

Генетика микроорганизмов.
До 40-х гг. 20 в. считалось, что, поскольку у микроорганизмов нет ядерного аппарата и мейоза, на них не распространяются законы Менделя и хромосомная теория наследственности. С начала 40-х гг. микроорганизмы становятся объектом интенсивных генетических исследований. Именно на них были решены многие кардинальные вопросы современной генетики. Так, первое указание на то, что материальным носителем наследственности служит дезоксирибонуклеиновая кислота (ДНК), было получено в опытах на пневмококках (американские генетики О. Т. Эйвери, К. Мак-Леод и М. Маккарти). Примерно в то же время были начаты интенсивные генетические исследования на хлебной плесени - нейроспоре. Изучение многочисленных биохимических мутантов нейроспоры (Дж. У. Бидл и Э. Л. Тейтем, США) привело к установлению очень важного положения: один ген - один фермент (ныне это положение более точно формулируется так: один ген - одна полипептидная цепь). Генетические исследования микроорганизмов особенно интенсивно стали развиваться после того, как американские генетики С. Лурия и М. Дельбрюк показали на кишечной палочке (Escherichia coli), что и бактерии подчиняются мутационным закономерностям. Ранее существовавшее представление об адекватной, адаптивной изменчивости у бактерий возникло вследствие методической ошибки, заключавшейся в изучении культуры как единицы изменчивости. Был предложен новый принцип изучения изменчивости у бактерий - клональный анализ, т. е. изучение потомства одной клетки - родоначальницы клона. Важной вехой в развитии генетики микроорганизмов явился разработанный американскими генетиками Дж. и Э. Ледербергами метод реплик, или отпечатков, позволивший доказать, что мутации возникают у бактерий независимо от условий культивирования, и, кроме того, значительно упростивший приёмы отбора вариантов микроорганизмов с желаемыми свойствами. Оказалось, что в больших популяциях бактериальных клеток мутации возникают спонтанно. В 1946 был открыт половой процесс у бактерий (конъюгация), что позволило применить для их исследования генетический анализ. В рсзультате установлены наличие у бактерий рекомбинации, существование у них генетических групп сцепления и построены генетические карты их хромосом.
Важнейшими признаками живых организмов являются изменчивость и наследственность.
Основу наследственного аппарата бактерий, как и всех других организмов, составляет ДНК (у РНК-содержащих вирусов — РНК).
Наряду с этим наследственный аппарат бактерий и возможности его изучения имеют ряд особенностей:
бактерии — гаплоидные организмы, т. е. они имеют 1 хромосому. В связи с этим при наследовании признаков отсутствует явление доминантности;
бактерии обладают высокой скоростью размножения, в связи с чем за короткий промежуток времени (сутки) сменяется несколько десятков поколений бактерий. Это дает возможность изучать огромные по численности популяции и достаточно легко выявлять даже редкие по частоте мутации.
Высокая разрешающая способность методов генетического анализа бактерий и вирусов, позволяющая обнаружить их мутантов с частотой 10-9 и ниже.
Половая дифференциация, заключающаяся в существовании донорных и реципиентных бактериальных клеток, соответственно отдающих или воспринимающих генетическую информацию.
Наличие у бактерий обособленных фрагментов ДНК – плазмид, транспозонов и Is-последовательностей.
Генетическая система бактерий состоит из ядерных и внеядерных структур. Аналог ядра прокариотов значительно отличается от ядра эукариотических клеток. Он представлен нуклеоидом, лишенным оболочки и включающем в себя почти всю ДНК бактерии.
Хотя бактерии являются гаплоидными организмами, т.е. имеют один набор генов, содержание ДНК у них непостоянно, оно может при благоприятных условиях достигать значений, эквивалентных по массе 2, 4, 6 и даже 8 хромосомам.
Бактериальная хромосома состоит из одной двунитевой молекулы ДНК кольцевой формы. Молекула ДНК построена из двух полинуклеотидных цепочек. Каждый нуклеотид состоит из азотистого основания, сахара дезоксирибозы и фосфатной группы. Азотистые основания представлены пуринами (аденин, гуанин) и пиримидинами (тимин, цитозин). Каждый нуклеотид обладает полярностью. У него имеются дезоксирибозный 3' -конец и фосфатный 5' -конец. Нуклеотиды соединяются в полинуклеотидную цепочку фосфодиэфирными связями между 5' -концом одного нуклеотида и 3' -концом другого. Соединение между двумя цепочками обеспечивается водородными связями комплементарных азотистых оснований: аденина с тимином, гуанина с цитозином.
Нуклеотидные цепи антипараллельны: на каждом конце линейной молекулы ДНК расположены 5' -конец одной цепи и 3' -конец другой цепи. Наследственная информация у бактерий хранится в форме последовательности нуклеотидов ДНК, которая определяет последовательность аминокислотных остатков в молекуле белка. Каждому белку соответствует свой ген, т.е., дискретный участок на ДНК, отличающийся числом и специфичностью последовательности нуклеотидов. Бактериальная хромосома содержит до 4000 отдельных генов. Совокупность всех генов называется геномом. Внешнее проявление генома называется фенотипом . Размеры бактериальной хромосомы у различных представителей царства Procaryotae варьируют от 3 х 10 8 до 2,5 х 10 9 Д. Бактериальная клетка гаплоидна, а удвоение хромосомы всегда сопровождается ее делением.
У бактерий в естественных условиях передача генетической информации происходит не только по вертикали, т.е. от родительской клетки к дочерним, но и по горизонтали с помощью различных механизмов: конъюгации, сексдукции, трансдукции,трансформации.
У бактерий очень часто помимо хромосомного генома имеется дополнительный плазмидный геном, наделяющий их важными биологическими свойствами, специфическим (приобретенным) иммунитетом к различным антибиотикам и другим химиопрепаратам.
Большая роль в изменчивости бактерий и других организмов принадлежит так называемым транспонируемым генетическим элементам, т.е. генетическим структурам, способным в интактной форме перемещаться внутри данного генома или переходить от одного генома к другому, например от плазмидного генома к бактериальному и наоборот. Различают четыре класса транспонируемых элементов:
1) IS-последовательности;
2) транспозоны;
3) эписомы
4) плазмиды.
IS-(инсерционные) последовательности(англ. insertion-вставка) – это короткие фрагменты ДНК, мигрирующие от одной хромосомы к другой, или между хромосомой и плазмидой. IS-элементы имеют обычно размеры, не превышающие 2 тысяч пар оснований, или 2 кб( килобаза-тысяча пар оснований). IS-элементы несут только один ген, кодирующий белок транспозазу, с помощью которой IS-элементы встраиваются в различные участки хромосомы. Их обозначают цифрами: IS1, IS2, IS3 и т.д. Содержат только гены, необходимые для собственной миграции. Фенотипических признаков не кодируют, самостоятельно не реплицируются.
Функции IS-элементов:
Координировать взаимодействие транспозонов, плазмид и умеренных фагов как между собой, так и с хромосомой бактериальной клетки и обеспечивать их рекомбинацию.
Вызывать инактивацию гена, в котором произошла интеграция IS-последовательности («выключение гена»), либо, будучи встроенными в определенном положении в бактериальную хромосому, служить промотором (участками ДНК, регулирующих экспрессию подлежащих структурных генов бактерий – реципиентов), который включает транскрипцию соответствующих генов, выполняя регуляторную функцию.
Индуцировать мутации типа делеций или инверсий при перемещении и дупликации в 5-9 парах нуклеотидов при включении в бактериальную хромосому.
Транспозоны – это более крупные молекулы ДНК. Так же как IS-последовательности являются мигрирующими генетическими элементами. Представляют собой нуклеотидные последовательности, включающие от 2000 до 20500 пар нуклеотидов, которые несут генетическую информацию, необходимую для транспозиции. При включении в бактериальную ДНК они вызывают в ней дупликации, а при перемещении – делеции и инверсии. Они реплицируются только в составе бактериальной хромосомы. При этом новые копии транспозонов могут мигрировать в некоторые плазмиды и ДНК фагов, которые, проникая в бактериальные клетки, способствуют их распространению в популяции. Т.о. важнейшим свойством транспозонов является их способность к перемещению с одного репликона (хромосомная ДНК) на другой (плазмида) и наоборот. Помимо генов, ответственных за транспозицию, они содержат и структурный ген, кодирующий тот или иной признак.
Кроме того некоторые транспозоны, так же как плазмиды, выполняют регуляторную и кодирующую функцию. В частности, они могут нести информацию для синтеза бактериальных токсинов, а также ферментов разрушающих или модифицирующих антибиотики. Транспозоны имеют особые концевые структуры нескольких типов, которые являются маркерами, позволяющими отличать их от других фрагментов ДНК. При интеграции транспозонов в хромосому клетки животных или человека они приобретают удивительное сходство с провирусами, находящимися в составе их хромосом. Транспозоны, как и IS-последовательности обозначают порядковым номером:Tn1, Tn2, Tn3 и т.д. Транспозоны могут существовать и вне хромосомы (автономно), но неспособны к автономной репликации.
Эписомы (умеренные лизогенные и дефектные фаги). Встраиваясь в хромосому, эти фаги вызывают лизогению бактерий, которые могут приобретать новые признаки. Собственно эписомы – это вирусы, обладающие, подобно другим транспонируемым элементам, способностью в интактной форме переходить из одного генома в другой. Изменчивость лизогенных бактерий связана либо с приобретением генов, переносимых данными фагами от их предыдущих хозяев (бактерий-доноров), либо с экспрессией «молчащих» генов бактерий-реципиентов. В последнем случае фаговая ДНК, встраиваясь вблизи поврежденного промотора, заменяет его. При этом синтезируются определенные продукты, например протоксины дифтерийных бактерий, ряда клостридий и др.
Плазмиды – дополнительный внехромосомный генетический материал. Представляет собой кольцевую, двунитевую молекулу ДНК, гены которой кодируют дополнительные свойства, придавая селективные преимущества клеткам. Плазмиды способны к автономной репликации, т. е. независимо от хромосомы или под слабым ее контролем. За счет автономной репликации плазмиды могут давать явление амплификации: одна и та же плазмида может находиться в нескольких копиях, тем самым усиливая проявление данного признака.
Многие плазмиды имеют в своем составе гены трансмиссивности и способны передаваться от одной клетки к другой при конъюгации (обмене генетической информацией). Такие плазмиды называются трансмиссивными или конъюгативными, которые имеют более крупные размеры и наряду с генетической областью, контролирующей их репликацию, содержат также так называемую tra-область или tra-оперон(англ. transfer перенос). который определяет способность клетки, содержащей плазмиду, быть генетическим донором, т.е. вступать в конъюгацию с другой клеткой (реципиентом) и передавать ей свой генетический материал (плазмидную либо хромосомную ДНК). Под контролем tra-генов синтезируются поверхностные «половые» ворсинки (F-пили) клетки-донора, необходимые для ее конъюгации с клеткой-реципиентом, а также ферменты, обеспечивающие метаболизм ДНК в процессе конъюгации. Неконъюгативные плазмиды. обычно не содержат tra-оперона и поэтому не могут самостоятельно передаваться из одной клетки в другую. Однако передача неконъюгативной плазмиды возможна за счет продуктов (белков) tra-генов конъюгативной плазмиды, находящейся вместе с неконъюгативной плазмидой в одной и той же клетке. Конъюгативные плазмиды переносятся от бактерии к бактерии внутри вида или между представителями близкородственных видов в процессе конъюгации. Чаще всего конъюгативными плазмидами являются F - или R -плазмиды. Подобные плазмиды относительно крупные (25-150 млн Д) и часто выявляются у грамотрицательных палочек. Большие плазмиды обычно присутствуют в количестве 1-2 копий на клетку и их репликация тесно связана с репликацией бактериальной хромосомы.
Неконъюгативные плазмиды обычно имеют небольшие размеры и характерны для грамположительных кокков, но встречаются также у некоторых грамотрицательных микроорганизмов (например, у Haemophilus influenzae, Neisseria gonorrhoeae ). Мелкие плазмиды могут присутствовать в больших количествах (более 30 на клетку), так как только наличие такого количества обеспечивает их распределение в потомстве во время клеточного деления. При наличии в бактерии одновременно конъюгативных и неконъюгативных плазмид донор может передавать и неконъюгативные плазмиды за счет связывания генетического материала последних с факторами, обеспечивающими их перенос в процессе конъюгации.
Самостоятельная репликация плазмидной ДНК способствует ее сохранению и распространению в потомстве. Встраивание плазмид, так же как и профагов, происходит только в гомологичные участки бактериальной хромосомы, в то время как IS-последовательностей и транспозонов – в любой ее участок.
В зависимости от свойств признаков, которые кодируют плазмиды, различают:
1) R-плазмиды. Известно большое количество R-плазмид, определяющих устойчивость бактерий-хозяев к разнообразным лекарственным препаратам. Передача R-плазмид от одних бактерий к другим привела к их широкому распространению среди патогенных и условно-патогенных бактерий, что чрезвычайно осложнило химиотерапию вызываемых ими заболеваний.
R-плазмиды имеют сложное молекулярное строение. В их состав входят: r-ген, который может содержать более мелкие мигрирующие элементы – IS-последовательности, транспозоны и tra-опероны. R-ген, ответственнен за устойчивость бактерий к какому-либо антибиотику или модификацию. Значительное число r-генов является транспозонами, которые могут перемещаться от плазмиды- носителя в другие репликоны. В одном r-гене может содержаться несколько транспозонов, контролирующих устойчивость к разным антибиотикам. Этим объясняется множественная лекарственная резистентность.
Tra-оперон, обеспечивающий конъюгативность плазмиды, входит в состав –плазмид грамотрицательных бактерий. Грамположительные бактерии содержат в основном неконъюгативные плазмиды, которые могут передаваться от одной бактерии к другой путем трансдукции.
2) F-плазмиды. Представляет собой циркулярно замкнутую нить ДНК. Она контролирует синтез половых ворсинок (sex или F-pili), которые способствуют эффективному спариванию бактерий-доноров с реципиентными клетками при конъюгации. Данная плазмида реплицируется в независимом от хромосомы состоянии и передается при конъюгации в клетки бактерий-реципиентов. Перенос генетического материала (ДНК) детерминируется tra-опероном F-плазмиды (англ. transfer перенос), обеспечивающим ее конъюгативность. F-плазмиду можно удалить (элиминировать) из клетки, обработав последнюю некоторыми веществами, например акриловым оранжевым, в результате чего клетки теряют свойства донора. Сравнительно легкая элиминация и очень быстрая и эффективная передача F-плазмиды реципиентным клеткам дали основание считать, что она располагается в цитоплазме бактерий вне хромосомы. Однако F-плазмида может встраиваться в бактериальную хромосому и находиться с ней в интегрированном состоянии
3) Col-плазмиды. Кодируют синтез бактериоцинов. Это бактерицидные вещества, действующие на близкородственные бактерии;
Бактериоцины обнаружены у кишечных бактерий (колицины), бактерий чумы (пестицины), холерных вибрионов (вибриоцины), стафилококков (стафилоцины).
Колицины энтеробактерий (продуцируемые под контролем колициногенных плазмид) представляют собой вещества белковой природы. Известно более 25 типов колицинов, различающихся по своим физико-химическим свойствам и по способности адсорбироваться на определенных участках поверхности бактериальных клеток. Они обозначаются латинскими буквами A, B, C, D, E1, E2, K и т.д. При обычных условиях культивирования в большинстве клеток бактериальной популяции, содержащей колициногенные особи, синтеза колицина не происходит. Примерно в одной из 1000 клеток отмечается так называемая спонтанная продукция колицина. Однако количество колицинпродуцирующих клеток может быть резко увеличено при обработке бактерий УФ-лучами и некоторыми другими агентами. Механизм бактерицидного действия колицинов неодинаков. Показано, что после адсорбции на рецепторах наружной мембраны бактерий один из колицинов (Е3) нарушает функцию рибосом, другой (Е) является ферментом – эндодезоксирибонуклеазой. Имеются колицины, действующие на цитоплазматическую мембрану бактерий. Колициногенные (Col) плазмиды находятся в клетках энтеробактерий в автономном состоянии и передаются при конъюгацииибез сцепления с хромосомой. Однако некоторые из них (ColV, ColB) могут встраиваться в бактериальную хромосому и находиться в ней в интегрированном состоянии. Они так же как F-плазмиды, передаются путем конъюгации в реципиентные клетки, благодаря имеющемуся у них tra-оперону.
Широкое распространение бактериоциногении среди микрофлоры организма человека имеет экологическое значение как один из факторов, влиляющих на формирование микробных биоценозов. Вместе с тем колицины, продуцируемые кишечной палочкой – нормальным обитателем кишечника, могут губительно действовать на патогенные энтеробактерии, попавшие в кишечник, способствуя нормализации его естественного микробиоценоза.
Способность продуцировать различные типы колицинов используется для эпидемиологического маркирования. Такое типирование осуществляется путем определения типа Col-плазмиды (колициногенотипирование) или типа колицина, образуемого патогенными бактериями (колицинотипирование).
4) Плазмиды патогенности:
Tox-плазмиды. Кодируют выработку экзотоксинов;
Hly-плазмиды. Кодирует синтез гемолизинов
5) Плазмиды биодеградации. Кодируют ферменты деградации (утилизации) природных (мочевина, углеводы) и неприродных (толуол, камфора, нафталин) соединений, необходимых для использования в качестве источников углерода или энергии, что обеспечивает им селективные преимущества перед другими бактериями данного вида. Патогенным бактериям подобные плазмиды придают преимущества перед представителями аутомикрофлоры.
Например, урологические штаммы кишечных палочек содержат плазмиду гидролизации мочевины. Плазмиды биодеградации несут информацию об утилизации ряда сахаров ( лактоза, сахароза, раффиноза и др.) и образовании протеолитических ферментов.
Криптические (скрытые) плазмиды не содержат генов, которые можно было бы обнаружить по их фенотипическому проявлению.
Потеря клеткой плазмиды не приводит к ее гибели. В одной и той же клетке могут находиться разные плазмиды.
Изменчивость у бактерий
Различают два вида изменчивости – фенотипическую и генотипическую.
Ненаследственная, фенотипическая изменчивость, или модификация, микроорганизмов возникает как ответ клетки на неблагоприятные условия ее существования. Эта адаптивная реакция на внешние раздражители не сопровождается изменением генотипа и поэтому не передается по наследству
Модификации затрагивают большинство особей в популяции. С течением времени наблюдается реверсия, т. е. возвращение к исходному фенотипу после устранения действия фактора, вызвавшего их образование, поскольку исчезает потребность в сохранении данной модификации.
Могут измениться морфология (удлиняется), культуральные свойства (стафилококки без пигмента при недостатке кислорода) биохимические или ферментативные свойства, вырабатываются адаптивные ферменты Е. coli, фермент лактаза на среде — с лактозой.
К модификациям можно отнести включение «молчащих» генов (без их перестройки) некоторых микроорганизмов, в результате чего происходит смена их антигенов в ходе инфекционного заболевания.
К модификациям можно отнести также запрограммированные изменения генетической информации, в основе которых лежат миграции гена на хромосоме и встраивание его с разной частотой в определенные локусы, в результате чего происходит изменение признака. Существует и механизм возврата гена к исходной локализации, что приводит к восстановлению этого признака. К модификациям такого рода относятся изменения антигенной структуры гонококка, трепонемы сифилиса, боррелий возвратного тифа, холерного вибриона.
Модификации могут возникать под непосредственным действием антибиотиков, например пенициллина. Образующиеся при этом L-формы бактерий, лишенные клеточной стенки, могут сохраняться и даже размножаться внутри клеток хозяина и вновь реверсировать к исходной форме после прекращения действия пенициллина. При выращивании многих бактерий на питательной среде с суббактериостатическими концентрациями антисептиков также можно получить их модификации, характеризующиеся изменением морфологических и других признаков
Генотипическая изменчивость затрагивает генотип. В основе ее лежат мутации и рекомбинации.
Мутации (от лат. mutatio — изменять) — это передаваемые по наследству структурные изменения генов. При мутациях изменяются участки геномов (т.е. наследственного аппарата).
По происхождению бактериальные мутации могут быть:
спонтанными (самопроизвольными)
индуцированными (направленными)
Спонтанные мутации возникают самопроизвольно на протяжении всей жизни организма в нормальных для него условиях окружающей среды с частотой около 10 − 9 — 10 − 12 на нуклеотид за клеточную генерацию. Составляют естественный, или спонтанный, фон, величина которого колеблется в зависимости от типа мутации и вида микробной популяции. Они появляются в микробных популяциях in vitro и in vivo (в естественных биотопах организма человека) под влиянием самых разнообразных причин и событий, например ошибок в работе репарирующих ферментов, или ДНК-полимеразы во время репликации ДНК. Мутации происходят в результате ошибочного включения в синтезируемую дочернюю цепь вместо одного азотистого основания другого, некомплементарного имеющемуся в родительской цепи, например вместо аденина, комплементарного тимину, гуанина или цитозина.
Причиной изменения естественного фона могут быть инсертационные мутации, которые возникают при встраивании в хромосому микробной клетки Is-последовательностей, транспозонов и плазмид. При этом фенотип мутации зависит от места их интеграции: если она происходит вблизи промотора, то нарушается функция регуляторного гена, а вблизи структурного гена – синтез закодированного в нем продукта. При наличии у бактерий генов-мутаторов частота мутаций увеличивается в 100 и более раз.
Индуцированными мутациями называют наследуемые изменения генома, возникающие в результате тех или иных мутагенных воздействий в искусственных (экспериментальных) условиях в результате обработки микроорганизмов специальными мутагенами (хим. веществами, температурой, излучением и т.д.) или при неблагоприятных воздействиях окружающей среды.
По локализации различают мутации:
1) генные (точечные);
2) хромосомные;
При хромосомных мутациях происходят крупные перестройки структуры отдельных хромосом. В этом случае наблюдаются потеря (делеция) или удвоение части (дупликация) генетического материала одной или нескольких хромосом, изменение ориентации (поворота участка ДНК на 1800) сегментов хромосом в отдельных хромосомах (инверсия), а также перенос части генетического материала с одной хромосомы на другую (транслокация) (крайний случай — объединение целых хромосом, т. н. Робертсоновская транслокация, которая является переходным вариантом от хромосомной мутации к геномной).
Делеция.
Различают
терминальные (утрата концевого участка
хромосомы) и интеркалярные (утрата
участка на внутреннем участке хромосомы)
делеции.

Исследованные делеции редко захватывают протяженные участки хромосом, обычно такие аберрации летальны.
Дупликация.

Дупликации появляются в результате неравного кроссинговера (в этом случае второй гомолог несет делецию) или в результате ошибки в ходе репликации.
Хромосомная инверсия
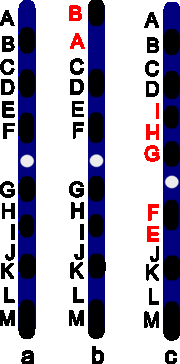
Инверсия. a — нормальная хромосома, b — парацентрическая инверсия, c — перицентрическая инверсия.
Различают парацентрические (инвертированный фрагмент лежит по одну сторону от центромеры) и перицентрические (инвертированный фрагмент лежит по разные стороны от центромеры) инверсии. При инверсиях не происходит потери генетического материала, поэтому инверсии как правило не влияют на фенотип.
Транслокации

Реципрокная транслокация 4 и 20 хромосом человека.
Помимо переносов участков с одной негомологичной хромосомы на другую, классифицируют также реципрокные транслокации (когда две негомологичные хромосомы обмениваются участками), Робертсоновские транслокации (при этом две негомологичные хромосомы объединяются в одну), а также транспозиции (перенос участка хромосомы на другое место на той же хромосоме).
Транслокация, реципрокная транслокация и транспозиция, которые не сопровождаются утратой генетического материала (сбалансированные транслокации), часто не проявляются фенотипически.
На генном уровне изменения первичной структуры ДНК генов под действием мутаций менее значительны, чем при хромосомных мутациях, однако генные мутации встречаются более часто. В результате генных мутаций происходят замены, делеции и вставки одного или нескольких нуклеотидов, транслокации, дупликации и инверсии различных частей гена. В том случае, когда под действием мутации изменяется лишь один нуклеотид, говорят о точковых мутациях. Поскольку в состав ДНК входят азотистые основания только двух типов — пурины и пиримидины, все точковые мутации с заменой оснований разделяют на два класса:
транзиции (замена пурина на пурин или пиримидина на пиримидин) и
трансверсии (замена пурина на пиримидин или наоборот).
Возможны четыре генетических последствия точковых мутаций:
сохранение смысла кодона из-за выроженности генетического кода (синонимическая замена нуклеотида),
изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация),
образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация). В генетическом коде имеются три бессмысленных кодона: амбер — UAG, охр — UAA и опал — UGA (в соответствии с этим получают название и мутации, приводящие к образованию бессмысленных триплетов — например амбер-мутация),
обратная замена (стоп-кодона на смысловой кодон).
По влиянию на экспрессию генов мутации разделяют на две категории:
мутации типа замен пар оснований
типа сдвига рамки считывания (frameshift). Последние представляют собой делеции или вставки нуклеотидов, число которых не кратно трем, что связано с триплетностью генетического кода.
Первичную мутацию иногда называют прямой мутацией, а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией, или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны.
При истинной реверсии восстанавливается не только фенотип, но и генотип. Восстановление одного фенотипа может произойти и в результате супрессии, т.е. подавления мутантного фенотипа, которое выражается в исправлении мутационного изменения. Так, например, если при первой мутации произошла вставка или выпадение пары нуклеотидов в одном из участков ДНК одного и того же гена, а в другом мутация противоположного рода (выпадение или вставка), то правильность считывания информации восстанавливается. Такая супрессия называется внутригенной.
При внегенной супрессии вторичные мутации, подавляющие выражение первичного мутационного изменения, локализованы в так называемых генах-супрессорах, кодирующих синтез транспортных РНК (тРНК). Мутации в таком виде могут привести к изменению тРНК, в результате чего в синтезируемый полипептид доставляется нужная аминокислота. При этом происходит восстановление фенотипа, но не генотипа.
По фенотипическим последствиям мутации подразделяют на:
нейтральные, фенотипически не проявляются какими-либо изменениями признаков, поскольку они заметно не отражаются на функциональной активности синтезируемого фермента.
Условно-летальные, которые приводят к изменению, но не к утрате функциональной активности фермента. В зависимости от условий окружающей среды микроорганизмы могут сохранять свою жизнеспособность или, наоборот утрачивать ее. Так, например, ts-мутанты (температурочувствительные) бактерии сохраняют способность к синтезу ферментов функционирующих при 370, но утрачивают этот признак при 420. В то же время у бактерий дикого типа соответствующие ферменты активны при обеих температурах.
Летальные мутации характеризуются полной утратой способности синтезировать жизненно важный для бактериальной клетки фермент или ферменты. Чаще всего эти мутации возникают при обширных делециях, захватывающих группу генов, или при других видах хромосомных мутаций. К ним относятся также мутации в генах, несущих информацию о синтезе ДНК-полимераз.
Мутации проявляются в фенотипе в виде утраты или изменения морфологических и биохимическихпризнаков, например жгутиков, пилей, капсулы, клеточной стенки; способности ферментировать какие-либо углеводы, синтезировать определенные аминокислоты, витамины и другие соединения, возникновении устойчивости к лекарственным или дезинфицирующим веществам и т.д.