

10096
.pdf41 |
|
холодных областей (или у переносящих холодные |
зимы) часто |
встречаются такие защитные морфологические особенности, как опушение почечных чешуй, зимнее засмоление почек (у хвойных), утолщенный пробковый слой, толстая кутикула, опушение листьев и т. д. Однако их защитное действие имело бы смысл лишь для сохранения собственного тепла гомеотермных организмов. Для растений же эти черты, хотя и способствуют терморегуляции (уменьшение лучеиспускания), в основном важны как защита от иссушения. В растительном мире есть интересные примеры адаптации, направленной на сохранение тепла (хотя и кратковременного) в отдельных частях растения. В высокогорьях Восточной Африки и Южной Америки у гигантских «розеточных» деревьев из родов Swecio, Lobelia, Espeletia и других от частых ночных морозов существует такая защита: ночью листья розетки закрываются, защищая наиболее уязвимые части – растущие верхушки (рис. 16). У некоторых видов листья опушены снаружи, у других в розетке скапливается выделяемая растением вода; ночью замерзает лишь поверхностный слой, а конусы нарастания оказываются защитным от мороза в своеобразной «ванне». Среди морфологических адаптаций растений к жизни в холодных местообитаниях важное значение имеют небольшие размеры и особые формы роста. Не только многие травянистые многолетники, но также кустарники и кустарнички в полярных и высокогорных областях имеют высоту не более нескольких сантиметров, сильно сближенные междоузлия, очень мелкие листья (явление нанизма, или карликовости, рис. 17). Кроме хорошо известного примера – карликовой березки Betula nana, можно назвать карликовые ивы Salix tolaris, Salix arctica, Salix herbacea и многие другие. Обычно высота этих растений соответствует глубине снежного покрова, под которым зимуют растения, так как все части, выступающие над снегом, гибнут от замерзания и высыхания. Очевидно, в образовании карликовых форм в

42
холодных местообитаниях немалую роль играют и бедность почвенного питания в результате подавления активности микробов, и торможение фотосинтеза низкими температурами. Но независимо от способа образования карликовые формы дают известное преимущество растениям в приспособлении их к низким температурам: они располагаются в припочвенной экологической микронише, наиболее прогреваемой летом, а зимой хорошо защищены снежным покровом и получают дополнительный (хотя и небольшой) приток тепла из глубины почвы.
А |
В |
Б |
Г |
Рис. 17. Нанизм высокогорных растений. А – В – растения альпийского пояса западного Кавказа, 3000–3500 м над уровнем моря (фото Примы В. М.). А – Jurinea filicifolia, Б –
Valdiva formosa, B – Arabis caucasica, Г – карликовая ива Salix herbacea (Восточные Карпаты, 1600 м над уровнем моря)

43
Рис. 18. Ветвление ели и переход к стланиковой форме в местообитаниях с сильными ветрами на верхней границе леса (Высокие Татры, Чехословакия) (по Plesnik P., 1971)
Другая адаптивная особенность формы роста – переход сравнительно крупных растений (кустарников и даже деревьев) от ортотропного (вертикального) к плагиотропному (горизонтальному) росту и образование стелющихся форм – стланцев, стлаников, стланничков. Такие формы способны образовывать кедровый стланик Pinus pumila, можжевельник
Juniperus sibirica, J. communis, J. turkestanica, рябину и др. Ветви стланцев распластаны по земле и приподнимаются не выше обычной глубины снежного покрова. Иногда это результат отмирания ствола и разрастания нижних ветвей (например, у ели, рис. 18). Иногда это рост дерева как бы «лежа на боку» с плагиотропным, укоренившимся во многих местах стволом и приподнимающимися ветвями. Интересная особенность некоторых древесных и кустарниковых стлаников – постоянное отмирание старой части ствола и нарастание «верхушки», в результате чего трудно определить возраст особи. Стланики распространены в высокогорных и полярных областях в условиях, которые уже не выдерживают древесные породы (например, на верхней границе леса). Своеобразные «стланиковые» формы в крайних условиях встречаются и у кустарничков

44
(рис. 19) и даже у видов лишайников, обычно имеющих прямостоячий кустистый рост: на скалах Антарктиды они образуют стелющиеся слоевища.
Рис. 19. Можжевельник туркестанский Juniperus turkesttmica на склонах хребта Терскей-Алатау (по Серебрякову И. Г., 1955):
А – древовидная форма (луто-во-лесной пояс, 2900 м над уровнем моря); Б – стланик (субальпийский пояс, 3200 м над уровнем моря)
К числу форм роста, способствующих выживанию растения в холодных местообитаниях, принадлежит еще одна чрезвычайно своеобразная – подушковидная. Форма растения-подушки образуется в результате усиленного ветвления и крайне замедленного роста скелетных осей и побегов. Мелкие ксерофильные листья и цветки расположены по периферии подушки. Между отдельными ветвями скапливаются мелкозем, пыль, мелкие камни. В результате некоторые виды растений-подушек приобретают большую компактность и необычайную плотность: по таким растениям можно ходить, как по твердой почве: таковы Silene acaulis. Gypsophila aretioides, Androsace helvetica, Acantholimon diapensioides.
Издали их трудно отличить от валунов. Менее плотны колючие подушки из родов Eurotia, Saxifraga.
45
Растения-подушки бывают разных размеров (до 1 м в поперечнике) и разнообразных очертаний: полушаровидные, плоские, вогнутые, иногда довольно причудливых форм (в Австралии и Новой Зеландии их называют «растительными овцами»).
Благодаря компактной структуре растения-подушки успешно противостоят холодным ветрам. Поверхность их нагревается почти так же, как и поверхность почвы, а колебания температуры внутри менее выражены, чем в окружающей среде. Среди других особенностей роста, помогающих растениям преодолевать действие холода, следует еще упомянуть различные приспособления, направленные на углубление зимующих частей растений в почву. Это развитие контрактильных (сократительных) корней – толстых и мясистых, с сильноразвитой механической тканью. Осенью они высыхают и сильно сокращаются в длину (что хорошо заметно по поперечной морщинистости). При этом возникают силы, втягивающие в почву зимующие почки возобновления, луковицы, корни, корневища (рис. 20).
Контрактильные корни встречаются у многих растений высокогорий, тундр и других холодных местообитаний. Они позволяют, в частности, успешно противостоять морозному выпиранию растений из почвы. В последнем случае они не только втягивают почку возобновления, но и ориентируют ее перпендикулярно поверхности, если растение повалено. Глубина втягивания контрактильными корнями варьируется от сантиметра до нескольких десятков сантиметров в зависимости от особенностей растения и механического состава почвы.

46
Рис 20. Контрактильные корни (по Strasburger E. et al., 1962):
А – Arum maculatum (углубление в почву от прорастания семени до взрослого растения); Б – участок корня Lilium martagon с поперечными морщинами; В – луковица Scilla sibirica
47
Глава 4. Вода в жизни растений
Вода – важнейший экологический фактор для всего живого на Земле. Для процессов обмена веществ со средой, составляющих основу жизни, необходимо участие воды в качестве растворителя метаболита. Так, у растений вода участвует в реакциях фотосинтеза: минеральные соли поступают в растение из почвы только в виде водных растворов и т. д. Вода – главная составная часть тела растений (от 30–40 до 95%). Даже находясь в анабиозе (например, глубокий покой семян), растения содержат воду. Особая роль воды для наземных растений заключается в постоянном пополнении больших трат ее на испарение в связи с развитием большой фотосинтезирующей поверхности. Вода, обусловливая необходимое тургорное давление, определенным образом участвует и в поддержании нормы наземных растений как организмов, не имеющих опорного скелета. Наконец, для большой группы растений, живущих в водоемах, морях и океанах, вода является непосредственной средой обитания.
4.1. Характеристика воды как экологического фактора
Основной источник воды для наземных растений – атмосферные осадки. Обеспеченность растительного покрова влагой в первую очередь зависит от общеклиматической характеристики местности. Есть районы, где в год выпадает несколько тысяч миллиметров осадков (области экваториального и муссонно-тропического климатов), а есть и такие, где среднегодовое количество осадков меньше 100 мм в год, причем дожди выпадают не каждый год (климат тропических пустынь).
Для оценки водообеспеченности растений мало знать только годовое количество осадков, поскольку одно и то же их количество может характеризовать и пустынный климат (в субтропиках), и очень влажный (в Арктике). Большую роль играет соотношение осадка и испаряемости (суммарного годового испарения со свободной водной поверхности), также весьма неодинаковой в разных районах земного шара. Области, в
48
которых эта величина превышает годовую сумму осадков, относят к а р и д н ы м (сухим, засушливым); здесь растения испытывают недостаток влаги в течение большей части вегетационного периода. Области, где растения достаточно обеспечены влагой, называют г у м и д н ы м и (влажными). Следует заметить, что граница между аридными и гумидными местообитаниями весьма условна. Иногда выделяют еще переходные – с е м и а р и д н ы е (полуаридные) местообитания. В случае крайне засушливых районов говорят об э к с т р а а р и д н ы х условиях.
Основной путь поступления воды в наземные растения – поглощение ее из почвы через корни (и ризоиды). Вода в почве находится в раздробленном состоянии: она вкраплена в поры разных размеров и форм и имеет огромную поверхность раздела с почвой, содержит ряд катионов и анионов. Поэтому почвенная влага весьма неоднородна по физическим и химическим свойствам. Различают три основные категории почвенной воды, отличающиеся по механизму удержания ее почвой: гравитационную,
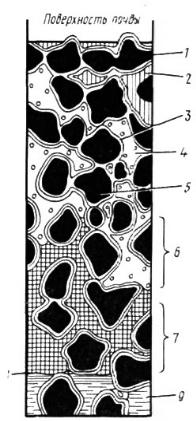
капиллярную и связанную (рис. 21). |
|
Гравитационная вода – |
недвижная вода, которая заполняет |
широкие промежутки между частицами почвы и просачивается вниз под действием силы тяжести (откуда и ее название), пока не достигнет грунтовых вод.
К ап и лл яр н ая вода заполняет тончайшие промежутки между частицами почвы и удерживается капиллярными силами сцепления. Обычно «капиллярная кайма» хорошо выражена над горизонтом грунтовых вод, выше могут находиться горизонты «подвешенной» капиллярной воды, не связанной с грунтовыми водами. Под влиянием испарения с поверхности почвы капиллярная вода может подтягиваться вверх, создавая восходящий ток влаги. Возможны и горизонтальные перемещения капиллярной влаги в почве под действием менисковых сил.
49
С вя зан н ая во д а удерживается на поверхности почвенных частиц адсорбционными силами. Количество ее значительно благодаря огромной поверхности почвенных частиц. Различают прочно связанную воду, находящуюся непосредственно на поверхности почвенных частиц, и рыхло связанную, находящуюся в более удаленных от частиц слоях. Первая неподвижна и не растворяет электролиты, вторая – обладает пониженной подлинностью и слабой растворяющей способностью.
Рис. 21. Вода в почве (по Качинскому Н. А., 1975):
1 – частицы почвы; 2 – гравитационная вода осадков, просачивающаяся в почву; 3 и 5 – гигроскопическая (пленочная и коллоидная) вода; 4 – почвенный воздух с парами воды, зона открытой капиллярной воды (часть пор заполнена воздухом, часть водой); 7 – зона замкнутой капиллярной воды (все поры заполнены водой); 8 – уровень грунтовой воды; 9 – грунтовая вода
50
Кроме перечисленных форм воды в почве всегда содержится парообразная влага, занимающая все свободные от воды поры. Это почти всегда (кроме пустынных почв) насыщенный водяной пар. При понижении температуры ниже 0° С почвенная влага переходит в лед ( вначале свободная вода, а при дальнейшем охлаждении – и часть связанной).
Только к этому показателю следует подходить с большой осторожностью, поскольку граница между двумя формами воды достаточно условна (зависит от метода определения) и между ними возможны взаимные переходы.
Водный дефицит. Случаи полного насыщения растений водой природных наземных местообитаний довольно редки. Обычно днем в результате расходов на транспирацию растения несколько недонасыщены водой. Величина этого недонасыщения называется водным дефицитом. Он определяется как разность между наибольшим содержанием воды в растении в состоянии насыщения (ранним утром или при искусственном донасыщении срезанных листьев во влажной камере) и реальным содержанием воды в растении в момент определения. Водный дефицит в природных условиях (называемый еще р е а л ь н ы м в о д н ы м д е ф и ц и т о м ) – величина изменчивая, зависящая от конкретных условий водоснабжения или погоды в тот или иной период. Она хорошо отражает динамику условий увлажнения и отчасти соотношение между расходом и поступлением воды в растения. Так, в жаркие и сухие периоды реальный водный дефицит растений сильно возрастает, а после продолжительных дождей или обильных рос падает до нуля.
Для характеристики выносливости растений к потере воды более употребителен другой показатель – с у б л е т а л ь н ы й в о д н ы й д е ф и ц и т (рис. 22), выражающий ту предельную величину потери воды, при которой растение еще сохраняет жизнеспособность и возможность восстановления нормальной оводненности тканей. Величина сублетального водного дефицита определяется в искусственных условиях в процессе