

Лекция № 32. Механизм действия ферментов
1. Механизм действия ферментов
В химической реакции, катализируемой ферментом, фермент E соединяется с субстратом S, образуя нестойкий промежуточный фермент-субстратный комплекс E-S, который в результате химической реакции распадается с образованием фермента и продуктов реакции P:
![]()
В процессе реакции выделяют несколько стадий:
присоединение молекулы субстрата к ферменту;
преобразование промежуточного соединения;
отделение конечных продуктов реакции от фермента.
В реакциях анаболизма (A + B = AB) фермент может взаимодействовать с субстратами по отдельности или вместе:
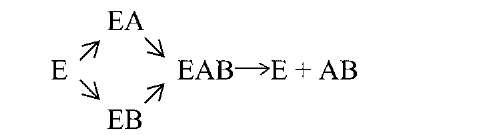
Реакции катаболизма также протекают с образованием промежуточного фермент-субстратного комплекса:
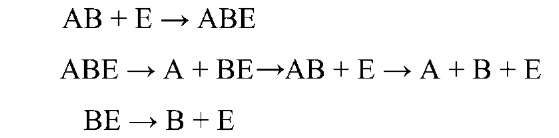
В образовании фермент-субстратных комплексов участвуют нековалентные (водородные, координационные связи, гидрофильно-гидрофобные взаимодействия) и ковалентные связи.
Большое значение в теории ферментативного катализа придается динамическим изменениям третичной структуры фермента при взаимодействии его с субстратом. Теория «индуцированного соответствия» Кошленда в противоположность жесткой модели «ключа и замка» Фишера допускает высокую конформационную лабильность молекулы фермента и гибкость его активного центра. При образовании фермент-субстратного комплекса субстрат индуцирует изменение конформации молекулы фермента так, чтобы активный центр принял определенную пространственную ориентацию, необходимую для взаимодействия с субстратом. Только в момент присоединения субстрата фермент будет находиться в активной T-форме, все остальное время он пребывает в неактивной R-форме. Существенное значение имеют точное соответствие между ферментом и субстратом, а также каталитические и термодинамические преимущества подобного соответствия. Предполагается наличие не только пространственной или геометрической компле- ментарности между ферментом и субстратом, но и электростатического соответствия, обусловленного спариванием полярных групп субстрата и активного центра фермента. Чем выше соответствие, тем более эффективен фермент в отношении данного субстрата.
С термодинамической точки зрения ферменты ускоряют химические реакции за счет снижения энергии активации. Энергия активации — энергия, необходимая для перевода всех молекул моля вещества в активированное состояние при данной температуре, или энергия, необходимая для запуска химической реакции. Фермент снижает энергию активации путем увеличения активированных молекул, которые на более низком энергетическом уровне уже становятся способными вступать в реакцию.
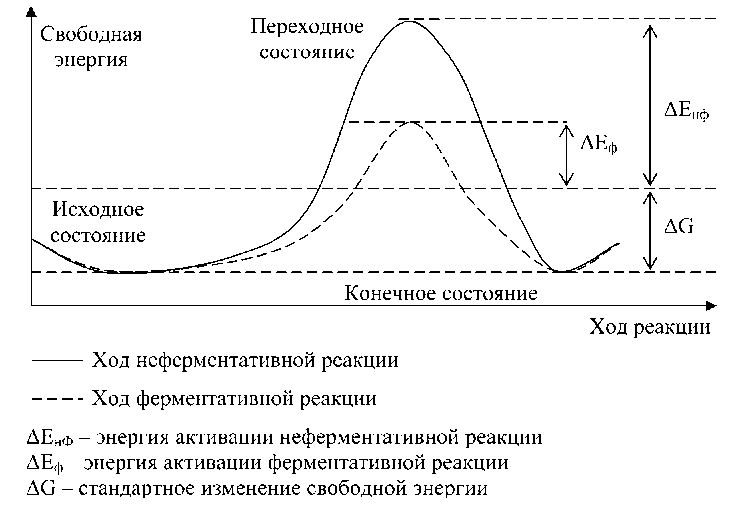
Величина стандартного изменения свободной энергии одинакова как для ферментативной, так и для неферментативной реакции, ферменты не изменяют равновесия между прямой и обратной реакциями и не влияют на величину свободной энергии реакции. Они лишь ускоряют наступление химического равновесия.
2. Кинетика ферментативных реакций
Живые организмы способны кинетически регулировать химические реакции, подавляя стремление к достижению термодинамического равновесия. Ферментативная кинетика изучает закономерности влияния химической природы реагентов (ферментов и субстратов) и условий реакции (таких как pH среды, температура, концентрация действующих веществ, присутствие активаторов или ингибиторов) на скорость ферментативной реакции.
Любая химическая реакция характеризуется константой термодинамического равновесия Кр. В реакции:
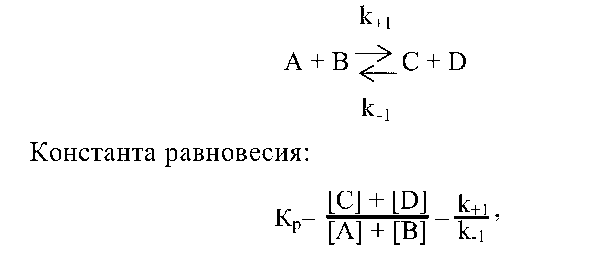
где [A], [B], [C], и [D] — концентрации действующих веществ; k+1 — константа скорости прямой реакции; k_i — константа скорости обратной реакции.
Величина, обратная константе равновесия, — константа диссоциации фермент-субстратного комплекса К8.
![]()
Константа диссоциации фермент-субстратного комплекса зависит от химической природы фермента и субстрата и определяет степень их сродства — чем ниже К8, тем выше сродство фермента к субстрату. При низкой концентрации субстрата скорость реакции прямо пропорциональна концентрации субстрата и подчиняется кинетике первого порядка. При высокой концентрации субстрата скорость реакции, достигнув своего максимума, становится постоянной, не зависящей от концентрации субстрата и подчиняется кинетике нулевого порядка. Скорость реакции в этом случае целиком определяется концентрацией фермента. Общая теория ферментативной кинетики, разработанная Л. Михаэ- лисом и М. Ментеном, предполагает, что если ферментативный процесс протекает в виде реакции:

где E — фермент; S — субстрат; P — продукт реакции, то количественное соотношение между концентрацией субстрата и скоростью ферментативной реакции выражается уравнением:
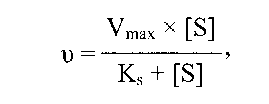
где ν — наблюдаемая скорость реакции при данной концентрации субстрата [S];
Ks — константа диссоциации фермент-субстратного комплекса;
Vmax — максимальная скорость реакции при полном насыщении фермента субстратом.
При высокой концентрации субстрата и низком значении Ks скорость реакции максимальна, при низкой концентрации субстрата скорость реакции прямо пропорциональна концентрации субстрата. Уравнение Бриггса—Холдейна учитывает влияние продуктов реакции на скорость ферментативного процесса:
![]()
где Km — константа Михаэлиса, определяемая опытным путем:
![]()
Константа Михаэлиса численно равна концентрации субстрата в моль/л, при которой скорость данной ферментативной реакции составляет половину от максимальной. Она всегда больше константы диссоциации фермент-субстратного комплекса на k+2 / k+1.