

Характеристика типа Ciliophora. Разнообразие, основные представители.
Этимология. Название инфузории происходит от латинского слова infusus - настой (в связи с обнаружением этих животных в настоях сена), название Ciliophora происходит от двух слов: латинского cilium - ресница и греческого слова phoros - несущий. Преамбула. Инфузории являются наиболее высокоорганизованными одноклеточными животными. Оценка их численности разными авторами колеблется от 3 до 30 тысяч видов. Размер инфузорий различных видов составляет от 10 мкм до 3-4.5 миллиметров. Форма тела чаще всего вытянуто-овальная или сферическая. Инфузории - преимущественно свободноплавающие одиночные организмы, прикрепленные и колониальные формы, а также паразитические виды и комменсалы встречаются значительно реже. В теле инфузорий, как и в теле других простейших, выделяются покровные структуры, цитоплазма и ядра. Покровные структуры. Инфузориям свойственен ресничный покров. У большинства инфузорий (класс Ciliata) реснички присутствуют в течение всей жизни, у представителей небольшого класса Suctoria реснички присутствуют лишь у расселительных стадий. Совокупность ресничек обозначается термином цилиатура. Выделяют соматическую и околоротовую цилиатуру. Расположение ресничек различно у разных групп: реснички могут покрывать тело сплошь или быть представленными на отдельных участках. Реснички инфузорий обычно расположены рядами, каждый такой ряд называется кинетой. Если ресничками покрыто все тело инфузории, то количество ресничек исчисляется тысячами, например у Paramecium caudatum ресничек около 15 тысяч. Репликация ресничек происходит перед делением клетки. Строение реснички аналогично строению жгутика и было рассмотрено ранее. Основной функцией ресничек инфузорий является, конечно, локомоция. При расположении ресничек рядами локомоция инфузории представлена плаванием, причем движение самой реснички включает гребной удар и более плавное распрямление и возвращение в исходное положение. Реснички в составе кинеты работают строго согласованно, причем движение соседних ресничек немного сдвинуто по фазе относительно друг друга. Реснички могут группироваться в пучки - цирры или в пластинки - мембранеллы. Цирры служат для "бега" - инфузория передвигается, опираясь циррами на субстрат, а мембранеллы - для создания тока воды, приносящего пищевые частицы. Реснички выполняют и сенсорные функции, у многих видов инфузорий эту функцию выполняют особые, укороченные реснички (щетинки), не участвующие в движении. Покровные структуры инфузорий включают пелликулу и кортекс (кортикальный цитоскелет). Некоторые авторы под кортексом понимают всю совокупность поверхностных структур клетки инфузорий: пелликулу, реснички, инфрацилиатуру и ряд других органелл. Соответственно наличию пелликулы и кортекса инфузории практически не способны менять форму тела, за исключением способности к сокращению всего тела или особого сократимого стебелька, что наблюдается у некоторых инфузорий. Пелликула инфузорий включает плазмалемму и подстилающие ее альвеолы (уплощенные мембранные мешочки). Напомним, что пелликула такой же структуры характерна для споровиков (и динофлагеллят). Между альвеолами располагаются кинетосомы ресничек. С кинетосомами связаны корешки, залегающие под пелликулой параллельно поверхности тела. Обычно от одной кинетосомы отходят три корешка: один построен из фибриллярных структур и называется кинетодесмальной фибриллой, два других состоят из микротрубочек. Кинетодесмальная фибрилла направлена вперед и тянется параллельно кинете. Один микротрубочковый корешок направлен в сторону соседней (правой) кинеты, он называется трансверсальным, другой микротрубочковый корешок направлен назад и называется постцилиарным. Кинетосома и связанные с ней корешковые структуры называются кинетиды. Кинетиды могут залегать поодиночке (монокинетиды) или по две (дикинетиды), или большим числом (поликинетиды). В двух последних случаях часто лишь одна кинетосома несет полный корешковый аппарат. В кортексе располагаются также микротрубочки и микрофиламенты, не связанные с ресничками. Микрофиламенты и микротрубочки инфузорий выполняют опорную и сократительную функции. В частности, корешковому аппарату кинетосом приписывают функцию якоря реснички. В покровных структурах инфузорий располагаются также экструсомы, клеточный анус (цитопрокт или порошица) и поры сократительных вакуолей. Экструсомы. Защитные образования - экструсомы - в общем были охарактеризованы ранее. У инфузорий встречается несколько типов экструсом, наиболее характерный тип представлен трихоцистами. Трихоцисты формируются из Гольджи-подобных структур и по сути дела являются небольшими вакуолями (в форме бутылки, кегли или т.п.), заполненными специфическим содержимым. Количество трихоцист у парамеций примерно втрое меньше количества ресничек (у Paramecium caudatum - около 5 тысяч). Трихоцисты выбрасываются при химических воздействиях или нападениях хищника. Выбрасывание трихоцист происходит в два этапа: сначала сливаются мембрана клетки и мембрана трихоциста, затем содержимое трихоциста в течение миллисекунды выбрасывается наружу, удлиняясь примерно в десять раз, на вершине развернувшейся нити находится заостренный наконечник. Затем мембрана трицисты отделяется от плазматической мембраны и распадается на мелкие пузырьки, которые позднее используются для создания новых трихоцист. Восстановление трихцист происходит примерно за 10 часов. Мутантные инфузории, лишенные трихоцист, вполне жизнеспособны, но поедаются хищниками быстрее, чем нормальные инфузории. Другим видом экструсом являются мукоцисты, они похожи на трихоцисты, но разворачиваются в течение нескольких секунд. Предположительно они участвуют в построении цист. У хищных инфузорий встречаются токсоцисты - экструсомы, служащие орудием нападения. Токсоцисты выстреливают трубчатую структуру, которая или выворачивается как палец перчатки, либо выдвигается телескопически. Вонзившись в тело жертвы, токсоцист парализует добычу и удерживает ее. Питание. Органоиды приема пищи и пищеварения. У инфузорий имеются особые органоиды приема пищи, представленные, например, у парамеций перистомом, ротовой воронкой, цитостомом, пищеварительной зоной цитоплазмы и цитопроктом. Перистом представляет собой длинную продольную выемку, косо расположенную на одной из сторон тела, которая условно обозначается как брюшная. На заднем его конце находится углубление - ротовая воронка. Ротовая воронка может нести только обычные реснички (в этом случае она носит название вестибулюм) или содержать сложные ресничные комплексы околоротовой цилиатуры (тогда ее называют буккальной полостью). Строение околоротовой цилиатуры крайне разнообразно и связано с характером питания. У парамеций околоротовая цилиатура представлена тремя мембранеллами (расположены слева), колебательные движения которых обеспечивают направленный ток воды, а также еще одной мембранеллой (расположена справа), которая задерживает пищевые частицы. Парамеции являются фильтраторами, причем фагоцитируют все частицы подходящего размера. На дне ротовой воронки находится цитостом. Напомним, что у всех простейших, имеющих пелликулу, цитостом представлет собой участок покровов клетки, где имеется только плазмалемма. Пищевые частицы поглощаются за счет фагоцитоза. Формирующаяся пищеварительная вакуоль сначала имеет трубчатую форму, но почти сразу его слепой конец начинает вздуваться, образуя вакуоль сферической формы. Начальный трубчатый участок пищеварительной вакуоли часто называют цитофаринксом (клеточной глоткой). У некоторых видов инфузорий цитофаринкс окружен кольцом микротрубочек, в этом случае он является постоянно существующим образованием. Пищеварительные вакуоли проделывают в центральной зоне цитоплазмы опредленный путь. В ходе этого перемещения в пищеварительные вакуоли порступают из лизосом ферменты и происходит пищеварение. В конце концов пищеварительльная вакуоль, содержащая уже только непереваренные остатки пищи, подходит к определенному участку покрова, где располагается цитопрокт. Место расположения цитопрокта у каждого вида строго фиксировано (у парамеций - позади ротового аппарата, примерно посередине между цитостомом и задним концом тела). В области цитопрокта прерваны пелликула и кортекс, и эндоплазма сорприкасается с плазмелеммой. Мембрана пищеварительных вакуолей, подошедших к цитопрокту, сливается с наружной мембраной, вследствие этого содержимое пищеварительной вакуоли попадает в окружающую среду. Как уже указывалось ранее, мембрана опорожненной пищеварительной вакуоли распадается на множество пузырьков, этот материал позднее используется клеткой для построения новых мембранных структур. Характер питания инфузорий различен: среди них имеются бактериофаги, альгофаги, фунгиофаги, хищники и даже каннибалы. Есть виды, потребляющие разнообразную пищу (полифаги), есть виды, специализированные на немногих или даже на одном виде пищи. Некоторые виды содержат в своем теле в качестве симбионтов зеленые водоросли (или только их хлоропласты) и получают питание за счет этих фотосинтетиков. Процесс поиска пищи инфузориями-фильтраторами описан Конрадом Лоренцом в упомянутой уже книге "Оборотная сторона зеркала": "Эти животные отыскивают с помощью своих топических рекаций (реакции, определяющие движение) среду с требуемыми свойствами, прежде всего с определенной концентрацией Н-ионов. Чаще всего встречается в природе угольная кислота (Н2СО3, быстро разлагается на углекислый газ и воду) , и ее повышенная концентрация обнаруживается в водах, где находят парамеций, чаще всего поблизости от гниющих остатоков растительных веществ, кислоту вызывают скопления бактерий, питающихся этими остатками. Связь эта столь надежна, а присутствие других, особенно ядовитых кислот, столь редко, что парамеция отлично обходится очень простой информацией, которую можно выразать словами: где имеется определенная концентрация кислоты, там собираются съедобные бактерии. Разумеется, программа вида не предусматривает экспериментирующего физиолога, который вводит в жизненное пространство парамеции каплю ядовитой щавелевой кислоты". Сосущие инфузории (Suctoria) являются подстерегающими хищниками. Они ведут прикреплееный образ жизни. От поверхности их тела отходят щупальца, которые представляют собой ветвящиеся или не ветвящиеся выросты цитоплазмы. Каждое щупальце на конце несет цитостом. Шупальце снабжено специализированными экструсомами - гаптоцистами. Если добыча прикасается к щупальцу, гаптоцисты выбрасываются, заякоривая добычу. Затем содержимое добычи поглощается через цитостом, то есть происходит фагоцитоз. Как уже указывалось, паразитизм среди инфузорий встречается не часто. Большая часть паразитических инфузорий паразитирует на коже и/или жабрах рыб. Единственной инфузорией, паразитирующей у человека, является балантидий - Balantidium coli. Вообще-то балантидий является типичным паразитом свиней, но при контакте человека со свиньями нередко осуществляется заражение человека, поэтому балантидий чаще встречается у сельского населения. Балантидий представлен в двух формах - вегетативной и цистной. Балантидий - самый крупный из кишечных протозойных паразитов: его длина на вегетативной стадии составляет 50 – 80 мкм. Как и у других протозойных паразитов кишечника, механизм распространения балантидия фекально-оральный. Паразитирование балантидия может иметь вид бессимптомного носительства или - при внедрении паразита в стенку кишечника - вызывать специфическую форму дизентерии, которая без лечения приводила к 50% смертности. Как комменсалов следует оценить инфузорий, обитающих в рубце и сетке жвачных (отр. Entodiniomorpha). Количество этих инфузорий составляет 2 млн. особей на 1 см3 содержимого желудка. Эти инфузории питаются бактериями и остатками пищи хозяина. Для хозяина эти инфузории, скорее всего, безразличны (хотя у северных оленей отмечалось проникновение такихз инфузорий вглубь тканей и поражение их): они не имеют ферментов, расщепляющих целлюлозу. Такие ферменты вырабатывают многочисленные бактерии, населяющие рубец хозяна и служащие пищей, как хозяину, так и инфузориям. Осморегуляция и выделение. Устройство сократительных вакуолей и принцип их деятельности были описаны ранее. Сократительные вакуоли имеют одну или - реже - 3-5 пор. Поры сократительных вакуолей расположены в поверхностных структурах клетки. Пора представляет собой короткий канал, стенки которого армированы микротрубочками. Основание поры закрывает мембранная диафрагма, которая исчезает при систоле и восстанавливается после выбрасывания жидкости. Ядерный аппарат. Для инфузорий характерен ядерный дуализм, то есть наличие ядер двух типов: микронуклеуса и макронуклеуса. Эти ядра различаются размерами и функциями. Микронуклеус обычно имеет сферическую форму, диаметр его - около 10 мкм. Если инфузория имеет 1 или два микронуклеуса, то они расположены рядом с макронуклеусом, если микронуклеусов несколько, то они расположены в разных частях эндоплазмы. Микронуклеус несет генеративную функцию и содержит диплоидный набор хромосом. Макронуклеус в вегетативной жизни транскрипционно активен. Количество ДНК в макронуклеусе в несколько сот раз превышает количество ДНК в микронуклеусе, однако макронуклеус содержит только около 2% ДНК микронуклеуса, остальная часть ДНК элиминируется. ДНК макронуклеуса организована не в форме целых хромосом, а в виде коротких фрагментов, но эти фрагменты представлены миллионами копий. У разных видов макронуклеус имеет различную форму (сферическую, бобовидную, колбасовидную, лентовидную, четкообразную и др.) и размеры (у крупных инфузорий длина макронуклеуса может достигать 2-3 мм). Макронуклеус может быть разделен на несколько частей. Бесполое размножение. Бесполое размножение проходит путем поперечного бинарного деления (монотомией). Для некоторых видов (преимущественно для Suctiria) характерно почкование. Половой процесс. Большую часть жизни инфузории не проявляют сексуальности, размножаясь бинарным делением. Для инфузорий (и только для них) характерен особый половой процесс - конъюгация, в ходе которой осуществляется обмен генетической информацией, но не происходит образования новых особей. Возможность вступления клеток в конюгацию определяется окончанием периода незрелости (у разных инфузорий эти сроки различны, у инфузории-туфельки этот период составляет около месяца). В ходе конъюгации две инфузории соединяются попарно цитоплазматическим мостиком. В каждой клетке происходит разборка старого макронуклеуса. Микронуклеус проходит двухступенчатый мейоз, в результате чего в каждой клетке формируется 4 гаплоидных ядра. Три из них резорбируются, а оставшееся делится митотически еще раз, так что в каждой клетке теперь находятся два ядра - пронуклеуса. Один из пронуклеусов (стационарное ядро) остается в клетке, другой (мигрирующее ядро) переходит по упомянутому ранее мостику в цитоплазму партнера. После этого цитоплазматический мостик разрушается, а пронуклеусы сливаются. В каждой из клеток формируется диплоидное ядро - синкарион (соответствует ядру зиготы). Два эксконъюганта возвращаются к самостоятельной жизни. В дальнейшем в серии специальных делений и сложного процесса созревания макронуклеуса в каждой клетке восстанавливается типичный набор ядер. Иногда при отсутствии партнеров для конъюгации половой процесс протекает в пределах одной клетки (автогамия). Интересно, что в пределах одного вида существует несколько различающихся генетически группировок инфузорий, которые называются типами спаривания. Конъюгируют между собой особи из разных, но комплементарных типов спаривания. При этом распознование происходит за счет выделения клетками специфических веществ. Жизненный цикл большинства инфузорий состоит из ряда бесполых поколений и периода конъюгации, которой жизненный цикл завершается форомированием генотипически новой особи. У сидячих видов, комменсалов и паразитических видов жизненный цикл усложняется наличием расселительных стадий.
Характеристика типа Porifera (Spongia). Отличия от настоящих многоклеточных. Разнообразие, основные представители.
Губки – древние примитивные многоклеточные животные. Обитают в морских, реже пресных водоемах. Ведут неподвижный прикрепленный образ жизни. Являются фильтраторами. Большая часть видов образует колонии. Не имеют тканей и органов. Почти все губки обладают внутренним скелетом. Скелет образуется в мезоглее, может быть минеральным (известковым или кремниевым), роговым (спонгиновым) или смешанным (кремниево-спонгиновым).
Выделяют три типа строения губок: аскон (асконоидный), сикон (сиконоидный), лейкон (лейконоидный) (рис. 1).
 рис. 1. Различные
типы строение губок:
1
- аскон, 2 - сикон, 3 - лейкон.
рис. 1. Различные
типы строение губок:
1
- аскон, 2 - сикон, 3 - лейкон.
Наиболее просто организованные губки асконоидного типа имеют форму мешка, который основанием прикреплен к субстрату, а устьем (оскулумом) обращен кверху.
Наружный слой стенки мешка образован покровными клетками (пинакоцитами), внутренний – воротничковыми жгутиковыми клетками (хоаноцитами). Хоаноциты выполняют функцию фильтрации воды и фагоцитоза.
Между наружным и внутренним слоями располагается бесструктурная масса – мезоглея, в которой находятся многочисленные клетки, в том числе образующие спикулы (иглы внутреннего скелета). Все тело губки пронизано тонкими каналами, ведущими в центральную атриальную полость. Непрерывная работа жгутиков хоаноцитов создает ток воды: поры → поровые каналы → атриальная полость → оскулум. Питается губка теми пищевыми частицами, которые приносит вода.
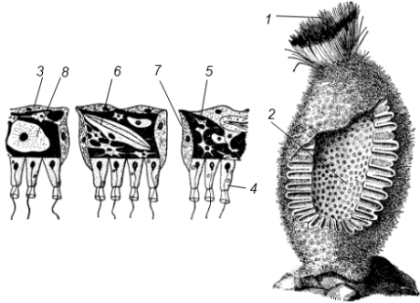 рис. 2. Строение
сикона (Sycon sp.):
1
- скелетные иглы, окружающие устье, 2 -
атриальная полость,
3 - пинакоцит, 4
- хоаноцит, 5 - звездчатая опорная
клетка,
6 - спикула, 7 - пора, 8 - амебоцит.
рис. 2. Строение
сикона (Sycon sp.):
1
- скелетные иглы, окружающие устье, 2 -
атриальная полость,
3 - пинакоцит, 4
- хоаноцит, 5 - звездчатая опорная
клетка,
6 - спикула, 7 - пора, 8 - амебоцит.
У губок сиконоидного типа происходит утолщение мезоглеи и образование внутренних впячиваний, имеющих вид карманов, выстланных жгутиковыми клетками (рис. 2). Ток воды в сиконоидной губке осуществляется по следующему пути: поры → поровые каналы → жгутиковые карманы → атриальная полость → оскулум.
Наиболее сложный тип губок – лейкон. Для губок этого типа характерен мощный слой мезоглеи с множеством скелетных элементов. Внутренние впячивания погружаются вглубь мезоглеи и приобретают вид жгутиковых камер, соединяющихся выносящими каналами сатриальной полостью. Атриальная полость у лейконоидных губок, так же как у сиконоидных выстлана пинакоцитами. Лейконоидные губки обычно образуют колонии с множеством устьев на поверхности: в виде корок, пластинок, комьев, кустов. Ток воды в лейконоидной губке осуществляется по следующему пути: поры → поровые каналы → жгутиковые камеры → выносящие каналы → атриальная полость → оскулум.
Губки обладают очень высокой способностью к регенерации.
Размножаются бесполым и половым способами. Бесполое размножение осуществляется в форме наружного почкования, внутреннего почкования, фрагментации, образования геммул и пр. При половом размножении из оплодотворенной яйцеклетки развивается бластула, состоящая из одного слоя клеток, имеющих жгутики (рис. 3). Затем часть клеток мигрирует внутрь и превращается в амебоидные клетки. После того, как личинка оседает на дно, происходит перемещение жгутиковых клеток внутрь, они становятся хоаноцитами, а амебоидные клетки выходят на поверхность и превращаются в пинакоциты.
Далее личинка превращается в молодую губку. То есть первичная эктодерма (мелкие жгутиковые клетки) становится на место энтодермы, а энтодерма – на место эктодермы: зародышевые пласты меняются местами. На этом основании зоологи называют губок животными, вывернутыми наизнанку (Enantiozoa).
Личинка большинства губок – паренхимула, по строению почти полностью соответствует гипотетической «фагоцителле» И.И. Мечникова. В связи с этим в настоящее время наиболее обоснованной считается гипотеза происхождения губок от фагоцителлообразного предка.
Тип Губки подразделяется на классы: 1) Известковые губки, 2) Стеклянные губки, 3) Обыкновенные губки.
Класс Известковые губки (Calcispongiae, или Calcarea)
Морские одиночные или колониальные губки с известковым скелетом. Скелетные иглы могут быть трех-, четырех- и одноосными. К этому классу относится сикон (рис. 2).
Класс Стеклянные губки (Hyalospongia, или Hexactinellida)
Морские глубоководные губки с кремниевым скелетом, состоящим из шестиосных игл. У ряда видов иглы спаиваются, образуя амфидиски или сложные решетки.
Скелеты некоторых видов очень красивы и используются в качестве коллекционных объектов и сувениров. Представители: корзинка Венеры (рис. 4), гиалонема.
Класс Обыкновенные губки (Demospongiae)
К этому классу относится подавляющее большинство современных видов губок. Скелет – кремниевый в сочетании со спонгиновыми нитями. У некоторых видов кремниевые иглы редуцируются, остаются лишь спонгиновые нити. Кремниевые иглы – четырех- или одноосные. Представители: туалетная губка (рис. 5), кубок Нептуна (рис. 6), бадяга, обитающая в пресных водоемах.
7. Radiata. Общая характеристика, разнообразие, представители. Чередование поколений.
8. Характеристика типа Ctenophora. Разнообразие, основные представители.
Этимология. Название происходит от двух греческих слов ktenos - гребень, phoros - несущий. Гребневики - это морские свободноплавающие в толще воды, значительно реже ползающие или сидячие радиальносимметричные животные. Часть видов выдерживает известное опреснение (имеются виды, живущие в Азовском море). Численность гребневиков оценивается в 80 видов. Размеры гребневиков - от нескольких миллиметров до 30 сантиметров. Лентовидный гребневик рода Cestum достигает длины 1.5 метра. Общий план организации. Тело плавающих гребневиков яйцевидной формы. На одном конце тела гребневиков - оральном полюсе - помещается рот, на противоположном - аборальном полюсе - специфический орган, именуемый аборальным органом. Ось, проходящая через оба полюса, называется главной осью. Большинство гребневиков имеет два длинных щупальца, снабженных многочисленными тонкими вы. Щупальца берут начала на дне особых углублений - щупальцевых влагалищ, расположенных на боковых сторонах тела. Различные органы гребневиков присутствуют в разных количествах - 2, 4 или 8. Таким образом, радиальная симметрия гребневиков оказывется комбинированной. Двулучевая симметрия гребневиков является полной, а четырехлучевая и восьмилучевая - неполными. Необходимо заметить, что четырехлучевая симметрия различных органов не вполне совпадает, что делает анализ строения тела гребневиков еще более сложным. Две плоскости двулучевой симметрии гребневиков перпендикулярны друг другу. В одной из этих плоскостей расположены щупальца гребневика (щупальцевая плоскость). Другая плоскость проходит через большой диаметр щелевидного (или эллиптического) рта, в этой же плоскости сплющена глотка (глоточная плоскость). Все плоскости симметрии гребневиков проходят через главную ось. На поверхности тела расположено 8 рядов гребных пластинок, они начинаются на аборальном полюсе, а заканчиваются на оральном, не доходя до рта. Скелета гребневики не имеют, опорную функцию несет мезоглея. Слои тела. Клеточные элементы. Стенка тела гребневиков образована эпидермисом, который снаружи покрыт слоем слизи, а изнутри подстилается мезоглеей. Эпидермис образован миоэпителиальными и эпителиальными клектами, часть которых дифференцирована в особые клеточные структуры: полицилиарные клетки, i-клетки, железистые клетки, продуцирующие слизь, клейкие клетки=коллоциты. В эпидермисе располождены также нервные клетки. Следует отметить, что ресничные полоициларные клетки приурочены к гребным пластинкам, а коллоциты - к щупальцам. В мезоглее находятся коллагеновые волокна и разнообразные клетки: миоциты, нервные клетки, амебоидные клетки. Мощным развитием мезоглеи гребневики напоминают медуз. Гастродерма включает эпителиальные ресничные клетки, железистые клетки, а также так называемые питающие клетки. Коллоциты. Коллоциты формируются из i-клеток эпидермы. Такая клетки имеет форму грибаи включает вздутую апикальную часть ("шляпку") и более узкий базальный отдел - стволик ("ножку"). Апикальный отдел содержит многочисленные пузырьки, заполненные клейким веществом. За счет этого вещества мелкие животные прилипают к коллооциты (или к нескольким коллоцитам). Стволик прочно, как якорь, закреплен в мышечном слое эпидермы. В состав коллоцитов входит также спиральная нить, отходящая от конца стволика и обвивающаяся вокруг него. Другим концом эта нить соединена со "шляпкой". Вероятно, эта спиральная нить, как пружинка, гасит механические нагрузки, когда приклеившаяся жертва пытается вырваться. Коллоциты используются лишь один раз, после чего замещаются новыми. Один из видов гребневиков Haeckelia rubra не имеет коллоцитов, это животное питается гидроидными медузами и встраивает стрекательные клетки жертв в свои щупальца, причем стрекательные клетки сохраняют дееспособность. Такие стрекательные клетки жертв, используемые их пожирателями, называются клептокниды. Органы движения. Движение. Как уже указывалось, на поверхности тела расположено 8 рядов гребных пластинок. Гребные пластинки состоят из сросшихся ресничек, в состав каждой гребной пластинки входит несколько тысяч ресничек. Длина ресничек очень велика, доходит до 2 миллиметров. Эффективный удар ресничек направлен в сторону аборального полюса, поэтому гребневики плывут ртом вперед. Однако при различных маневрах тела в пространстве направление удара может временно меняться на противоположное. Скорость передвижения гребневиков около 50 мм/сек. Такой способ передвижения используют пелагические гребневики типичной формы, которые являются самыми крупными одиночными животными, использующими ресничный способ локомоции. У всех гребневиков имеется мускулатура. Мышечные элементы расположены в эпидермисе (миоэпителиальные клетки) и в мезоглее (миоциты). Работа мышц у всех гребневиков обеспечивает открывание и закрывание рта, изменение объема глотки, движение щупалец, втягивание аборального органа и рядов гребных пластинок, (два эти действия носят защитный характер). Лентовидные пелагические гребневики и ползающие гребневики движутся за счет ундуляции тела, которая в свою очередь обеспечивается сокращением мышц. Аборальный орган. Аборальный орган служит для определения направления силы тяжести (орган равновесия). Этот орган размещается на аборальном полюсе. Он включает четыре изогнутые дужки (балансиры), образованные слившимися ресничками. На этих четырех дужках лежит статолит, состоящий из минеральных конкреций. От дужек по ыполнении к оральному полюсу отходят узкие ресничные бороздки, каждая из которых быстро выполненется на две. Получившиеся восемь рядов переходят в восемь рядов уже упоминавшихся гребных пластинок. Сверху аборальный орган прикрыт "шатром" из слипшихся ресничек. Дно аборального органа выстлано сенсорным эпителием. Под аборальным органом находится скопление нервных клеток. Дужки способны к спонтанному биению, то есть представляют собой пейсмейкеры. Генерируемые дужками импульсы передается гребным рядам по упомянутым ресничным бороздкам, но не за счет работы нервной системы. Частота колебаний дужек контролируется давлением лежащего на них статолита. При перемене положения тела меняется давление статолита на упомянутые дужки: давление на дужки, оаказавшиеся ниже других увеличивается, на остальные - уменьшается. Неравномерная нагрузка на дужки приводит к тому, что они начинают колебаться с разной частотой, что в свою очередь вызывает изменение частоты гребных движенией соответствующих гребных рядов. Таким образом осуществляется регуляция движения гребневиков. Возможно, что сенсорный эпителий дна аборального органа выполняет функции фоторецепции, барорецепции и хеморецепции. Нервная система. Нервная система гребневиков состоит из эпидермального сплетения нервных клеток, которые под рядами гребных пластинок образуют более плотные тяжи, идущие к аборальному полюсу. Напомним, что под аборальным органом располагается скопление нервных клеток. Сгущения нервных клеток также приурочены к щупальцам и глотке. Хемо- и терморецепторы разбросаны по всей поверхности тела и в большом количестве представлены вокруг ротового отверстия. Присутствуют нейроны и в мезоглее, эти нервные клетки иннервируют расположенные здесь же мышечные пучки. Пищеварительная система. Питание. Рот гребневиков ведет в ээпидермальную глотку (можно напомнить, что в соответствии с щелевидной формой рта глотка сплющена). В состав стенки глотки входят мышечные элементы, а также железистые клетки. Глотка переходит в желудок, котрый сплющен в плоскости, перпендикулярной плоскости сплющивания глотки. От желудка отходят каналы гастроваскулярной системы. Наличие этих каналов обеспечивает увеличение площади гастродермы, транспорт питательных веществ и обеспечение ими других органов. Каналы расположены тремя ярусами. В направлении к аборальному полюсу отходит акрогастрический канал, он на своей вершине делится на 4 веточки, две из них слепо замкнуты, две открываются наружу порами. Следует отметить, что эти четыре разветвления лежат не в отмеченных ранее плоскостях симметрии, а между ними. Следующим ярусом от желудка отходят два мезогастрических канала. Оба они дважды делятся дихотомически, так что формируется 8 разветвлений. Эти разветвления продолжаются в меридионально вытянутые восемь каналов, проходящие под рядами гребных пластинок. Функция этих каналов - снабжение гребных пластинок питательными веществами. У части видов гребневиков меридиональные каналы впадают в околоротовой кольцевой канал, у других видов такой канал отсутствует. От проксимальных участков мезогастрических каналов берут начало два канала, которые направлены к влагалищам щупалец и называются щупальцевыми. Третьим ярусом от желудка отходят два неразветвленных канала, направляющиеся к оральному полюсу, эти каналы называются метагастрическими. Все разветвления каналов, кроме двух веточек акрогастера, заканчиваются слепо. Стенки каналов гастроваскулярной системы представлены двумя типами клеток - высокими питающими клетками и плоскими ресничными. Питающие клетки меридиональных каналов перемежаются с клетками гонад. Меридиональные, щупальцевые и метагстрические каналы располагаются у поверхности тела, в непосредственной близости от органов, которые они снабжают питательными веществами: меридиональные - под рядами гребных пластинок и нервными тяжами, щупальцевые контактируют с влагалищами щупадец, метагастрические - со стенками глотки. Точно также апикальная часть акрогастрического канала прилежит ко дну аборального органа. Питающие клетки гастродермиса, расположены на той стороне канала, которой он прилежит к соответствующему органу. На противоположной стороне канала гастродермис представлен плоскими ресничными клетками. Здесь распорожены поры, окруженные двумя венчиками ресничек. Функция этих образований неизвестна, предполагается, что они участвуют в транспортировке питательных веществ к мышечным элементам мезоглеи. Гребневики - хищники, питаются они ракообразными и другими мелкими планктонными животными, медузами, другими гребневиками. Гребневики, питающиеся мелкими планктонными организмами, ловят добычу с помощью коллоцитов, расположенных на щупальцах. Когда добыча приклеилась к коллоцитам, происходят совместные движения щупальца (оно сокращается и подтягивается к ротовому отверстию) и всего тела (оно разворачивается), так что ротовое отверстие оказывается вплотную к добыче. Гребневики, питающиеся медузами и гребневиками, имеют просто устроенные щупальца или щупалец не имеют и ловят добычу прямо ртом, активно ее преследуя. Обработка добычи начинается уже в глотке: происходит механическая обработка – за счет сокращения мышц глотки и химическая - за счет действия ферментов. В глотке пища доходит до состояния мелкодисперсной смеси. Крупные фрагменты пищи, не поддающиеся обработке, выбрасываются через ротовое отверстие. Мелкодисперсная взвесь работой ресничек направляется в систему каналов. Здесь пищевые частички захватываются питающими клетками, которые переваривают их внутриклеточно и передают питательные вещества клеткам гребных пластинок, аборального органа, гонад и других структур. Перемещение пищевых частиц по каналам совершается в двух противоположных направлениях. Непереваренные фрагменты отчасти удаляются через поры двух разветвлений акрогастера, но большая часть их выбрасывается через рот. Газообмен и выделение. Газообмен и выделение осуществляются путем диффузии. Удаление продуктов обмена веществ может проходит через поверхность тела или через гастродерму. Половая система и размножение. Гребневики гермафродиты (раздельнополость харасктерна только для двух видов). Гонады расположены в гастродерме меридиональных каналов. С одной стороны каждого меридионального канала расположен яичник, с другой - семенник. Как уже указывалось, что созревающие яйцеклетки окружены группой питающих клеток, "клеток-нянек", которые снабжают их питательными веществами. Зрелые половые клетки через разрыв стенки поступают в канал, а оттуда через рот наружу, где и происходит оплодотворение. Для гребневиков характерно полное детерминированное дробленгие. Развитие у гребневиков прямое. Однако ювенильная стадия - цидиппида - некоторыми авторами оценивается как личиночная, Бесполое размножение в форме фрагментации известно только для ползающих гребневиков. Возможное происхождение гребневиков. В свете представлений гипотезы фагоцителлы гребневики оцениваются как потомки фагоцителл, достигших третьего эволюционного этапа, главным отличием которого является эпителизация периферического фагоцитобласта. Эпителизованный периферический фагоцитобласт соответствует каналам гастроваскулярной системы гребневиков. Эволюция пелагических гребневиков не была связана с оседанием на субстрат, в течение всего развития гребневики сохраняли пеланический образ жизни. Исключением явились немногочисленные ползающие гребневики, которые в какой-то момент исторического развития перешли к жизни на субстрате, причем осели они на оральный полюс (современные ползающие гребневики ползают ртом вниз). В свете представлений той же гипотезы гребневикам придается очень большое значение как группе, которая была организована по тому же плану, что и предки вторичноротых. Это предположение основано на том, что расположение энтодермальных карманов, которые в ходе онтогенеза вторичноротых дают начало мезодерме и полости тела, соответствует расположению главных каналов гастроваскулярной системы гребневиков. Экологические формы гребневиков. Как уже указывалось, среди гребневиков представлены пелагические (таких гребневиков большинство), ползающие формы и сидячие гребневики. В свою очередь пелагические планктонные гребневики представлены в с большинстве своем видами, имеющими яйцевидную или близкую к таковой форму, и лентовидные. Приведеное выше описание гребневиков базировалось на гребневиках с типовой формой тела. Отметим, что у гребневиков с уклоняющейся формой тела молодь (личинки) устроены по типовому плану строения. Для лентовидных гребневиков (2 вида в тропических и субтропических морях, в том числе "пояс Венеры" - Cestum veneris) характерна ремневидная форма тела. Главная ось этих животных короткая, зато в глоточной плоскости они сильно вытянуты. Движения генерируются при ундуляции тела. Развито только 4 ряда гребных пластинок аборальной половины тела. Щупальца рудиментарны. На оральной стороне тела расположены 4 бороздки (по 2 с каждой стороны), которые тянутся от краев тела к ротовому отверстию. Эти бороздки несут выросты, усаженные коллоцитами. Ползающие гребневики очень сильно сплющены по главной оси. Форма их тела в общем овальная, тело вытянуто в щупальцевой плоскости (до 25 миллиметров). Оральный полюс их обращен к субстрату. Рот находится в центре оральной стороны. Два щупальца расположены по краям тела. Ряды гребных пластинок отсутствуют (но имеются у личинок). Каналы гастрорваскулярной системы образуют сложную сеть за счет анастомозов меридиональных каналов. Глотка развита очень мощно, выворачивается наружу и формирует ползательную подошву. Часть видов способна не только ползать, но и плавать. В тропических и субтропических морях. Сидячие гребневики имеют форму толстого усеченного конуса, основание которого овально. Высота этого конуса соответствует главной оси. Тело несколько вытянуто в щупальцевой плоскости. Оральная поверхность обращена к субстрату и превращена в прикорептильный диск. Средняя часть рта зарастает, концы рта вытягиваются вверх в виде трубок. Из этих трубок выступают щупальца. Известны два вида из арктических морей.