

10096
.pdf51
завядания растения (или листьев). Тем не менее, она служит весьма выразительной экологической характеристикой водного режима растений (табл. 2). Эта величина достаточно постоянна и не зависит от кратковременных изменений водоснабжения.
Таблица 2
Примеры водного дефицита в листьях растений различных местообитаний (по данным разных авторов)
Растительное сообщество или местообитания |
Водный дефицит, % |
||
|
|
|
|
|
реальный |
сублетальный |
|
|
|
|
|
Водные и прибрежные растения |
0,1 – 3 |
5 – 15 |
|
Тундра: Зап. Гренландия |
2,8 – 12 |
– |
|
Лесотундра: древесные породы |
11 |
– 13 |
– |
кустарнички и травы |
3 – 10 |
– |
|
Еловый лес: кустарнички и травы |
0,1 – 12 |
14 – 55 |
|
Вечнозелёные леса: Средиземноморье |
12 |
– 55 |
– |
Юж. Чили |
– |
|
42 – 78 |
Луга: Воронежская обл. |
14 |
– 20 |
– |
Заросли крупнотравья: Камчатка |
от 5 – 8 |
– |
|
|
до 20 – 47 |
– |
|
Лесостепная дубрава: ранневесенние эфемероиды |
1,5 – 13 |
26 – 41 |
|
летневегетирующие травы |
3 – 40 |
40 – 68 |
|
Субальпийские луга: Западный Кавказ |
0,3 – 15 |
– |
|
Субнивальный пояс (2500 – 3000 м) |
|
|
– |
Центральный Кавказ |
25 – 32 |
– |
|
Сухие степи |
2 – 25 |
35 – 80 |
|
Пустыни Средней Азии |
от 3 до |
46 – 53 |
|
|
25 – 30 |
– |
|
Пустыни Северной Африки |
46 – 70 |
– |
|
|
|||
|
|
|
|
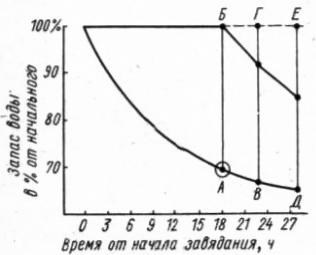
52
Рис. 22. Экспериментальное определение сублетального водного дефицита растений. Отделенные от растения листья или ветви постепенно теряют воду (нижняя кривая), что регистрируется путем периодического определения содержания воды. Через определенные промежутки времени пробы листьев из общего образца ставят во влажную камеру «решками» в воду. В результате на первых этапах завядания листья восстанавливают тургор, и содержание воды достигает начального (верхняя кривая). При более длительном завядании, когда влажность листьев падает ниже критической (точка Л), потеря воды при насыщении уже не восстанавливается, и завядание становится необратимым. Отрезок АБ – величина сублетального водного дефицита. Большие величины (ВГ, ДЕ н i. д.) уже детальны.
4.2. Экологические типы наземных растений по отношению к воде
Соответственно способу регулирования водного режима все наземные растения делятся на две основные группы.
П о й к и л о г и д р и д н ы е растения – это виды, не способные активно регулировать свой водный режим. У них нет каких-либо особенностей анатомического строения, способствующих защите от испарения (у большинства отсутствуют устьица). Они отдают или поглощают воду как физическое тело, и транспирация у них равна простому испарению. Содержание воды в клетках находится в равновесии с давлением паров в окружающем воздухе (т. е. определяется его влажностью) и зависит от его колебаний. К пойкилогидридным растениям относятся наземные водоросли, грибы, лишайники, некоторые мхи, а также некоторые высшие споровые растения – тонколистные папоротники тропических лесов. Немногочисленную группу составляют цветковые
53
растения, имеющие устьица, – отдельные представители семейства геснериевых, обитающих в расщелинах скал на Балканах и в Южной Африке, а также среднеазиатская пустынная осока Carex physodes. Считают, что у них пойкилогидридность – вновь приобретенный признак. Листья этих видов способны высыхать почти до воздушно-сухого состояния, но после смачивания вновь «оживают» и зеленеют.
Г о м е о г и д р и д н ы е растения составляют огромное большинство высших сосудистых растений. Они способны в определенных пределах регулировать потерю воды путем закрывания устьиц и складывания листьев. В их клеточных оболочках откладываются водонепроницаемые вещества (суберин, кутин), поверхность листьев покрыта кутикулой и т. д. Благодаря этому у гомеогидридных растений поддерживается на довольно постоянном уровне содержание воды в клетках и давление водяных паров в межклетниках. Транспирация по величине, дневной и сезонной динамике значительно отличается от свободного испарения смоченного физического тела. Именно эта группа растений составляет основную массу растительного покрова Земли (иначе вместо зеленеющих лесов и лугов даже в умеренных широтах мы наблюдали бы свежую зелень только после дождей).
По приуроченности к местообитаниям с разными условиями увлажнения и выработке соответствующих приспособлений среди наземных растений различают три основных экологических типа: гигрофиты, мезофиты и ксерофиты.
Гигрофиты – это растения избыточно увлажненных местообитаний с высокой влажностью воздуха и почвы. При довольно большом разнообразии местообитаний, особенностей водного режима и анатомоморфологических черт всех гигрофитов объединяет отсутствие приспособлений, ограничивающих расход воды, и неспособность выносить даже незначительную ее потерю. Ярко выраженные гигрофиты –
54
травянистые растения и эпифиты влажных тропических лесов, не выносящие сколько-нибудь заметного понижения влажности воздуха. Недаром в оранжереях многие виды приходится культивировать только под стеклом или в специальных влажных боксах. Даже в разгар сезона дождей мелкие эпифитные папоротники на стволах деревьев теряют тургор и засыхают, если на них в течение 2– 3 ч падают солнечные лучи. В эксперименте папоротники из семейства Hymenophyllaceae погибают уже при снижении относительной влажности воздуха до 90%.
Весьма гигрофильны плесневые грибы из родов Aspergillus и Penlcillium. Уже при влажности воздуха 95– 96% рост их резко замедляется, а при 85 % почти останавливается. Много гигрофильных видов среди шляпочных грибов, мхов, в том числе живущих в тропических лесах или близ источников (хороший пример представляют виды, поселившиеся на стенках фонтанов под струями каскадов в Новом Петергофе).
Черты гигрофитов (хотя и не столь выраженные) имеют травянистые растения темнохвойных лесов (Oxalis acetosella, Majanthemum bifolium, Circaea alpina и др.). К гигрофитам можно отнести и виды, растущие на открытых и хорошо освещенных местообитаниях, но в условиях избытка почвенной влаги: близ водоемов, в дельтах рек, в местах выхода грунтовых вод и т. д. В отличие от лесных гигрофитов А. П. Шенников называет их «световыми гигрофитами». В наших широтах примером могут служить прибрежные виды: калужница Caltha palustris, плакун-трава Lythrum salicaria, а в странах жаркого климата – папирус, болотные пальмы. Из культурных растений к таким «световым гигрофитам» относится рис культивируемый, как известно, на полях, залитых водой.
Характерные структурные черты гигрофитов – тонкие нежные листовые пластинки с небольшим числом устьиц, не имеющих толстой кутикулы, рыхлое сложение тканей листа с крупными межклетниками, слабое развитие воспроизводящей системы, тонкие, слаборазветвленные корни.

55
Способность гигрофитов к регуляции водного режима ограничена: устьица большей частью широко открыты, так что транспирация мало отличается от физического испарения. Благодаря беспрепятственному притоку воды и отсутствию защитных приспособлений интенсивность транспирации очень высока: у световых гигрофитов в дневное время листья могут терять за час количество воды, в 4– 5 раз превышающее массу листа. Высокая оводненносгь тканей гигрофитов поддерживается в основном за счет постоянного притока влаги из окружающей среды. Поэтому из группы гомеогидридных растений гигрофиты можно считать наиболее близкими к пойкилогидридным.
Рис. 23. Типы гуттации у разных растений (по Гродзинскому А. М., 1965):
1 – у злаков, 2 – у земляники, 3 – у тюльпана, 4 – у молочая, 5 – у беллевалии сарматской, 6 – у клевера

56
Другие характерные физиологические черты гигрофитов, обусловленные легкой доступностью влаги, – низкое осмотическое давление клеточного сока, незначительная водоудерживающая способность, приводящая к быстрой потере запасов воды. Хорошо известно, как быстро вянут в руках цветы, собранные по берегам водоемов. Особенно показательны для гигрофитов небольшие величины сублетального водного дефицита (см. табл. 2). Так, для кислицы и майника потеря 15– 20% запаса воды уже необратима и ведет к гибели.
А |
Б |
В
Рис. 24. Утренняя гуттация у растений дубового леса:
А – гусиный лук Gagea minima; Б – хохлатка Corydalis halieri (фото Самойлова Ю.
И.); В – сныть Aegopodium podugraria
В некоторых случаях у растений сильно увлажненных местообитаний возникает необходимость удаления избытка влаги. Обычно

57
это бывает, когда почва хорошо прогрета и корни активно всасывают воду, а транспирация отсутствует (например, утром или при тумане, когда
влажность |
воздуха |
100%). |
Избыточная |
влага |
удаляется путем |
г у т т а ц и и |
(рис. |
23, 24) – |
выделения |
воды |
через специальные |
выделительные клетки, расположенные по краю или на острие листа. К гуттации способны не только гигрофиты, но и многие мезофиты. Так, в украинских степях гуттация обнаружена более чем у половины всех видов. Многие луговые травы гуттируют столь сильно, что увлажняют поверхность почвы. По словам А. Фрей-Вислинга (1949), при более точном наблюдении можно установить, что большая часть росы, увлажняющей по утрам наши луга, не падает с неба, а выделяется самими луговыми травами.
Рис. 25. Рыхлые ткани с системой межклетников на поперечных срезах прибрежного растения Alisma plantago-aquatica (по Василевской В. К., из Шенникова А. П., 1950): А – лист; Б – стебель; В – корень
Избыточное увлажнение почвы может сказываться на растениях и отрицательно, поскольку затрудняет аэрацию, а, следовательно, дыхание и всасывающую деятельность корней. Борьба растений с избыточной влагой
– это, прежде всего, борьба за доступ воздуха. У многих травянистых видов избыточно увлажненных местообитаний (прибрежных, болотных) хорошо развита система воздушных полостей в листьях и стеблях, а также специальная воздухоносная ткань – аэренхима (рис. 25). Благодаря этому создается непрерывный путь от надземных частей к корням, по которому
58
подземные части растений снабжаются воздухом. Так, экспериментально показано передвижение кислорода вниз по стеблям прибрежных трав
(манника Glycerin aquatica, камыша Scirpus sylvaticus и др.), у которых стебель представляет собой полую трубку со стойкими стенками.
Влияние избытка влаги в почве испытывают и леса. На севере и северозападе европейской части России заболоченные леса и насаждения с постоянным или временным (весной и осенью) избытком влаги занимают около 60% всех лесных площадей. В таких местообитаниях на древесные породы угнетающе действует недостаток или полное отсутствие в воде свободного кислорода, а также появление недоокисленных соединений, накопление токсических веществ – СО2, H2S и др., что приводит к снижению поглощения корнями влаги и минеральных веществ из почвенного раствора, затруднению аэробного дыхания корней, сокращению фотосинтеза и другим нарушениям.
Ксерофиты – это растения сухих местообитаний, способные переносить значительный недостаток влаги – почвенную и атмосферную засуху. Ксерофиты распространены в областях с жарким и сухим климатом. Они обильны и разнообразны. К этой группе принадлежат виды пустынь, сухих степей, саванн, колючих редколесий, сухих субтропиков и т. д. В более гумидных районах ксерофиты участвуют в растительном покрове лишь в наиболее прогреваемых и наименее увлажненных местообитаниях (например, на склонах южной экспозиции, на скалах и т. д.).
Неблагоприятный водный режим растений в сухих местообитаниях обусловлен: во-первых, ограниченным поступлением воды при недостатке в почве и, во-вторых, увеличением расхода влаги на транспирацию при большой сухости воздуха и высоких температурах. Следовательно, для преодоления недостатка влаги возможны разные пути: увеличение ее поглощения и сокращение расхода, кроме того, способность переносить
59
большие потери воды. Все это используется ксерофитами при адаптации к сухости, но у разных растений в неодинаковой степени, в связи с чем некоторые авторы различают два основных способа преодоления ксерофитами засухи: возможность противостоять иссушению тканей или активно регулировать водный баланс и способность выносить сильное иссушение. Большое значение для ксерофитов имеют разнообразные структурные приспособления к условиям недостатка влаги. Корневые системы обычно сильно развиты (рис. 26), что помогает растениям увеличить поглощение почвенной влаги. По общей массе корневые системы ксерофитов нередко превышают надземные части, иногда весьма значительно. Так, у многих травянистых и кустарниковых видов среднеазиатских пустынь подземная масса больше надземной в 9– 10 раз, а у ксерофитов памирских высокогорных холодных пустынь – в 300– 400 раз. Корневые системы ксерофитов часто бывают экстенсивного типа (рис. 27), т. е. растения имеют длинные корни, распространяющиеся в большом объеме почвы, но сравнительно мало разветвленные. Проникновение таких корней на большую глубину позволяет ксерофитам (например, многим пустынным кустарникам) использовать влагу глубоких почвенных горизонтов, а в отдельных случаях – и грунтовых вод. Встречаются и широко раскинутые поверхностные корневые системы, приспособленные к поглощению скудных атмосферных осадков, смачивающих лишь верхние горизонты почвы.
У других видов (например, степных злаков) корневые системы интенсивного типа (рис. 27): они охватывают относительно небольшой объем почвы, но благодаря очень густому ветвлению максимально используют почвенную влагу. Корни ряда ксерофильных видов имеют специальные приспособления для запасания влаги («корневые шишки»). Они присутствуют у степной таволги Filipendula hexapetala, получившей из-за этого название «земляные орешки»; утолщенные и деформированные корни пустынных кустарников из рода Calligonutn. Надземные органы ксерофитов

60
также отличаются своеобразными (так называемыми ксероморфными) чертами, которые носят отпечаток трудных условий водоснабжения. У них сильно развита водопроводящая система, что хорошо заметно по густоте сети жилок в листьях (рис.28), подводящих воду к тканям. Эта черта облегчает ксерофитам пополнение запасов влаги, расходуемой на транспирацию.
Рис. 26. Корневая система |
Рис. 27. Различные типы корневых |
пустынного кустарника Pitiranthos |
систем. |
tortuosus (по Вальтеру Г., 1968) |
А – экстенсивный (верблюжья колючка |
|
Alhagi camelorum); Б – интенсивный |
|
(пшеница) |