидностью целлюлозы – альгулезой. В клетках имеется одно крупное ядро, крупные вакуоли с клеточным соком и мелкие вакуоли, содержащие дубильные вещества, хроматофоры в виде дисков или зерен. Запасные вещества – ламинарин, маннит и, в небольшом количестве, жиры. Размножение бурых водорослей: вегетативное (частями талломов), бесполое (зооспорами) и половое (изогамия, гетерогамия и оогамия). Представители данного рода (ламинария сахарная - Laminaria saccharina, ламинария пальчатая - L. digitata, ламинария японская – L. japonica) образуют целые плантации в северных и дальневосточных морях (в местах с постоянным движением воды, у открытых берегов). Они также известны под названием «морская капуста». Таллом их расчленен на ризоиды, «стеблевую» и «листовую» части. Внутреннее строение таллома довольно сложно, заметна дифференциация на примитивные ткани. Таллом многолетний, пластина сменяется ежегодно. Описанный таллом – это диплоидный спорофит. На поверхности листовой пластины спорофита группами образуются одногнездные зооспорангии, в них - многочисленные зооспоры. Зооспоры гаплоидны и физиологически неравнозначны. Прорастая, зооспоры дают микроскопического размера гаметофиты - заростки в виде ветвящихся нитей, состоящих из небольшого числа клеток. На заростках образуются органы полового размножения: женские (оогонии) и мужские (антеридии). Антеридии многоклеточные (признак более высокой организации). В гаметангиях формируются по одной гамете (яйцеклетке и сперматозоиду соответственно). После оплодотворения из зиготы без периода покоя вырастает новый диплоидный спорофит. Ламинарии используют: в качестве пищевых продуктов (диетическая пища, богатая йодом); для получения лекарственных препаратов (содержат йод и полисахариды, концентрация йода составляет не менее 0,1 %); для получения полисахарида альгина (применяют в качестве желеобразующих веществ в кондитерской промышленности, при изготовлении лаков, красок); на корм сельскохозяйственным животным; в качестве удобрения (богаты калием). Отдел Зеленые водоросли – Chlorophycota Обитают преимущественно в пресных водах и в морях, некоторые на снеге, стволах деревьев и в почве. У зеленых водорослей представлены все типы организации таллома: одноклеточный, колониальный, многоклеточные нитчатый и пластинчатый Зеленые водоросли содержат пигменты - хлорофиллы «а» и «в», каротины и ксантофиллы. Запасным продуктом является крахмал, который откладывается в хроматофоре вокруг пиреноида. В строме хроматофоров зеленых водорослей находится светочувствительный глазок – стигма. Клеточные стенки обычно содержат целлюлозу и пектины. Эндоплазматическая сеть более развита, чем у остальных водорослей. Представлены все типы размножения и все виды полового процесса. Представители отдела: Род Хламидомонада (Chlamydomonas) из класса Вольвоксовые - включает около 500 наиболее примитивных видов отдела зеленых водорослей. Сюда относятся одноклеточные, округлые или грушевидные, двужгутиковые, имеющие ядро, цитоплазму, хроматофор с пиреноидом; две пульсирующие вакуоли, красный глазок, ориентирующий ее движение под действием света; стенку, состоящую из пектиновых веществ и гемицеллюлозы. Способны к фотосинтезу, но могут потреблять готовые органические вещества. Чаще всего встречаются в загрязненной воде (лужи, канавы). Размножается вегетативно, бесполым и половым путем. Бесполое размножение осуществляется делением материнской клетки на 2, 4, 8 частей, из которых формируются зооспоры, превращающиеся затем в маленькие вегетативные особи. При половом размножении образуются гаметы, при их слиянии образуется зигота, которая зимует, а весной делится и образует молодые особи. Хлорелла (Chlorella) из класса Хлорококковые - очень широко распространенная водоросль, которая встречается в планктоне и бентосе различных водоемов, во влажной почве, на коре деревьев и является обычным симбионтом лишайников. Хлорелла – это одноклеточная водоросль шаровидной формы, лишенная подвижности (пассивно переносятся токами воды). В ее протопласте находится колоколообразный хроматофор с впадиной, содержащей ядро. Размножение только бесполое, с образованием четырех или восьми неподвижных спор. Род Улотрикс (Ulothrix) из класса Улотриксовые включает 25 видов, предпочитающих пресные и солоноватые, чистые водоемы. Улотрикс - зеленая многоклеточная нитчатая водоросль. Нить неветвящаяся, длиной до 10 см. Самая нижняя клетка нити вытянута в длинный бесцветный ризоид (подошва), которым водоросль прикрепляется к субстрату. Остальные клетки одинаковы, каждая имеет один хроматофор с пиреноидами, ядро, тонкую, иногда ослизненную, целлюлозную стенку. Бесполое размножение осуществляется делением клеток на 2 или 4 четырехжгутиковые зооспоры, которые выходят в воду, прикрепляются к подводным предметам, делятся и образуют нить водоросли. При половом размножении (изогамия) образуются мелкие подвижные гаметы, которые выходят в воду и попарно сливаются. Образовавшаяся зигота покрывается плотной оболочкой, при благоприятных условиях она делится на 4 безжгутиковые споры, которые, прикрепляясь к подводным предметам, делятся, образуя нить водоросли. Ульва (Ulva) из класса Улотриксовые широко распространена по побережью южных и северных морей. Ее таллом многоклеточный, пластинчатый; пластинки из двух слоев клеток. У основания пластинка сужается в короткий черешок с подошвой, которой она прикрепляется к твердому субстрату. Пластинка ульвы может легко обрываться и продолжать жить в таком состоянии. Специализированных репродуктивных орагнов нет. Потенциально каждая клетка может стать спорангием у диплоидного поколения, либо гаметангием у гаплоидного. Спирогира (Spirogyra) из класса Конъюгаты - многоклеточная неветвящаяся нитчатая водоросль, образующая в прудах основную массу тины ярко-зеленого цвета. В клетках содержится одно ядро, в центре вакуоль, в пристенном слое цитоплазмы лентовидные, спирально скрученные хроматофоры, по средней линии которых расположены пиреноиды, окруженные зернышками крахмала. Вегетативное размножение - обрывками нитей, бесполое размножение - неподвижными апланоспорами, образовавшимися по одной в каждой клетке. Половое размножение – конъюгация: от супротивных клеток двух расположенных рядом нитей вырастают отростки, которые сливаются (содержимое одной клетки переливается в другую), образуя зиготу, после периода покоя содержимое зиготы дважды делится, образуя 4 клетки, 1 из которых прорастает в новую особь. Каулерпа (Caulerpa) из класса Сифоновые - имеет крупный таллом, одетый толстой оболочкой, клеточных перегородок нет; вакуоль и постенная цитоплазма с многочисленными ядрами непрерывны. Внешне сифоновые имеют сходство с листостебельными растениями. Бесполое размножение зооспорами отсутствует, половое происходит по типу изогамии. Вошерия (Vaucheria) из класса Сифоновые. Эта водоросль живет на влажной почве, в быстротекущих и стоячих водоемах. Некоторые виды этого рода предпочитают солоноватую воду. Слоевище вошерии одноклеточное, многоядерное, в виде тонких зеленых стелющихся нитей. Хлоропласты линзовидной и веретеновидной формы, без пиреноидов. Запасное вещество масло. Размножение обрывками нитей, бесполое – многоядерными многожгутиковыми зооспорами, апланоспорами и цистами. Половой процесс – оогамия. Хара (Chara) из класса Харовые - встречается в пресных и солоноватых водоемах (но сильной солености не переносит). Это крупные, до 50 см и больше, многоклеточные водоросли, предпочитающие чистые, обогащенные солями кальция водоемы. Своим видом хара напоминает высшие растения, в частности хвощи. Так называемый стебель рассечен на так называемые узлы и междоузлия, от узлов отходят «листья» – мутовки коротких боковых ветвей, к субстрату прикрепляется ризоидами. Хроматофоры мелкие, дисковидные. Вегетативное размножение – клубеньками, бесполого размножения нет, половое – оогамия. Половые органы многоклеточные. Род хара включает 40 видов. Представителей харовых иногда выделяют в особый отдел Харовые
ЗНАЧЕНИЕ И ИСПОЛЬЗОВАНИЕ ВОДОРОСЛЕЙ
вырабатывают около 80% всей массы органических веществ, образующихся на Земле Являясь первичными накопителями органического вещества, водоросли прямо или косвенно служат источником пищи для всех водных животных, в том числе и для рыб.
Вред от растительного планктона в рыбном хозяйстве возможен при «цветении» водоемов, следствием которого является замор рыбы. Некоторые виды водорослей (из отделов Cyanophycophyta, Pyrrophycophyta, Chlorophycophyta, Chrysophycophyta) известны своей токсичностью. В период интенсивного размножения они являются причиной появления в воде ядовитых веществ, вызывающих иногда гибель скота, приходящего на водопой. Сильное развитие водорослей способствует загрязнению насосных станций и водопроводов.
Представители отдела Cyanophycophyta играют большую роль в обогащении почв азотом, фиксируя его из атмосферы подобно клубеньковым бактериям. Многие водоросли принимают активное участие в процессе биологической очистки сточных вод. Они могут служить также показателем качества питьевой воды, степени ее загрязнения и пригодности для питья. В приморских районах водоросли используются как ценные удобрения, так как содержат большое количество калийных солей. Ряд водорослей участвует в образовании лечебных грязей.
Некоторые морские макрофиты (например, ламинария) обладают целебными свойствами и употребляются в медицине при болезнях желудочно-кишечного тракта, ревматизме, микседеме, зобе и других заболеваниях, связанных с нарушением функций щитовидной железы. Хондрус используется в народной медицине при легочных заболеваниях, в Англии его применяют при лечении язвы желудка. Кораллина, церамиум, энтероморфа обладают глистогонным действием.
Морские водоросли — наиболее урожайные растения на Земле. Их запасы в Мировом океане исчисляются сотнями миллионов тонн.
Из морских макрофитов получают закрепители для красок, студне- и слизеобразующие вещества. Эти соединения широко используются в пищевой промышленности как заменители желатина, в медицине, при изготовлении красителей, в текстильной промышленности и т. д.
4)А́стровые (лат. Asteráceae), или Сложноцве́тные (лат. Compósitae) Плод сложноцветных — семянка, то есть одногнёздный односемянной, нерастрескивающийся орешек с кожистой или деревянистой оболочкой.
цветкиТрубчатые звезда Са2-3-5~Со(5)А(5)G(2)нижняя
ложноязычковые стрелкаСа0;2;3;5;~Со(7)-~A0G0редко (2)нижняя
Билет № 11. 1) Основными,общими компонентами, из которых построены клетки, являются ядро, цитоплазма с многочисленными органоидами различного строения и функций, оболочка, вакуоль.
Оболочка покрывает клетку снаружи, под ней находится цитоплазма, в ней — ядро и одна или несколько вакуолей
.Оболочка, или клеточная стенка, — это защитное образование. Под оболочкой находится цитоплазма. Важнейшими органоидами цитоплазмы являются митохондрии, эндоплазматический ретикулум (эндоплазматическая сеть), аппарат Гольджи, рибосомы, пластиды, лизосомы.
Аппарат Гольджи особенно развит в выделительных (секреторных) клетках, в которых откладываются или из которых выводятся различные вещества. Он синтезирует и выделяет вещества, образующие клеточную оболочку.
Те части клетки, которые отмирают в процессе ее развития, разрушаются с помощью ферментов лизосом.
Рибосомы представляют собой аппараты для синтеза белка. Пластиды — органоиды, присущие только растительным клеткам.(хлоропласты,лейкопласты,хромопласты)
В типичной растительной клетке имеется крупная вакуоль, наполненная жидким содержимым.Содержимое вакуоли — клеточный сок — это водный раствор очень многих веществ: сахаров, аминокислот, других органических кислот, пигментов (красящих веществ), витаминов, дубильных веществ,
Ядро всегда лежит в цитоплазме. Содержимое ядра — зернистое основное вещество (ядерный сок, или нуклеоплазма), в котором помещаются более плотные структуры — хромосомы и ядрышко.
ФУНКЦИИ:Клетки поглощают питательные вещества, перерабатывают их, добывая энергию и строя собственное тело, дышат, освобождаются от ненужных веществ, борются с различными повреждениями, реагируют на изменения внешних условий, перестраивая свою жизнедеятельность, растут.
Протопласт (от др.-греч. πρῶτος — «первый» + πλαστός — образованный, вылепленный и др.) — содержимое растительной или бактериальной клетки, за исключением внешней клеточной оболочки (клеточной стенки), однако при сохранении клеточной (плазматической) мембраны.
Клеточная оболочка состоит из 2 слоев: внутренний - клеточная стенка (цитоплазматическая мембрана, плазматическая мембрана, клеточная мембрана) и внешнего - у животных (гликокаликс), у растений (клеточная стенка (в состав которой входит целлюлоза)).
Протопласт включает: цитоплазму, ядро, все органеллы клеточную мембрану
То есть протоплазму + мембрану.
Термин протопласт введен А. Ганштейном в 1880 году. Выделение протопластов широко используется в исследованиях клеток и генной инженерии. Жизнеспособные протопласты получают, в частности, в результате обработки клеток специальными ферментами, разрушающими клеточную оболочку.
Это позволяет путем химических и механических методов выделять и исследовать структурные компоненты и даже отдельные молекулы, вносить изменения в генетическую структуру и так далее. Протопласты могут регенировать клеточную оболочку, что используется, в частности, для получения полноценных генетически измененных клеток.
После слияния (соматической гибридизации) протопласты образуют целые живые организмы — регенеранты. Таким путем могут быть получены соматические гибриды растений.
L-формы бактерий и микоплазмы изначально лишены клеточных стенок, по этому признаку их относят к протопластам.
Протопласт – активное живое содержимое клетки. Протопласт представляет собой чрезвычайно сложное образование, дифференцированное на различные компоненты, называемые органеллами (органоидами), которые постоянно в нем встречаются, имеют характерное строение и выполняют специфические функции (рис. 2.1 ). К органеллам клетки относятся ядро, пластиды, митохондрии, рибосомы, эндоплазматическая сеть, аппаратГольджи, лизосомы, микротельца. Органеллы погружены в гиалоплазму, которая обеспечивает их взаимодействие. Гиалоплазма с органеллами, за вычетом ядра, составляет цитоплазму клетки. От клеточной стенки протопласт отделен наружной мембраной – плазмалеммой, от вакуоли - внутренней мембраной –тонопластом. В протопласте осуществляются все основные процессы обмена веществ. В состав протопласта входят и неорганические вещества в виде ионов минеральных солей (2-6%). Белки, нуклеиновые кислоты, липиды и углеводы синтезируются самим протопластом. -в клетке присутствуют запасные вещества Цитоплазма - происходят все процессы клеточного обмена, кроме синтеза нуклеиновых кислот. Основу цитоплазмы составляет ее матрикс, или гиалоплазма, в который погружены органеллы. Гиалоплазма – сложная бесцветная, оптически прозрачная коллоидная система, она связывает все погруженные в нее органеллы, обеспечивая их взаимодействие. Гиалоплазма содержит ферменты и активно участвует в клеточном метаболизме, в ней протекают такие биохимические процессы, как гликолиз, синтез аминокислот, синтез жирных кислот и масел и др. Она способна к активному движению и участвует во внутриклеточном транспорте веществ. Часть структурных белковых компонентов гиалоплазмы формирует надмолекулярные агрегаты со строго упорядоченным расположением молекул - микротрубочки и микрофиламенты. Микротрубочки – это тонкие цилиндрические структуры диаметром около 24 нм и длиной до нескольких микрометров. Их стенка состоит из спирально расположенных сферических субъединиц белка тубулина. Микротрубочки участвуют в ориентации образуемых плазмалеммой целлюлозных микрофибрилл клеточной стенки, во внутриклеточном транспорте, поддержании формы протопласта. Из них образуются нити веретена деления во время митоза, жгутики и реснички. Микрофиламенты представляют собой длинные нити толщиной 5-7 нм, состоящие из сократительного белка актина. В гиалоплазме они образуют пучки – цитоплазматические волокна, или принимают вид трехмерной сети, прикрепляясь к плазмалемме, пластидам, элементам эндоплазматической сети, рибосомам, микротрубочкам. Считается, что, сокращаясь, микрофиламенты генерируют движение гиалоплазмы и направленное перемещение прикрепленных к ним органелл. Совокупность микротрубочек и микрофиламентов составляет цитоскелет.
Локализация ДНК и РНК в клетке различна. ДНК локализована в ядре клетки, РНК – в ядре и цитоплазме. Только тРНК находится преимущественно к цитоплазме в растворенном виде. Как правило, нуклеиновые кислоты (ДНК и РНК) в живой клетке находятся в форме нуклеопротеинов. Основные структуры, содержащие нуклеопротеины, - это хроматин, где локализована ДНК, и рибосомы, где локализована РНК. В клетке встречаются также полирибосомы – образования, состоящие из вытянутой молекулы мРНК, окруженной глобулярными белками, на которую нанизаны рибосомы.
2) Развитие плода
Как правило, плод развивается после оплодотворения, но у части покрытосеменных может образовываться и в результате апомиксиса, то есть развитие зародыша семени без оплодотворения (партенокарпические плоды).
Морфологической основой плода является гинецей, прежде всего завязь. Прочие части цветка — околоцветник, тычинки и чашечка — чаще быстро увядают, но нередко изменяются и вместе с гинецеем также принимают участие в формировании плода, становясь сочными или, напротив, деревянистыми или пленчатыми. Самые глубокие изменения происходят в завязи. Её стенки разрастаются за счёт усиленного деления клеток и увеличения их размеров. После опыления происходит существенное изменение направленности транспортных потоков и перераспределение питательных веществ в сторону развивающихся плодов. У травянистых растений (особенно у однолетних) практически все синтезируемые органические вещества используются развивающимися семенами и плодами, что ведёт к истощению других тканей растения. Многие растения плодоносят только раз за всю свою жизнь.
Созревание начинается с того, что плод прекращает рост, разлагаются хлорофилл и дубильные вещества, в вакуолях накапливаютсяпигменты, определяющие характерную для данного вида окраску плодов. У некоторых видов растений, например картофеля, плоды даже при созревании остаются зелёными. В стенках плода сосредотачиваются различные пластические и энергетические вещества:белки, крахмал, сахара, жирные масла, некоторые витамины и т. п. Зрелый плод характеризуется совокупностью только ему присущих особенностей. Плод несёт семя или семена, которых у ряда растений может быть до нескольких тысяч. Это важнейшая его часть, обеспечивающая воспроизведение данного вида в ряду поколений. Однако, по массе семена, как правило, относятся к меньшей части плода. В природе и особенно в культуре встречаются бессемяные плоды. В результате длительной селекции выведены бессемянные сорта винограда (Vitis), банана (Musa) и т. д.
Зрелые семена прикрепляются к околоплоднику в тех местах, где в завязи располагалась плацента, либо свободно лежат в полости плода, либо плотно окружены мясистой стенкой. Максимальное число семян в плоде равно числу семязачатков, но обычно меньше, так как не все семязачатки достигают зрелости.
Созревший плод переходит в последнюю стадию — отмирания, в ходе которой в плод обычно не поступают новые вещества, не делятся и не растут клетки, и постепенно ткани плода разрушаются и сгнивают. У большинства цветковых растений созревший плод опадает и отмирает уже на грунте.
[править]Строение плода
В образовании плода принимают участие различные части цветка, но прежде всего завязь. Из стенки завязи формируется стенка плода — околоплодник или перикарпий. В перикарпии обычно различают три слоя: наружный — экзокарпий или эпикарпий, средний —мезокарпий и внутренний — эндокарпий. Семена формируются из семяпочек.
Все три слоя, обычно, хорошо различимы. Например, в плоде вишни тонкий кожистый наружный слой — экзокарпий, съедобная сочная мякоть плода — мезокарпий, твёрдая косточка из каменистой ткани, окружающая единственное семя — эндокарпий. Нередко эти слои околоплодника различаются слабо, даже при анатомическом исследовании, что связано с деформацией и сдавливанием клеток при созревании плода.
[править]Классификация плодов
В большинстве классификаций плоды обычно разделяют на настоящие или истинные (формирующиеся из разросшейся завязи) иложные (в их образовании принимают участие и другие органы). Настоящие плоды подразделяют на простые (сформированные из одного пестика) и сворные, сложные (возникшие из многочленного апокарпного гинецея). Пример сворных плодов: сложный орешекили многоорешек (шиповник), сложная семянка (клубника, земляника), сложная костянка (малина), фрага или земляничина(многоорешек на разросшемся при созревании мясистом цветоложе). Простые плоды делят по консистенции околоплодника на сухие исочные.
I. Сухие — с сухим околоплодником:
1. Коробочковидные — многосеменные: собственно коробочка (мак, тюльпан, дурман); крыночка; боб (Семейство Бобовые); мешочек; стручок или стручочек (Семейство Крестоцветные); листовка.
2. Ореховидные или односеменныеорех, орешек (лещина, фундук);зерновка (злаки);крылатка (клён);жёлудь (дуб);семянка.
II. Сочные — с сочным околоплодником:
1. Ягодовидные — многосеменные:ягода (плод черники, смородины, томата);яблоко (плоды яблони, груши, рябина);тыквина (плоды арбуза, тыквы, кабачка);гесперидий, или померанец (плод цитрусовых);гранатина (плод граната).
2. Костянковидные:сочная костянка (вишня, слива, персики);сухая костянка (грецкий орех).
Сложные плоды называют, исходя из названий простых плодов (многолистовка, многокостянка, многоорешек и т. д.).
3)Водоросли
Это низшие хлорофиллсодержащие растения, не расчлененные на стебель, корень и листья. Обитают преимущественно в пресных водоемах и морях.
Отдел зеленые водоросли Зеленые водоросли делятся на одноклеточные и многоклеточные формы, содержат хлорофилл. У них встречаются все виды бесполого и полового размножения. Зеленые водоросли встречаются в соленых и пресных водоемах, в почве, на коре деревьев, на камнях и скалах. Этот отдел насчитывает до 20 тыс. видов и включает пять классов:
Класс вольвоксовые - наиболее примитивные одноклеточные водоросли со жгутиками. Некоторые их виды представляют собой колонию
.Класс протококковые - одноклеточные и многоклеточные бейжгутиковые формы.
Класс улотриксовые - имеют нитчатое или пластинчатое строение слоевища.
Класс жаровые — строением напоминают высшие растения - хвощи.
Класс сифоновые — внешне похожи на другие водоросли или на высшие растения, состоят из одной многоядерной клетки, достигая размеров до 1 м.
Одноклеточная зеленая пресноводная водоросль - хламидомонада. Имеет овальную или круглую форму тела, на вытянутом переднем конце два жгутика. Хроматофор чашевидный, с пиреноидом, содержащим зерна крахмала. В передней части клетки красный глазок - это светочувствительный орган. Ядро одно, с маленьким ядрышком. Две пульсирующие вакуоли смещены к переднему концу клетки. Хламидомонада питается автотрофно, но при отсутствии света может перейти на гетеротрофное питание, если в воде присутствуют органические вещества. Размножается бесполым и половым путем. При бесполом размножении содержимое клетки (спорофит) делится на 4 части и образуются 4 гаплоидных зооспоры. С наступлением холодов 2 зооспоры сливаются, образуя диплоидную зиготоспору. Весной она делится митозом, вновь образуя гаплоидные водоросли.
Спирогира - пресноводная зеленая многоклеточная нитчатая водоросль. Нити составлены одним рядом одноядерных цилиндрических клеток со спиралевидными хлоропластами и пиреноидами. Рост нити в длину происходит бесполо за счет поперечного деления клеток. Размножается частями нити или половым путем. Половой процесс называется конъюгацией.
Отдел бурые водоросли
Многоклеточные морские водоросли. Насчитывается ок. 1500 видов. Имеют желтовато-бурую окраску, обусловленную большим количеством желтых и бурых пигментов. Размер и форма их различны. Встречаются нитевидные, корковидные, шаровидные, пластинчатые и кустообразные растения. Слоевища (тела) многих видов содержат газовые пузырьки, удерживающие водоросли в вертикальном положении. Вегетативное тело расчленено на подошву или ризоиды, служащие органами прикрепления, и на простую или рассеченную пластину, соединяющуюся с подошвой черешком. Пигменты, придающие им бурый цвет, сосредоточены только в поверхностных слоях клеток, внутренние клетки талома бесцветны. Это свидетельствует о дифференциации клеток по функциям: фотосимтетической и загасающей. У бурых водорослей нет настоящей проводящей системы, однако, в центре слоевища имеются ткани, по которым передвигаются продукты ассимиляции. Всасывание минеральных веществ осуществляется всей поверхностью слоевища.
У бурых водорослейьвстречаются все формы размножения: вегетативное (при случайных отделениях частей слоевища), споровое, половое (три формы: изогамная, гетерогамная и моногамная).
Отдел красные водоросли (багрянки)
Встречаются обычно на больших глубинах теплых морей. Насчитывают ок. 4000 видов. Имеют расчлененное слоевище, ризоидом или подошвой крепятся к субстрату. Помимо обычных хлорофиллов и каратиноидов, в пластидах багрянок содержатся фикобилины. Другая их особенность - сложный половой процесс. Гаметы и споры красных водорослей лишены жгутиков и неподвижны. Оплодотворение происходит при пассивном переносе мужских половых клеток к женскому половому органу:
Значение водорослей
Водоросли - первичные продуценты с высокой продуктивностью. С них начинаются большинство пищевых цепей морей, океанов и пресных водоемов Одноклеточные водоросли являются главным компонентом фитопланктона, который служит кормом многим видам водных животных. Водоросли обогащают атмосферу кислородом.
Из водорослей получают много ценных продуктов. Например, из красных водорослей получают полисахариды агар-агар и карраген (используются для получения желе, в косметике и как пищевые добавки); из бурых водорослей получают альгиновые кислоты (применяют в качестве отвердителей, желеобразующих веществ в пищевой, косметической промышленности, для изготовления красок и паков).
4)это корневище однодольного так как середина объекта занята паренхимой, в ЦОЦ беспорядочно распложены концентрические центроксилемные пучки.покровная ткань эпидерма в состав первичной коры входит запасающая паренхима.
Билет № 12. 1)Клеточная стенка (оболочка) является неотъемлемым компонентом клеток растений и грибов и представляет собой продукт их жизнедеятельности. Она придает клеткам механическую прочность, защищает их содержимое от повреждений и избыточной потери воды, поддерживает форму клеток и их размер, а также препятствует разрыву клеток в гипотонической среде. Клеточная стенка участвует в поглощении и обмене различных ионов, т. е. является ионообменником. Через клеточную оболочку осуществляется транспорт веществ.
Клеточная стенка, формирующаяся во время деления клеток и их роста путем растяжения, называется первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои, и образуется прочная вторичная клеточная оболочка.
В состав клеточной стенки входят структурные компоненты (целлюлоза у растений и хитин у грибов), компоненты матрикса (гемицеллюлоза, пектин, белки), инкрустирующие компоненты (лигнин, суберин) и вещества, откладывающиеся на поверхности оболочки (кутин и воск).
Молекулы целлюлозы за счет водородных связей объединяются в пучки —микрофибриллы. Переплетенные микрофибриллы составляют каркас клеточной оболочки. У большинства грибов микрофибриллы клеточной стенки состоят изхитина.
Микрофибриллы погружены в матрикс клеточной стенки. Матрикс состоит из смеси различных химических веществ, среди которых преобладают полисахариды (гемицеллюлозы и пектиновые вещества).
Гемицеллюлозы — это группа полисахаридов (полимеры пен-тоз и гексоз — ксилозы, галактозы, маннозы, глюкозы и др.). Молекулы гемицеллюлоз, как и целлюлозы, имеют форму цепи, но в отличие от последней их цепи короче, менее упорядочены и сильно разветвлены. Они легче растворяются и разрушаются ферментами.
Пектиновые вещества — это полимеры, построенные из моносахаридов (арабинозы и галактозы), галактуроновой кислоты (сахарной кислоты) и метилового спирта. Длинные молекулы пектиновых веществ могут быть линейны ми или разветвленными. Молекулы пектиновых веществ содержат большое количество карбоксильных групп и поэтому способны соединяться с ионами Mg2+ и Са2-. При этом образуются клейкие, студнеобразные пектаты магния и кальция, из которых затем складываются срединные пластинки, скрепляющие стенки двух соседних клеток.
Ионы двухвалентных металлов могут обмениваться на другие катионы (Н-, К+ и т. д.). Это обусловливает катионообменную способность клеточных оболочек.
Пектиновыми веществами и пектатами богаты оболочки клеток многих плодов. Так как при их извлечении из оболочек и добавлении сахара образуются гели, пектины используют как желе-образующие вещества для изготовления мармелада и др.
Помимо углеводных компонентов, в состав матрикса клеточной стенки входит структурный белок экстенеин —гликонроте-ин, который по своему составу близок к межклеточным белкам животных —коллагенам.
На долю матрикса приходится до 60% сухого вещества клеточной оболочки. Матрикс оболочки не просто заполняет промежутки между микрофибриллами, а образует прочные химические (водородные и ковалентные) связи между макромолекулами и микрофибриллами, что обеспечивает прочность клеточной стенки, ее эластичность и пластичность.
Основным инкрустирующим веществом оболочки клеток растений являетсялигнин — полимер с неразветвленной молекулой, состоящей из ароматических спиртов.
Интенсивная лигнификация (пропитка слоев целлюлозы лигнином) клеточных оболочек начинается после прекращения роста клетки. Лигнин может откладываться отдельными участками — в виде колец, спиралей или сетки, как это наблюдается в оболочках клеток проводящей ткани — ксилемы, или сплошным слоем, за исключением тех мест, где осуществляются контакты между соседними клетками в виде плазмодесм.
Лигнин скрепляет целлюлозные волокна и действует как очень твердый и жесткий каркас, усиливающий прочность клеточных стенок на растяжение и сжатие. Он же обеспечивает клеткам дополнительную защиту от физических и химических воздействий, снижает водопроницаемость. Содержание лигнина в оболочке достигает 30%. Инкрустация им клеточных оболочек приводит к их одревеснению,которое часто влечет за собой отмирание живого содержимого клетки.
Лигнин в сочетании с целлюлозой придает особые свойства древесине, которые делают ее незаменимым строительным материалом.
На клеточную оболочку могут откладываться также жиропо-добные вещества — суберин, кутин и воск.
Суберин откладывается на оболочку изнутри и делает ее практически непроницаемой для воды и растворов. В результате протопласт клетки отмирает и клетка заполняется воздухом. Такой процесс называется опробковением.Наблюдается опробковение оболочки клеток в покровных тканях многолетних древесных растений — перидерме, корке, а также в эндодерме корня.
Поверхность эпидермальных клеток растений защищена гидрофобными веществами — кутином и восками. Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами (целлюлозой и пектином) и образует кутикулу. Воск часто откладывается в кристаллической форме на поверхности частей растений (листьев, плодов), образуя восковой налет.
Кутикула и восковой налет защищают клетки от повреждений и проникновения инфекции, уменьшают испарение воды с поверхности органов.
В оболочках эпидермальных клеток некоторых растений (злаков, осок и др.) накапливается большое количество минеральных веществ (минерализация), в первую очередь карбоната кальция и кремнезема. При минерализации листья и стебли растений становятся жесткими, твердыми и в меньшей степени поедаются животными.
Таким образом, клеточная стенка играет важную роль в жизни клеток растений и грибов и выполняет ряд специфических функций.
2) Вторичное строение корня характерно только для двудольных покрытосеменных и голосеменных растений, т. е. для растений, имеющих вторичные меристемы — камбий и феллоген. В формировании вторичного строения участвует только центральный цилиндр, а первичная кора и ризодерма отмирают и слущиваются. Центральный же цилиндр разрастается в ширину за счет работы камбия, который закладывается из прокамбия, оставшегося между ксилемой и флоэмой радиального проводящего пучка. Снаружи корень покрывается пробкой, образовавшейся из феллогена перициклического происхождения. За исключением первичной ксилемы проводящего пучка, расположенной в центре органа, все ткани корня вторичного строения имеют вторичное происхождение. Морфолого-анатомические особенности строения корневищ однодольных и двудольных Корневища однодольных имеют разбросанно-пучко-вый тип строения; центральный осевой цилиндр отделен от первичной коры эндодермой; проводящие пучки расположены без какого-либо порядка в большем числе в центральном осевом цилиндре и в меньшем числе в коре; поверхность коры покрыта опробковевшей тканью. 2. Корневища двудольных имеют кольцевое, пучковое или беспучковое строение. Проводящие пучки расположены в основной ткани, по кругу к периферии первичная кора, в центре широкая сердцевина; в корневищах с беспучковым строением древесина (ксилема) расположена широким поясом, отделенным от коры кольцом камбия, во вторичной коре и в древесине радиально расположены сердцевинные лучи, в центре - сердцевина. Покровная ткань - пробка. Цвет корней и корневищ определяют снаружи и на свежем изломе (на воздухе поверхность излома темнеет), запах - при измельчении. Вкус определяют только у неядовитых объектов сухого сырья 3) СЕМЕЙСТВО ОРХИДНЫЕ (ORCHIDIACAEA)
Около 800 родов и 30 000 видов. Распространены на всех континентах, в разных растительных зонах и поясах, за исключением полярных областей и пустынь. Наиболее представлены во влажных тропических лесах южной и юго-восточной Азии и Америки. Это наиболее высокоспециализированная в направлении энтомофилии группа однодольных растений. Это многолетние травы, автотрофные, иногда сапрофиты. В тропиках - часто эпифиты. Все орхидные образуют микоризу.
Признаки семейства:
Листья очередные, редко супротивные или мутовчатые, цельные, влагалищные
Цветки разнообразной формы и окраски, образуют соцветия (колосья, кисти , метелки) или одиночные, обоеполые, реже однополые, зигоморфные
Тычинки в числе 1-2
Гинецей паракарпный, из 3 плодолистиков.
Завязь - нижняя
Плод - коробочка
Отдел покрытосеменные (цветковые) - Angiospermae, Anthophyta, Magnoliophyta Класс однодольные (лилиопсиды) – Monocotyledones (Liliorsida) Подкласс лилииды - Liliidae Порядок орхидные – Orchidales Семейство орхидные – Orchidales
*(классификация приведена по книге «Жизнь растений» т. 6, М., Просвещение, 1982 год)
Одно из крупнейших семейств цветковых растений, в его состав входят около 20 000 видов. Большинство из них произрастает в тропических странах. На территории бывшего СССР распространено 57 родов и 153 вида представителей этого семейства. Орхидные космополиты. Они встречаются почти во всех пригодных для обитания растений областях земли, от Швеции и Аляски на севере до Огненной земли и субарктического острова Маккуори на юге. Большинство представителей семейства сосредоточены в тропических широтах, особенно в Америке и юго-восточной Азии. Растут орхидные повсюду: от склонов самых высоких гор до лесных просторов равнин, от болот и водоемов до сухих степей и пустынных оазисов. В Колумбии некоторые орхидные растут в нетающих снегах гор, а в Австралии под землёй.
Орхидные многолетние травянистые растения: наземные в умеренных широтах и преимущественно эпифитные – в тропических; с корневищами, подземными или надземными клубнями. Листья очередные, реже супротивные или мутовчатые, цельнокрайние, с дуговидным или параллельным жилкованием, иногда влагалищные, обычно слегка мясистые. Большинство наземных и эпифитных орхидей являются автотрофными растениями с зелёными листьями в которых осуществляется фотосинтез. У некоторых эпифитных орхидей функцией фотосинтеза наделены также зелёные стебли, цветки и нередко воздушные корни. Некоторые виды лишены зелёной окраски и питаются за счет симбиоза с грибами, являясь сапрофитами, а по мнению некоторых исследователей даже паразитами. Цветки одиночные или в соцветиях, обоеполые, неправильные. С простым венчиковидным, часто яркоокрашенным околоцветником, состоящим из двух трехчленных кругов. Один листок внутреннего круга обычно резко отличается от других формой, большими размерами и окраской и превращен в так называемую губу. Чаще развиты одна (две) тычинки, остальные превратились в бесплодные тычинки-стаминодии. Тычинки обычно срастаются со столбиком, образуя колонку. Завязь нижняя, обычно одногнездная. Пальца обычно не рассыпается на отдельные пылинки, а склеена в один комочек – поллиний, имеющий ножку с липким кончиком (все это вместе называется поллинарием). Насекомое опылитель садиться на губу, всовывает голову в цветок и к ней прилипают мелкие подушечки поллинариев. При посещении следующего цветка поллинарии на голове насекомого приходятся как раз против рыльца. Этот оригинальный способ опыления был изучен ещё Ч. Дарвином и подробно описан в большом количестве книг. Плод коробочка раскрывающаяся шестью продольными щелями, семена очень мелкие, многочисленные, пылевидные. Особенностью семейства, является обязательное участие гриба в прорастании семени и присутствии эндотрофной микоризы в течение всей или большей части жизни до цветения. Цветение у некоторых видов наступает на 15-20 год после прорастания семени.
У большинства орхидных цветы очень красивы, что служит причиной их массового истребления. Многие виды занесены в Красную книгу. 4) Это семейство бурачниковые (Boraginaceae). Формула: *Са(5)Со(5)А5G(2)_- окопник лекарственный ( Symphytum officinale). Плод – ценокарпий(четырехорешек)
Билет 13 Вопрос 1. Клеточная стенка. Жесткая клеточная стенка(оболочка), окружающая растительную клетку, состоит из целлюлозных микрофибрилл, погруженных в матрикс, в состав которого входят другие сложные полисахариды – гемицеллюлозы и пектиновые вещества. Клеточная стенка обеспечивает сохранение формы клетки, её механическую опору и защиту протопласта. Клеточная стенка способна к растяжению. Стенка может расти только в контакте с протопластом. Через клеточную стенку происходит передвижение воды и минеральных солей, но для высокомолекулярных веществ она полностью или частично непроницаема. При отмирании протопласта стенка может продолжать выполнять функцию проведения воды. Наличие клеточной стенки больше всех других признаков отличает растительную клетку от животной. Она играет существенную роль в поглощении, транспорте и выделении веществ. Целлюлозный каркас клеточной стенки заполнен нецеллюлозными молекулами матрикса. В состав матрикса входят: полисахариды, называемые гемицеллюлозами; пектиновые вещества(пектин) и гликопротеиды. Пектиновые вещества, сливаясь между соседними клетками, образуют срединную пластинку. При растворении или разрушении срединной пластинки( в мякоти созревших плодов) происходит процесс называемый мицерацией. Клеточная стенка в процессе мицерации может подвергаться различным видоизменениям – одревеснению, опробковению, ослизнению, кутинизации и минерализации. Одревеснение связано с внедрением между молекулами целлюлозы лигнина( увеличивает жесткость оболочки). Обычно содержится в клетках выполняющих опорную или механическую функцию. Реактив HCl + флороглюцин на лигнин дает красно-малиновое окрашивание. Кутин, суберин и воск – жироподобные вещества. Кутин и воск обычно откладываются на поверхность клеток эпидермы. Кутиновая пленка образует кутикулу. Суберин пропитывает клеточные стенки вторичной покровной ткани, вызывая опробковение. Клеточные стенки могут пропитываться оксалатом кальция и кремнеземом, что придает им твердость и хрупкость и приводит к минерализации. Клеточные стенки кожуры семян(льна) способны к ослизнению. Это происходит за счет превращения целлюлозы и пектина в слизи и камеди, обеспечивая защиту от высыхания.
Вопрос 2.Метаморфозы побега, анатомические различия надземного стебля и корневища. Подземные видоизменения побегов: 1)корневище(папоротник, ландыш) – многолетний подземный побег, имеющий редуцированные листья в виде бесцветных или бурых мелких чешуек, в пазухе которых лежат почки. 2) кубень(картофель) – метаморфоз побега с ярко выраженной запасающей функцией стебля, наличием чешуевидных листьев, которые быстро сшелушиваются, и почек, формирующихся в пазухах листьев и называемых глазками. Столоны-однолетние подземные недолговечные корневища, на которых образуются клубни. 3)луковица – укороченный побег, стеблевая часть которого называется донцем. Различают два типа видоизмененных листьев: 1-листья с чешуевидными сочными основаниями, запасающими воду с растворенными в ней питательными веществами, 2-сухие листья, покрывающие луковицу снаружи и выполняющие защитную функцию. 4) клубнелуковица(гладиолус) – видоизмененная луковица с разросшимся донцем, образующим клубень, покрытый основаниями зеленых листьев. Надземные видоизменения побегов: 1) колючки выполняют главным образом защитную функцию. Могут образовываться за счет превращения бокового побега в острие – колючку. 2) усики характерны для растений, которые могут поддерживать вертикальное положение и образуются в пазухе листа. Служит органом прикрепления. 3) Кладодии и филлокладии – видоизмененные побеги, выполняющие функцию листьев. Кладодии – боковые побеги, сохраняющие способность к длительному росту, находящиеся на зеленых плоских длинных стеблях (опунция). Филлокладии – уплощенные боковые побеги, имеющие ограниченный рост, так как верхушечная меристема быстро дифференцируется в постоянные ткани. Для стеблей однодольных растений характерны: 1) первичное строение на протяжении всей жизни, 2) слабо выражена первичная кора, 3) разбросанное положение сосудисто-волокнистых пучков, 4) закрытые коллатеральные пучки(без камбия), 5) наличие во флоэмк только ситовидных трубок с клетками-спутницами, 6) отсутствие сердцевины. Особенности строения корневищ однодольных: 1) покровная ткань – эпидерма(часто одревесневшая), 2) первичная кора > ЦОЦ и представлена запасающей паренхимой, 3) во внутреннем слое первичной коры формируется однослойная эндодерма( подковообразная или с пятнами Каспари), 4) ЦОЦ начинается с живого перицикла, 5) закрытые коллатеральные или концентрические пучки, расположенные беспорядочно в ЦОЦ. Особенности строения стебля двудольных: 1) рост в толщину, 2) хорошо дифференцированная первичная кора (колленхима, хлорофиллоносная паренхима, крахмалоносная эндодерма), 3) коллатеральные(биколлатеральные) открытые(с камбием) пучки, 4) непучковое строение( пучки по кольцу или сливаются), 5) есть сердцевина, 6) наличие в ксилеме годичных колец(для древесных). Особенности строения корневищ двудольных: 1) покровная ткань – эпидерма или перидерма(у многолетних), 2) первичная кора – запасающая паренхима и эндодерма с пятнами Каспари, 3) первичная кора примерно=ЦОЦ, 4) ЦОЦ, пучки те же, что и у наземных стеблей.
Вопрос 3. Подкласс розиды, порядок аралиецветные, семейства аралиевые, зонтичные… Подкласс розиды(Rosidae), порядок зонтичные(сельдерейные)(Apiales или Umbelliferae), семейство аралиевые (Araliaceae). Обитает в тропиках и субтропиках. Жизненная форма – небольшие деревья или кустарники, реже многолетние травы и лианы. Листорасположение обычно очередное. Листья простые цельные, пальчато-или перистосложные, с влагалищами. Цветки правильные, обоеполые, или раздельнополые у двудомных растений. Околоцветник двойной, 5- или 4-членный. Чашечка из 4 или 5 чашелистиков, прирастающих к завязи. Лепестков 5(иногда меньше), свободных или частично срастающихся. Количество тычинок = числу лепестков, чередующиеся с ними и прикрепленные к краям особого железистого диска, окружающего верхушку завязи. Гинецей ценокарпный, состоит из 2-5 сросшихся плодолистиков. Завязь нижняя двух- и пятигнездная. Соцветия – кисти и метелки из простых зонтиков, плод – ценокарпная костянка. Представители – 1) плющ обыкновенный (Hedera helix - *Са(5)Со5А5G(5)) , 2) женьшень ( Panax ginseng - *Са(5)Со5А5G(2)). Женьшень – корень жизни. Семейство зонтичные или сельдерейные( Apiales или Umbelliferae). Обычно обитают в тропиках, субтропиках. Жизненная форма – многолетние травы, редко кустарники. Цветок - *Са5Со5А5G(2). Листорасположение очередное. Листья простые, обычно сильнорассеченные(3, 4), редко-цельные, цельнокрайние без прилистников, основания образуют влагалища. Цветки правильные, обоеполые, растения однодомные, реже двудомные. Околоцветник двойной, пятичленный. Чашечка состоит из 5 зубцов, венчик из 5 свободных лепестков. Тычинок всегда 5, чередующихся с лепестками, прикрепленных к железистому диску. Гинецей ценокарпный, сросшийся из 2 плодолистиков, образующих нижнюю двугнездную завязь. На верхушки завязи есть подстолбие. Соцветие – сложный зонтик. Плод – ценокарпий: вислоплодник. Представители: 1) укроп ( Anethum graveolens), морковь ( Daucus carota), цикута (Cicuta virosa), сельдерей ( Apium graveolens), фенхель ( Foeniculum vulgare)...В медицине используют, как отхаркивающее, противовоспалительное и желчегонное средство.
Вопрос 4. Т. Это семейство диоскорейные(Dioscoreaceae). Цветки раздельнополые: 1)женские - *P(3+3)A0G(3), 2)мужские - *P(3+3)A3+3G0. Плод – ценокарпии: крылатые вскрывающиеся коробочки.
Билет 14 Вопрос 1. Физиологические функции корня. Корень как орган превращения и поглощения веществ. Опыты Шмука. Корень – осевой орган цилиндрической формы, обладающий радиальной симметрией и положительным геотропизмом.Функции: 1)минеральное и водное питание, 2) закрепление растения в почве(заяковыряние), 3) синтез продуктов первичного и вторичного метаболизма, 4)накопление запасных веществ, 5)вегетативное размножение, 6)симбиоз с бактериями, 7) функция дыхательного органа( монстера, филодендрон…). Зона всасывания ( от нескольких мм до см) характеризуется наличием корневых волосков, являющихся выростами клеток ризодермы. Корневые волоски увеличивают всасывающую поверхность корня и обеспечивает активное всасывание воды и растворов солей, но они не долговечны. Новые корневые волоски образуются под зоной всасывания и отмирают над зоной всасывания. По мере роста растения зона всасывания перемещается и растение имеет возможность поглощать минеральные вещества из разных слоев почвы. Очень интересные опыты проведены акад. А. А. Шмуком (1941, 1945 гг.). Оказалось, что в табаке, привитом на томате, не образуется никотина; если же, наоборот, томат привить на табак, то в листьях томатов накапливается несвойственный ему никотин (до 4%). Следовательно, никотин образуется не в листьях, а в корнях. Корни выделяют в наружную среду сахара, органические кислоты, соли фосфора, калия и др. Корневая система во взаимосвязи с листьями регулирует окислительно - восстановительные процессы в растении. Вопрос 2. Способы утолщения стеблей однодольных и двудольных растений. Анатомическое строение этих стеблей. Согласно теории туники и корпуса в конусе нарастания стебля покрытосеменных выделяются две зоны: наружная – туника и внутренняя – корпус. Слои туники образуют все ткани первичной коры. Развитие стебля осуществляется за счет дифференциации клеток туники и корпуса – первичных меристем. Из них образуется первичная покровная ткань – эпидерма, первичная кора и ЦОЦ. Первичное строение стебля формируется за счет деятельности первичных меристем апекса. С поверхности стебель однодольных покрыт однослойной эпидермой, которая впоследствии покрывается кутикулой. Под эпидермой располагается первичная кора. Она представлена однородными клетками хлорофиллоносной паренхимы, граничащей со склеренхимой перециклического происхождения ЦОЦ. Она придает прочность растению. Вторичное утолщение стеблей древесных однодольных растений осуществляется за счет кольца утолщения, которое представляет собой особый валик вокруг конуса нарастания, дающий дополнительный ряд сосудисто-волокнистых пучков. . Для стеблей однодольных растений характерны: 1) первичное строение на протяжении всей жизни, 2) слабо выражена первичная кора, 3) разбросанное положение сосудисто-волокнистых пучков, 4) закрытые коллатеральные пучки(без камбия), 5) наличие во флоэмк только ситовидных трубок с клетками-спутницами, 6) отсутствие сердцевины. Двудольное растение. Пучковое строение стебля. Прокамбинальные тяжи закладываются в конусе нарастания в один круг по переферии ЦОЦ. Каждый прокамбиалный тяж превращается в коллатеральный пучок, состоящий из первичной флоэмы и первичной ксилемы. Между ними закладывается камбий и за счет его антиклинальных делений формируются вторичная флоэма и вторичная ксилема. Деятельность пучков камбия стимулирует межпучковую паренхиму к делению. Из нее формируется межпучковый камбий, который далее сливается с пучковым в одно камбиальное кольцо. За счет деятельности камбиального кольца происходит утолщение стебля. Для стеблей двудольных растений характерна дифференциация первичной коры, в состав которой входят колленхима( уголковая или пластинчатая), хлорофиллоносная паренхима и эндодерма – внутренний слой. Непучковое строение. В конусе нарастания прокамбиальные тяжи сливаются и образуют сплошной цилиндр, видимый на поперечном срезе в виде кольца. Кольцо прокамбия к наружи формирует кольцо первичной флоэмы, а внутрь – кольцо первичной ксилемы; между ними закладывается кольцо камбия. Клетки камбия делятся антиклинально (параллельно поверхности органа) и наружу откладывают кольцо вторичной флоэмы. Особенности строения стебля двудольных: 1) рост в толщину, 2) хорошо дифференцированная первичная кора (колленхима, хлорофиллоносная паренхима, крахмалоносная эндодерма), 3) коллатеральные(биколлатеральные) открытые(с камбием) пучки, 4) непучковое строение( пучки по кольцу или сливаются), 5) есть сердцевина, 6) наличие в ксилеме годичных колец(для древесных).
Вопрос 3. Подкласс дилленииды. Порядок первоцветные. Общая характеристика, эволюционные связи, лекарственные виды. Подкласс дилленииды( Dilleniidae). Порядок первоцветные или примуловые (Primulales). Распространены в умеренных и субтропических районах, часто в горах. Жизненная форма – многолетние и однолетние травы. Листорасположение очередное, супротивное или мутовчатое. Листья простые, без прилистников. Цветки обоеполые, правильные. Формула цветка: *Са(5)Со(5)А5G(5)_- первоцвет весенний(Primula veris). Околоцветник двойной, чаще пятичленный. Чашечка сростнолистная. Венчик спайнолепестный с длинной трубкой , лепестки изредка свободные. Андроцей состоит из 5 тычинок, свободных или прирастающих к трубке венчика. Завязь верхняя, одногнездная. Один столбик с головчатым рыльцем. Соцветия зончик, кистевидные или метельчатое. Плод – коробочка. Представители: вербейник обыкновенный(Lysimachia vulgaris) или вербейник монетчатый (L. nummularia), цикломен персидский(Cyclamen persicum). Для первоцветных характерны секреторные клетки, содержащие фенолы и сапонины.
4)А́стровые (лат. Asteráceae), или Сложноцве́тные (лат. Compósitae) Плод сложноцветных — семянка, то есть одногнёздный односемянной, нерастрескивающийся орешек с кожистой или деревянистой оболочкой.
цветкиТрубчатые звезда Са2-3-5~Со(5)А(5)G(2)нижняя
ложноязычковые стрелкаСа0;2;3;5;~Со(7)-~A0G0редко (2)нижняя
Билет 15 Вопрос 1. Задачи современной систематики растений, основные таксономические категории и таксоны, бинарная номенклатура. Понятие о виде. Систематика изучает многообразие растительных организмов и ее основная задача – классификация этого огромного многообразия. Вид представляет собой основную таксономическую категорию в систематике. Каждый вид принадлежит к какому-либо роду, род – к семейству, семейство – к порядку…При необходимости используют промежуточные таксономические категории – подвид, подрод, подсемейство…Таксонами принято называть реально существующие или существовавшие группы организмов, которые в процессе классификации отнесены к определенным таксономическим категориям. Например, ранги рода или вида являются таксономическими категориями, а род шиповник и вид шиповник собачий – конкретные таксоны. Научные названия всех таксонов выше вида состоят из одного латинского слова. Для видов приняты биноминальные газвания, состоящие из двух латинских слов. Первое слово обозначает род, второе – видовой эпитет. После названия заглавная буква обозначает фамилию ученого, описавшего этот вид.( Convallaria majalis L.). Подвиды – более мелкие таксоны внутри вида, занимающие свой ареал. Разновидности – не имеют своего ареала, но при этом морфологические признаки закреплены наследственно. Сорт – группа особей внутри вида, подвида, разновидностей, имеющих ряд наследственных, стойких признаков, не передающихся по наследству.
Вопрос 2. Особенности анатомии и морфологии листьев хвойных растений. Хвоя — листоподобные органы многих голосеменных (хвойных) растений — сосны, ели, туи и др. Ювенильная хвоя чаще всего бывает игловидной, заостренной, иногда серповидно изогнутой. Зрелая хвоя многих растений становится чешуевидной, укороченной. Хвоя содержит ценные биологические компоненты: хлорофилл (до 1,4 %), витамины, макро- и микроэлементы, фитогормоны, фитонциды, бактериостатические и антигельминтные вещества. У хвойных листья игольчатые (хвоя) или чешуйчатые, цельные с одной жилкой. Они имеют в большинстве случаев ясно выраженное ксероморфное строение. Они покрыты обычно толстым слоем кутикулы, энидермальные клетки у них мелкие и с сильно утолщенными стенками. Устьица погружены в углубления, которые заполнены зернышками воска (приспособление для уменьшения испарения). Под эпидермой большинства хвойных развита гиподерма (от греч. hypo—вниз и derma — кожа), или подкожица, из 1—3 слоев удлиненных толстостенных клеток, благодаря которой листья имеют обычно твердый наружный скелет, придающий им характерную жесткость. У некоторых листопадных родов (особенно у таксодиума и метасеквойи) строение листьев более или менее мезоморфное. У многих хвойных в мезофилле расположены крупные смоляпые каналы.
Вопос 3. Цикл развития покрытосеменных растений. После оплодотворения образуются семена, защищенные тканями околоплодника, и плод. Поэтому они и получили название покрытосеменных; пыльца попадает не на микропиле (как у голосеменных), а на рыльце пестика, по пестику прорастает до семяпочки и яйцеклетки; часто формируется хорошо развитый околоцветник, защищающий тычинки и пестики от повреждения и выполняющий вспомогательную функцию при опылении, привлекая насекомых-опылителей; они имеют более совершенную водопроводящую систему - кроме трахеид развиваются сосуды-трахеи, а флоэма (ситовидные трубки) уже содержит клетки-спутницы, которые отсутствуют у голосеменных растений; наблюдается еще большая редукция гаметофита по сравнению с голосеменными - гаметофит представляет собой несколько клеток, полностью паразитирующих на спорофите (последний доминирует); характерная особенность - так называемое двойное оплодотворение, которое заключается в том, что один из образующихся в мужском гаметофите спермиев сливается с яйцеклеткой, а другой - с двумя свободными или уже слившимися полярными ядрами гаметофита (тройное слияние). Из оплодотворенной яйцеклетки образуется диплоидный зародыш, а при тройном слиянии - триплоидное ядро вторичного эндосперма. Спорофит покрытосеменных растений - это обычные, хорошо знакомые деревья, травы, кустарники. Семязачатки развиваются внутри завязи пестиков и по сравнению с семязачатками голосеменных растений они небольшие, со слабо развитыми покровами и почти полностью лишены запаса питательных веществ. У цветковых растений в мегаспорангии образуется обычно одна материнская клетка мегаспоры, из которой в результате редукционного деления образуются четыре гаплоидные мегаспоры, но три из них отмирают и разрушаются, одна оставшаяся увеличивается в размерах. Ядро мегаспоры делится три раза подряд митозом и образует восьми ядерный женский гаметофит - зародышевый мешок. Затем свободноядерная стадия развития женского гаметофита переходит в клеточную. У входа микропиле образуются яйцеклетка и две клетки синергиды. У противоположного конца - три клетки антиподы. В центре зародышевого мешка располагаются полярные ядра (их два). В пыльниках тычинок образуются из спорогенных клеток материнские клетки микроспор; из каждой материнской клетки микроспоры в результате редукционного деления образуется по четыре гаплоидных микроспоры. Из микроспоры развивается сильно редуцированный мужской гаметофит, состоящий в большинстве случаев всего из двух клеток: маленькой генеративной и большой клетки - трубки (вегетативной клетки), называемой еще пыльцевым зерном (пылинкой). Пыльца из пыльников переносится на рыльце пестика и прорастает. По пыльцевой трубке передвигаются ядро вегетативной клетки и генеративное ядро, которое делится митотически и образует два спермия, один из которых проникает в яйцеклетку и сливается с ней, а другой - с полярными ядрами зародышевого мешка. Из оплодотворенной яйцеклетки - зиготы - развивается зародыш семени, а от слияния спермия с полярными ядрами образуется вторичный эндосперм семени. Синергиды и антиподы после оплодотворения исчезают. Эндосперм покрытосеменных растений триплоиден. После оплодотворения семязачаток превращается в семя, а его покровы - в кожуру семени; из завязи пестика (или других частей цветка) образуется плод, представляющий собой дополнительную защитную оболочку (покров). Прорастание семян стимулируется теплом и влагой, при этом необходим кислород. Зародыш и эндосперм набухают и разрывают семенные покровы. В результате клеточных делений, претерпеваемых зиготой после оплодотворения, образуется клеточная нить - подвесок. Из этих клеток у двудольных растений развиваются два первых листа (или семядоли) и стебелек. Из части стебелька, лежащей ниже семядолей, появляется первичный корень, обладающий положительным геотропизмом и направляющийся вертикально вниз в почву. Из верхней части стебелька (над семядолями) образуются стебель и листья. Прорастание семени первоначально идет за счет питательных веществ эндосперма до того момента, когда молодой проросток уже будет способен к самостоятельному синтезу органических веществ за счет фотосинтеза.
4.Это корень вторичного строения,так как в центре препарата 2 луча ксилемы, а между ними 2 открытых сосудисто-волокнистых пучка. Первичная кора отсутствовала.Покровная ткань пробка.
Билет 16. 1).Экологические факторы, их классиф. и роль в формир. экологич. групп растений
Свойства среды, оказывающие на живые организмы разностороннее воздействие, называются экологическими факторами.Они очень разнообразны: это световой, водный, температурный режим, состав и свойства почвы, жвижение, влажность воздуха, географическое положение, влияние человека и животных огранизмов.Эти факторы взаимно связаны, и изменение одного влечет за собой изменение другого или целого их комплекса.Группы эк.ф.:климатические-это температурные условия, почвенная и атмосферная влага, световые условия и ряд других,зависящих от географич. координат местности; почвенные (эдафические)- это свойства почвы, особенности ее физического и химического состава; топографические, связанные с особенностями рельефа.По отношению к действию какого-либо фактора выделябт экологические группы растений: гидрофиты- растения водных и излишне увлажненных мест, ксерофиты- засухоустойчивые, мезофиты- живущие в достаточных условиях увлажнения, гигрофиты-в местах с высокой влажностью воздуха и почвы.
2).Проводящие ткани, СВП
Проводящие ткани обеспечивают восходящий и нисходящий ток растения.Восходящий ток- это ток минеральных солей, раств. в воде, идущих от корней по стеблю к листьям по сосудам и трахеидам ксилемы(древесины).Нисходящий- ток орг.в-в от листье к корням по ситовидным элементам флоэмы (луба).Проводящими элементами ксилемы явл. трахеиды-вытянутые клетки с заостренными концами, имеют одревесневшую кл. стенку (бывают 4 типов- кольчатые, спиральные, пористые и лестничные) и сосуды( трахеи)- однорядный тяж клеток, называемых члениками.они произошли от трахеид.Их утолщения бывают кольчатыми, спиральными, лестничными, сетчатыми и пористыми.Проводящими элементами флоэмы явл. ситовидные клетки.У покрытосеменных- ситовидные трубки- продольный тяж клеток, называемый члениками.В ситовидном элементе разрушен тонопласт и ядро.Каждому чденику ситовидной трубки сопутствует клетка-спутница, участвующая в транспорте орг.в-в, создавая нисх.ток. Флоэма и ксилема образуют СВП.Они располагаются в ЦОЦ.Закрытые пучки- без камбия, открытые- с камбием между ксилемой и флоэмой.В результате деятельности камбия пучок разрастается и происходит утолщение органа.4 типа СВП: коллатеральные- флоэма лежит по одну сторону от ксилемы, бывают открытые (есть камбий-в органах 2-дольн.) и закрытые(стебли и корневища 1-дольн.); биколлатеральные- флоэма примыкает к ксилеме с двух сторон( в стеблях, корнях и корневищах 2-дольных); радиальные-лучи ксилемы расположена по радиусам, между ними флоэма (корни первичного строения); концентрические- складываются из кс., окружающей фл.- центрофлоэмные, центроксилемные- фл. окружает кс.
3).Лютиковые.
Подкласс Ранункулиды- Ranunculales включает порядки: лютиковые, маковые, пионовые.Порядок лютикоцветные- Ranunculales, среди них наибольшее хозяйственное значение имеют сем. барбарисовые( Berberidaceae) и лютиковые (Ranunculaceae). Барбарисовые: многолетние травы с простыми или сложными листьями, часть их видоизменена в острые иголки.Цветки мелкие, актиноморфные, с двойным околоцветником, собраны в кисти.Чашечка и венчик трех-, реже двухчленные.Тычинки в 2 круга, G монокарпный, с семязачатками. Завязь верхняя, рыльце расширенное.Плод монокарпий в виде продолговатой красной ягоды. Барбарис обыкновенный (Berberis vulgaris) *Сa3,3 Co3+3 A3+3 G1. Препараты из корней и корневищ подофилла щитовидного (Podophyllum peltatum) обладает противоопухолевой активностью.Лютиковые: многолетние травы, реже полукустарники.Листорасположение очередное, листья простые цельные или рассеченные, без прилистников.Цветки обоеполые, (а)циклические, актиноморфные, реже зигоморфные.Околоцветник простой или двойной, характерны стаминодии-нектарники и органы, промежуточные между стаминодиями и лепестками.Чашечка пятичленная, венчик пяти- или многочленный.Тычинок много.Характерны нектарники, связанные с тычинками.G апокарпный, реже монокарпный.Завязь одногнездная, верхняя.Плоды- монокарпии и апокарпии из листовок или орешков.Семена с эндоспермом и мелким зародышем.Соцветия цимозные, ботрические.Лютиковые делятся на 2 подсемейства: зимовниковые (Helleboroideae)- купальница(Trollius) , аконит(Aconitum), живокость(Delphinium).Формирование двойного околоцветника происходило за счет образования лепестков из стамодиев, т.е. лепестки тычиночного происхождения; собственно лютиковые (Ranunculoideae)- лютик(Ranunculus), василистник(Thalictrum), ветреница(Anemone).Формирование двойного околоцветника происходило за счет образования чашелистиков из верхних вегетативных листьев.Т.о. и лепестки, и чашелистики имеют листовую природу.Ценными лекарств. видами явл. адонис весенний (Adonis vernalis)- при серд.-сос. недостаточности; клопогон даурский(Cimicifuga dahurica) и василистник вонючий(Thalictrum foetidum).
4 Задача :это корневище двудольного так как середина объекта занята паренхимой, первичная кора широкая, из запасающей паренхимы и эндодермы с пятнами Каспари. В ЦОЦ по кольцу расположены сосудисто-волокнистые пучки.
Открытый колатеральные пучки .Эндодерма подковообразная и пятнами каспария. Первичная ткань эпидерма .в цоц входят перицикл,открыт коллатеральные паренхима запасающая
Билет 17. 1). Вегетативное размн.
Вегетативное размножение -размножение с помощью вегетативных органов: частей побега, корня, корневища, листа.Благодаря вегетативному размножению цветковые растения (в частности, сорняки) быстро распространяются на соседние участки и заселяют их.
При вегетативном размножении дочерние организмы сохраняют большое сходство с материнским организмом. Поэтому человек использует вегетативное размножение для сохранения у потомства ценных признаков материнских растений, для размножения культурных растений, при озеленении сел и городов, в сельском хозяйстве. Так, флоксы, маргаритки, крыжовник, смородину, малину размножают путем деления куста, землянику - усами, картофель - клубнями.
В практике в целях размножения проводят прививку - приживление части одного растения к другому. Прививкой размножают растения, у которых затруднено образование придаточных корней, когда хотят сохранить ценные признаки сорта. Для прививки у одного растения берут черенки или почку и сращивают с другим растением тем или иным способом (в расщеп, под кору, глазками, сближением и др.)Способы иск. вегет. размножения: деление кустов, размножение отпрысками, отводами, черенкование.
Хозяйственное значение вегетативного размножения:
Возможность быстрого воспроизводства двулетних и многолетних растений; необходимость сохранения наследственных признаков сорта; возможность сочетания полезных признаков нескольких растений в одном; успешная конкуренция с другими растениями; растения, появившиеся путем вегетативного размножения могут раньше перейти к плодоношению; растения, возникшие вегетативным путем, обладают такими же свойствами, какие были у материнского растения.
2).Устьичные комплексы.
Устьица- это высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток бобовидной формы и устьичной щели (своеобразного межклетника между ними).Имеются в листьях, но встречаются и на стебле.Стенки замыкающих клеток утолщены неравномерно: стенки, направленные к щели (брюшные), значительно утолщены по сравнению со спинными стенками.Щель может расширяться и сужаться, регулируя транспирацию и газообмен.Под щелью располагается крупная полость (межклетник), наз. дыхательной, кот. окружена клетками мезофилла листа.Замыкающие клетки окружены околоустьичными клетками, кот. совместно с замыкающими клетками образуют устьичный комплекс . 6 типов устьичных комплексов: амоноцитный (беспорядочный)- замыкающие клетки не имеют ярко выраженных околоустьичных клеток; характерен для всех высших растений, исключая хвойные;
анизоцитный( неравноклеточный)- замыкающие клетки устьица окружены тремя околоустьичными клетками, одна из кот. крупнее/меньше остальных;
парацитный (параллельноклеточный)- одна или более околоустьичных клеток расположены параллельно замыкающим;
диацидный(перекрестноклеточный)- 2 околоуст. клетки расположены перпендикулярно замыкающим;
тетрацидный- 4 околоуст. клетки, у 1-дольных
анциклоцидный- околоуст.кл. по кругу, у папоротников, голосеменных и цветковых.
Собственно эпидермальные клетки листа 2-дольн. растений в очертании волнистые, 1-дольн. вытянутые, ромбической формы.Механизм работы устьиц обусловлен осмотическими свойствами клеток.На свету происходит активный процесс фотосинтеза.Клетки насыщаются сахарами, что влечет за собой поступление в клетки ионов К, из-за чего концентрация клеточного сока в замыкающих клетках возрастает.Возникает разность концентрации клеточного сока околоустьичных и хамыкающих клеток.Вода из околоуст. клеток поступает в замыкающие, что ведет к увеличению объема замык.кл. и резкому возрастанию тургора.Замык.кл. приобретают выраженную бобовидную форму и устьичная щель открывается.При падении интенсивности фотосинтеза происходят обратные процессы (снижается образование сахаров, приток ионов К прекращается...) Вода путем осмоса уходит из замык.кл., понижая тургор=> устьичная щель закрывается.
3). Гречишные.
Подкласс кариофиллиды- Caryophyllidae вклучает порядки гвоздичные и гречишноцветные.Здесь преобладают травы и полукустарники.К порядку гречишноцветные( гречишные) (Polygonales) относится единственное семейство -гречишные( Polygonaceae).Это однолетние и многолетние травы и даже небольшие деревья.Корневая система стержневая или мочковатая.Листья простые, цельные с раструбом; листорасположение очередное.Цветки правильные, обоеполые, реже-однополые в ботрических соцветиях( кисть, метелка), реже одиночные в пазухах листьев.Околоцветник простой из 3-6 листочков, тычинок от 6 до 9, они свободные, располагаются в 2 круга.Гинецей ценокарпный из 2-4 срастающихся плодолистиков.Плод-псевдомонокарпий:трехгранный небольшой орех.Некоторые гречишные явл. ценными пищевыми продуктами( гречиха, щавель, ревень). Лек. значение имеет трава некоторых видов горцев как кровоостанавливающее и противовоспалительное средство, а также ревень тангутский в качестве слабительного.Гречиха съедобная (Fagopyrum esculentum), горец птичий(Polygonum aviculare), щавель конский(Rumex confertus), ревень тангутский(Rheum tanguticum).
4)Стебель однодольного растения
Закрытый коллатеральный сосудисто-волокнистый пучок
Покровная ткань-эпидерма
Билет 18.1).Ядро.
Ядро- самая крупная органелла, заключенная в оболочку из двух мембран, пронизанная ядерными порами.В ядре находится хромати: гетерохроматин (компактный, менее активный) и эухроматин (диффузный, активный)- в такой форме раскрученные хромосомы находятся в интерфазе.Хромосомы содержа нуклеиновую кислоту ДНК, отвечающую за наследственность.ДНК эукариот образует комплекс с гистоновыми белками и содержит информационно-функциональные участки- гены. Деление ядра лежит в основе размножения клеток.Внутри ядра находится хорошо заметная округлая структура-ядрышко, в нем происходит синтез рРНК.В ядре может быть 1 или несколько ядрышек.
2).Побеги, структуры стеблей 2-дольных
Побег- это стебель с листьями и почками.По характеру расположения побега в пространстве различают: прямостоящий побег, приподнимающийся побег, стелющийся (растет параллельно поверхности земли), ползучий( усы)- если на стелющимся стебле имеются пазушные почки, которые укореняются, вьющийся- обвивает опору, в нем плохо развиты механические ткани, цепляющийся- растет вокруг опоры с помощью усиков.В травянистых стеблях 2-дольных растений различают следующие анатомические структуры: в состав первичной коры входит колленхима (уголковая или пластинчатая), хлорофиллоносная паренхима и эндодерма.В эндодерме может накапливаться крахмал-это крахмалоносное влагалище.На границе ПК и ЦОЦ находится перициклическая склеренхима.Сердцевина представлена паренхимой.В сердцевине часто откладываются запасные питательные в-ва, могут встречаться клетки идиобласты, заполненные таннидами, кристаллами, слизью.Периферическая часть сердцевины называется перимедуллярной зоной.
3). Сосновые
Отдел сосновые (голосеменные)(Pinophyta)- древняя группа растений, средством для рассления у которой явл. семя.Половой процесс не связан с капельно-жидкой средой.Сем. сосновые (Pinaceae)-вечнозелёные (исключение — листопадные лиственница) деревья, реже кустарники; смолистые и пахучие, однодомные.Листья (хвоя) простые, опадают по одному (пучками у сосен), по листорасположению очерёдные и спиральные. Хвоинки вырастают либо по одной (спирально) на длинных отростках, либо пучками на коротких; молодые листья (если присутствуют) растут на длинных отростках. Листья имеют смоляные каналы. У большинства вечнозелёных родов хвоя живёт 2—5 лет.Почки покрыты плотно прилегающими друг к другу тонкими смолистыми чешуями. Генеративные почки образуют мужские шишки (микростробилы) и женские шишечки (мегастробилы). Микростробилы обычно малозаметны, располагаются на концах побегов, красные или жёлтые, их микроспорофиллы мелкие, несут на нижней стороне по два микроспорангия, содержащих пыльцу. Мегастробилы собраны в сложные шишки, которые имеют ось с сидящими на ней многочисленными парами чешуй: кроющей и расположенной в её пазухе семенной.Размножение сосны обыкновенной (Pinus sylvestris).Взрослое растение сосны явл. спорофитом (2п).В мае у основания ее молодых побегов образуются грозди желтоватых мужских шишек.Каждая шишка имеет ось, к кот. прикрепляются микроспорофиллы, имеющие с нижней стороны по 2 пыльцевых мешка-микроспорангия.В начале в них имеются диплоидные материнские клетки, из кот. весной в рез-те мейоза образ. гаплоидные микроспоры.Микроспора одноядерная покрыта 2 оболочками: интиной( внутренней) и экзиной (наружней) и имеет 2 воздушных мешка.Они возникают в рез-те отслоения экзины от интины.Внутри микроспорангия микроспора делится, т.е. образует заросток, но при этом не покидает микроспорангия.Ядро микроспоры делится и образуется 2 клетки.Из одной клетки образ.2 проталлиальные клетки, которые вскоре исчезают, из другой образ. антеридиальная клетка, формирующая мужские половые гаметы (спермии), и более крупная вегетативная клетка, из кот. формируется пыльцевая трубка.Пыльца-мужской гаметофит.При вскрытии пыльцевого мешка высыпается наружу.Женские шишки образ. на верхушках молодых побегов.Они состоят из наружных кроющих чешуй, под кот. располагаются 2 семязачатка.Молодой семязачаток состоит из нуцеллуса и ингумента.Нуцеллус- мегаспорангий.Он срастается с ингументом.Ингумент имеет отверстие( микропиле).В начале развития семязачатка нуцеллус состоит из однородных диплоидных клеток.Затем в среднейчасти обособляется 1 материнская археспориальная клетка.Она мейотически делится и образует 4 гаплоидные мегаспоры, из которых 1 способна к развитию.Эта мегаспора многократно делится митозом и образуется новая ткань-эндосперм, кот. явл. вегетативным телом женского гаметофита.Из 2 наружных клеток эндосперма, ориентированных к микропиле, образуются 2 архегония-женские органы полового размножения.Ветром пыльца из мужских шишек переносится на семязачатки.Улавливается пыльца каплями густой жидкости, заполняющей пространство между ницеллусом и ингументом.Эта жидкость затягивает пыльцу в пыльцевую камеру.Мужской гаметофит развивается на мегаспорангии.Экзина лопается, и вегетативная клетка, окруженная интиной, развивается в пыльцевую трубку, кот. внедряется в ткань нуцеллуса и растет в направлении архегония.Антеридиальная клетка делится и образует 2 клетки: клетку-ножку антеридия и генеративную клетку.Генеративная клетка перемещается в пыльцевую трубку
, по которой и достигает архегония.Из зиготы развивается зародыш, его развитие идет за счет запасных веществ эндосперма.Сформированное семя сосны соединяет в себе тело женского гаметофита и спорофит.Одна часть семени-эндосперм- это тело женского гаметофита(п).Другая часть- зародыш-зачаток дочернего спорофита (2п).Семенная кожура (2п) формируется из ингумента, а тонкая пленка(2п) представляет собой высохший нуцеллус.
4)А́стровые (лат. Asteráceae), или Сложноцве́тные (лат. Compósitae) Плод сложноцветных — семянка, то есть одногнёздный односемянной, нерастрескивающийся орешек с кожистой или деревянистой оболочкой.
цветкиТрубчатые звезда Са2-3-5~Со(5)А(5)G(2)нижняя
ложноязычковые стрелкаСа0;2;3;5;~Со(7)-~A0G0редко (2)нижняя
Билет №19 1.Экологические группы растений и их характеристика.
1)Группы растений по отношению к режиму влаги:
Гидрофиты (водные растения) – постоянно живущие в воде. Характеристика: обладают очень большой поверхностью тела; многие имеют недоразвитую или редуцированную корневую систему; гетерофилия (разнолистность); многие имеют систему ползучих побегов корневищ. (выполняют функцию запаса в-в и вегетат.размножения); высокая способность к вегетативному размножению; цветки выносятся над поверхностью воды, после опыления втягиваются в воду и семена созревают под водой; система межклетников заполненных воздухом ; эпидерма лишена защитных образований (волосков, воскового налета, кутикулы); проводящие и механические ткани развиты слабо; низкое осмотическое давление клеточного сока; лист красноватого цвета (накопление антоциана) – помогает лучше улавливать лучи света
Ксерофиты – растения сухих мест обитания. Делятся на 2 группы: суккуленты и склерофиты. Суккуленты –сочные мясистые многолетние растения, обычно с сильно видоизмененными побегами (кактусы, агавы, алоэ). Стебли у суккулентов голые, покрыты толстым слоем кутикулы. СВП развиты слабо. Вся толща ствола заполнена водой. Корни очень тонкие, отмирают в сухое время года. Осмотическое давление низкое. Склерофиты суховатые, твердые, жесткие растения с малой оводненностью тканей. Высокое осмотическое давление.
Мезофиты – растения мест достаточного увлажнения. Малоразвлетленная корневая система, в стеблях и корнях развита система межклетников. Мезофилл листа плохо дифференцирован на столбчатую и губчатую хлоренхиму, чаще представлен рыхлой губчатой тканью. Клетки эпидермы крупные, устьиц мало.
2) Группы растений по восприимчивости к свету:
Гелиофиты (светолюбивые) – растения открытых мест обитания, не выносящих длительного затемнения. Имеют толстые листья с мелкоклеточной столбчатой и губчатой паренхимой и большим количеством устьиц и хлоропластов. Высокая интенсивность фотосинтеза.
Сциофиты (теневыносливые) – растения хорошо развивающиеся на прямом солнечном свету, но выносящие некоторое затемнение. У листьев слабая дифференциаиция столбчатой и губчатой паренхимы, меньшее кол-во хлоропластов, меньшая интенивность фотосинтеза.
Тенелюбивые растения – растения, не выносящие сильного освещения.
3) Группы растений по потребностям в тепле:
Термофильные (теплолюбивые)
Криофильные (холодостойкие)
2. Части цветка и их функции. Форма цветоложа и положение завязи в цветке. Анатомическое строение пыльника и завязи.
Цветок – укороченный, метаморфизированный побег, выполняющий функции бесполого иполового размножения, образования семян и плодов. Основные части цветка: стеблевая (цветоножка и цветоложе, функция прикрепление), листовая (чашелистики и лепестки, функция защита), генеративная часть (пестики и тычинки; генеративная функция).
Цветоложе имеет различную форму: плоское, выпуклое, вогнутое. В результате срастания цветолож с нижней частей покрова и тычинок образуется гипантий (характерен для сем.Розовые). Завязь в цветке может быть нижней (пестик срастается с другими частями цветка) и верхней (образована только тканями пестика). Пыльник имеет две половинки, соединенные связником (продолжением тычиночной нити). Каждая половинка несет два гензда – микроспорангия. Гнезда пыльника называют пыльцевыми мешками. В зрелом пыльнике перегородки между гнездами часто не выявляются. Снаружи пыльник покрыт эпидермой, а под ней распологается слой клеток, за счет которых при подсыхании вскрываются гнезда.
Завязь.Место прикрепления семязачатков к завязи называется плацентой. Семязачатки сообщаются с плацентой с помощью семяножки. Плацента имеет вид вздутия , образованного тканями завязи.
3. Отдел моховидные. Класс листостебельные мхи. Подкласс сфагновые мхи, их характеристика, лекарственные виды.
Класс листостебельных мхов (Bryopsida) включает три подкласса: андреевые, сфагновые и бриевые. Наиболее многочисленные подклассы сфагновых и бриевых мхов.
Подкласс сфагновые мхи (Sphagnidae).
Подкласс представлен одним семейством сфагновые (Sphagnaceae) и единственным родом сфагнум (Sphagnum).
Сфагновые имеют прямостоячие стебли с пучковидно расположенными олиственными ветвями. Листья однослойные с двумя типами клеток - хлорофиллоносными и гиалиновыми (водоносными, крупные ромбовидные, бесцветные, мертвые). Хлорофиллоносные клетки содержат хлоропласты. Они распложены между крупными бесцветными водоносными клетками, лишенными клеточного содержимого. Благодаря наличию гиалиновых клеток сфагнумы способны всасывать большое количество воды.
Антеридии и архегонии образуются в верхней части стеблей. Спорофит представлен коробочкой, которая развивается из оплодотворенной яйцеклетки из архегония. Распространены сфагновые мхи в северном полушарии в умеренных и холодных широтах. Являются доминантами болот и влажных лесов.
Бриевые мхи (Bryidae) - зеленые (реже красно-бурые или черные) многолетние растения. Стебель сложного строения, нередко дифференцирован на ткани. Коробочка на ножке, после опадания крышечки открывается наверху широким отверстием устьем. По краю устья располагаются 1 или 2 ряда зубцов, образующих перистом. Перистом служит для рассеивания спор. Для зеленых мхов характерна хорошо развитая нитчатая протонема. Распространены бриевые мхи широко: на болотах, в хвойных лесах, на лугах, в горах и тундрах. Представители подкласса: мний (Mnium), кукушкин лен обыкновенный (Polytrichum commune).
Сфагновые мхи благодаря гигроскопичности и наличию антисептика сфагнола могут быть использованы в качестве перевязочного материала.
4) 16. Семейство розоцветные (Rosaceae)
Подсемейство розовые (Rosoideae)
Вид земляника лесная (Fragaria vesca) формула- *Ca5+5 Co5 A∞G∞ плод- земляничина(фрага)
Билет №20 1.Бесполое и половое размножение растений. Понятие о чередовании поколений. Место мейоза в жизненном цикле растений.
У высших растений различают два способа размножения: бесполое и половое. В бесполом размножении участвует только одна особь, способная к образованию спор или отделению жизнеспособных участков вегетативного тела, из которых в благоприятных условиях формируются полноценные дочерние особи. При половом размножении новая диплоидная особь образуется из диплоидной клетки - зиготы, возникающей при слиянии (копуляции) двух гаплоидных клеток, называемых гаметами. В цикле развития растений происходит закономерное чередование двух поколений: полового – гаметофит и бесполого – спорофит. Гаметофит способен к половому размножению и образует гаметы. Поскольку образование гамет происходит в результате митоза, они тоже гаплоидны. Сливаясь, гаметы образуют диплоидную зиготу, из которой вырастает следующее поколение — спорофит. Они называются спорофитами, потому что способны к бесполому размножению с образованием спор. Споры образуются в результате мейоза, т. е. здесь происходит возврат к гаплоидному состоянию. Гаплоидные споры дают начало гаметофитному поколению.
Одно из этих двух поколений всегда преобладает над другим, и на его долю приходится большая часть жизненного цикла; это поколение называют доминантным. У моховидных доминирует поколение гаметофитов, у всех остальных наземных растений — поколение спорофитов. Доминирующее поколение принято помещать в верхнюю половину схемы, изображающей жизненный цикл.