

Нейроанатомия эмоций
Для обнаружения и анализа мозговых структур, непосредственно ответственных за возникновение и реализацию эмоциональных реакций, огромное значение имела методика прямой стимуляции мозга электрическим током через заранее вживленные электроды [Hess, 1968]. Успехи стереотоксической нейрохирургии обусловили допустимость применения этой методики у человека в диагностических и лечебных целях, что открыло возможность сопоставления эффектов стимуляции с речевым отчетом пациентов о своем эмоциональном состоянии [С. Сэм-Джэкобсон, Р. Гис, X. Дельгадо, Н. П. Бехтерева, В. М. Смирнов и др.)« На рис. 23, заимствованном из работы Н. П. Бехтеревой [1971], показаны зоны, раздражение которых вызывало у пациентов эмоциональные реакции с различной выраженностью и постоянством.
Наиболее полную сводку данных, полученных с помощью лечебно-диагностических стимуляций, привел в своей монографии В. М. Смирнов [1976].
Суммируя результаты собственных наблюдений и сведения, почерпнутые в литературе, В. М. Смирнов отмечает, что при раздражении ядер миндалины пациент сообщает о возникновении состояний страха, гнева, ярости, изредка - удовольствия. Стимуляция перегородки, напротив, как правило, сопровождается переживанием эйфории, наслаждения, полового возбуждения, общего подъема настроения. При раздражении переднего и заднего отделов гипоталамуса наблюдаются реакции тревоги и ярости, а при стимуляции структур среднего мозга - широкий спектр эмоций от гнева и напряжения до полового возбуждения с выраженной положительной окраской. В отличие от перечисленных мозговых образований стимуляция гиппокампа не сопровождается ни страхом, ни яростью, ни удовольствием. Зарегистрированы только спутанность сознания, временная потеря контакта с врачом и эпизодически - страх в виде вторичной эмоциональной реакции субъекта на расстройство восприятия окружающего мира.
При раздражении мезэнцефального отдела ствола и неспецифического таламуса возникают состояния повышенной активации или инактивации. Активационные состояния имеют положительную эмоциональную окраску, реже активно-отрицательную (гнев, злоба, но не тоска и печаль). Инактивационные состояния характеризуются успокоением и безразличием. Возникающие при этих двух функциональных состояниях ощущения необычайной «легкости тела» или его «тяжести» не удалось непосред ственно связать с изменениями мышечного тонуса или вестибулярных функций.
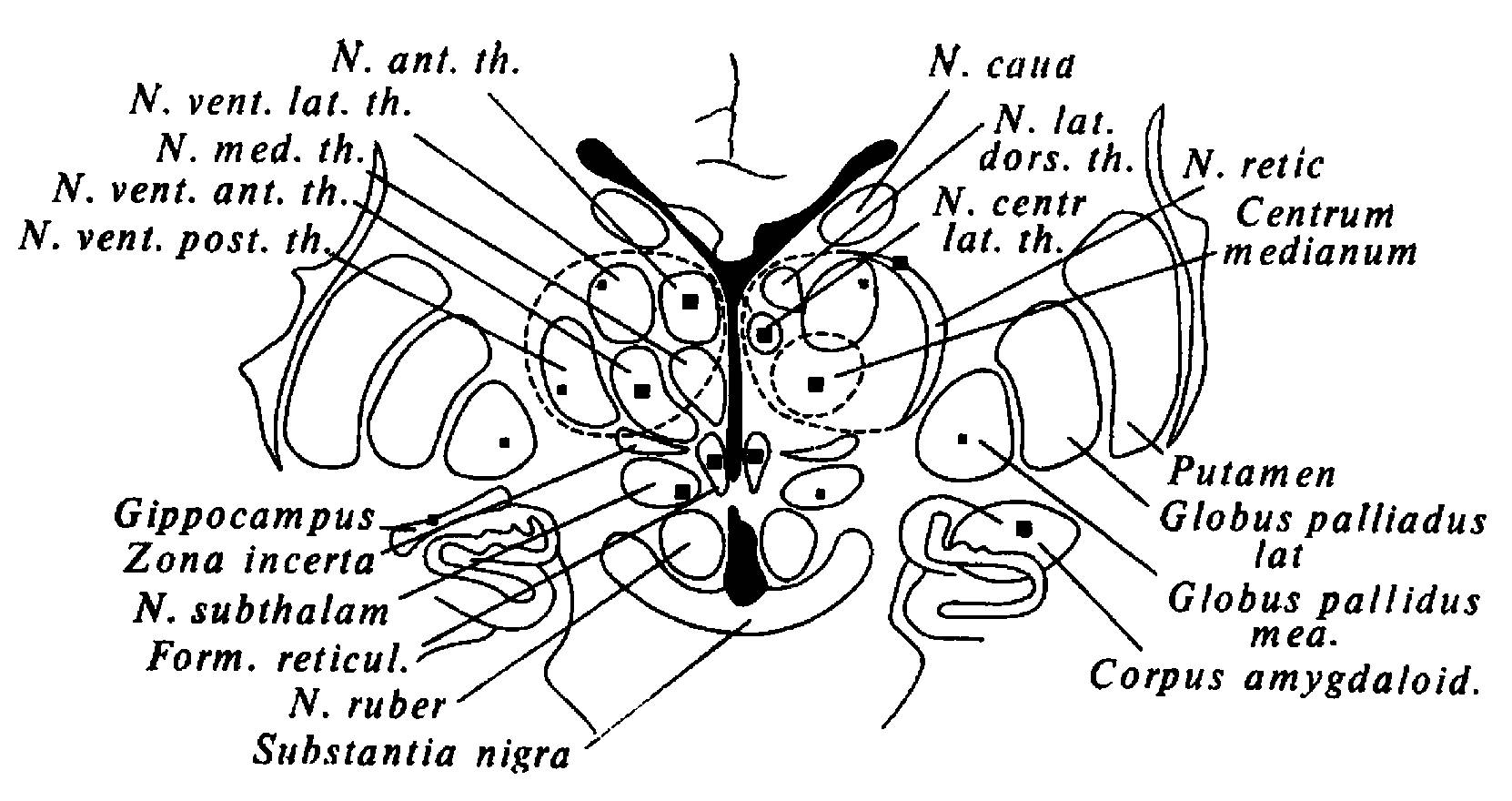
Рис. 23. Схема мозга человека
Крупными черными квадратами помечены зоны, раздражение которых вызывало эмоциональные реакции большей выраженности и с большим постоянством; маленькими квадратами - те зоны, при раздражении которых эмоциональные реакции развивались менее постоянно и были менее отчетливыми (по Н. П. Бехтеревой).
Помимо перечисленных выше эмоциональных состояний, пациенты В. М. Смирнова сообщали о кратковременных реакциях, возникавших в момент электрического раздражения. К ним относятся: чувство растерянности, недоумения (мезэнцефальные отделы ствола, субталамус, ретикулярное таламическое ядро); страх, причина которого субъекту неясна или связана с эмоциональной реакцией на соматические сдвиги (лимбикоретикулярная система); удовольствие в виде приятных, не всегда точно квалифицируемых ощущений; немотивированная радость, возникающая при стимуляции срединного центра таламуса, бледного шара, мезэнцефального отдела ствола.
На протяжении многих лет нейрофизиологи пытались найти тот отдел мозга, который в наибольшей мере ответственен за возникновение эмоциональных состояний: гипоталамус (Гесс, Гельгорн), таламус (Кенон, Бард), гиппокамп (Беритов, Пейпец, Брэйди), ретикулярная формация (Линдсли) и т. д. Эта тенденция получила отражение даже в названиях соответствующих теорий: «таламической», «гипоталамо-лимбической», «акти-вационной». Новейшей модификацией подобного подхода к проблеме локализации эмоций являются представления Мак Лина [McLean, 1970]. По мнению Мак Лина, в гипоталамусе репрезентированы стереотипные формы поведения, как правило, сопровождающиеся эмоциями (реакции защиты, нападения, поиска, добывания), а к вышерасположенным отделам мозга приурочены те или иные мотивации: к миндалине - потребности самосохранения (голод, защита и т. п.), к перегородке - продолжение рода, секс, к цингулярной и префронтальной коре - зоо-социальные, групповые, стадные мотивации; в неокортексе высших позвоночных аккумулирован их индивидуально приобретенный опыт.
Однако результаты экспериментов с прямым электрическим раздражением различных мозговых образований, равно как с их разрушением, не позволяют рассматривать ту или иную структуру в качестве локального «центра» эмоций страха, ярости, удовольствия и т. п. Даже полная хирургическая изоляция гипоталамуса у кошек [Ellison, Flynn, 1968] не ведет к выпадению защитных и пищевых реакций с характерной для них эмоциональной окраской. Оценивая сложившуюся ситуацию, А. В. Вальдман пишет: «По существу попытки топографического представительства в головном мозгу «эмоций», равно как и других психических функций, являются отголоском локализационализма... Вопрос о «субстрате эмоций» в настоящее время может быть решен определенно только в отношении топографии интегративных центров зон эмоционально-выразительных проявлений» [Вальдман, 1972, с. 14]. С этой точкой зрения солидаризируется и В. М. Смирнов: «...трудно назвать мозговые структуры, ответственные исключительно за эмоции... Поэтому мы вполне согласны с А. В. Вальдманом [1972], что нейрофизиологическое изучение эмоций и эмоционального поведения не должно быть направлено на поиск структурных элементов эмоций и что его следует базировать на системном принципе» [Смирнов, 1976, с. 166].
Близких взглядов придерживается в своих последних работах Р. Айсааксон: «Не существует одиночных анатомических образований для одиночных поведенческих функций... не существует единых поведенческих функций, которые могли бы быть приурочены к анатомическим образованиям таким образом, чтобы эти функции оказывались полезными в любых обстоятельствах и при всех внешних условиях» [Isaacson, 1978].
Понимая всю обоснованность разочарования в результатах поиска «точечно» локализованных эмоциогенных структур, мы вместе с тем не можем вернуться к расплывчатым представлениям об эмоциях как функции «мозга в целом», как следствии «корково-подкорковых отношений». При всей сложности проблемы экспериментальная нейрофизиология эмоций не вправе отказаться от фундаментального принципа «приурочивания динамики к структуре», завещанного нам И. П. Павловым.
Сегодня уже невозможно найти исследователя, который представлял бы себе нервные «центры» эмоций в виде ограниченного участка нервной ткани. Каждый, кто пытается рассмотреть организацию церебрального субстрата эмоций, непременно говорит о системе, о широко разветвленной констелляции нервных образований, представленных на различных уровнях головного мозга высших животных и человека.
Нейрофизиологии эмоций особенно близко то понимание доминантного очага как «функционального органа», как системы, которое мы связываем с именем А. А. Ухтомского. «Доминантный очаг,- пишет В. С. Русинов [1967, с. 200],- это констелляция А. А. Ухтомского, образующаяся как система в ходе текущей деятельности организма на всех этажах центральной нервной системы в разных ее местах, но с первичным очагом в одном из отделов и с переменным значением функций отдельных компонентов системы». А. А. Ухтомский специально подчеркнул, что фактором, определяющим временную целостность «функционального органа», является достижение определенного приспособительного результата. Он писал: «С именем «органа» мы привыкли связывать представление о морфологически сложившемся, статически постоянном образовании. Это совершенно не обязательно. Органом может быть всякое временное сочетание сил, способное осуществить определенное достижение [Ухтомский, 1950, с. 279. Курсив наш - П. С]. Идею результата как системообразующего фактора в дальнейшем энергично разрабатывал П. К. Анохин.
Однако системный подход продуктивен только в том случае, если мы более или менее определенно указываем на конкретные элементы, из которых состоит данная система, на функциональное значение каждого из этих элементов и на правила взаимодействия элементов друг с другом при функционировании системы в целом. Причем под «элементами» мы подразумеваем не гипотетические «блоки» логических схем, а реальные мозговые образования, ту приуроченность динамики к структуре, которую И. П. Павлов считал фундаментальным принципом своей материалистической теории. К сожалению, именно последнее требование оказалось критическим моментом для нейрофизиологического обоснования целого ряда очень красивых и логически стройных концепций. Иными словами, любая концепция, претендующая на объяснение принципов организации поведения, должна быть сопоставима с анатомическим строением мозга, поскольку и первое и второе формировались в русле единого эволюционного процесса.
В последние годы появились попытки выделить тот или иной конгломерат анатомически и функционально связанных между собой мозговых образований, к которому можно приурочить не отдельный фрагмент целенаправленного поведения, а достаточно универсальную поведенческую функцию. В. Д. Небылицин [1968, 1971] полагал, что нейроанатомической основой частных свойств нервной системы являются анализаторы (зрение, слух, осязание и т. д.). Что касается общих свойств, то их можно разделить на активность и эмоциональность. Первая зависит от индивидуальных особенностей функционирования лобно-ретикулярного комплекса мозговых структур, вторая - от лобно-лимбического комплекса. Р. Ю. Ильюченок выделяет эмоциогенную систему, включающую в себя следующие отделы головного мозга: миндалину, зону инсерта, пери- и паравентрикулярные ядра гипоталамуса и центральное серое вещество. Разрушение любого из этих образований, по данным автора, делает невозможным выработку условного рефлекса у крыс после одного сочетания, что свидетельствует о нарушении эмоционального компонента подкрепления [Hyutchenok, 1979]. Сходство дефекта, возникающего при повреждении любого звена комплекса мозговых структур, явилось основанием и для выделения «системы переднего-мозга», описанной В. А. Черкесом [1978]. В систему входят: ассоциативная кора, неостриатум (хвостатое ядро), гиппокамп и миндалина. «Это комплекс структур, ответственный за организацию тех форм поведения, которые являются наиболее сложными у данного вида животных» [Черкес, 1978, с. 147]. К сожалению, понятие о «наиболее сложной» для данного вида форме поведения слишком расплывчато.
Р. Дуглас [Douglas, 1972] предлагает различать две основные системы мозга: анализаторную (неокортекс, специфические ядра таламуса, гиппокамп, миндалина) и мотивационную (мозговой ствол, гипоталамус, неспецифические ядра таламуса). Автор подчеркивает значение взаимных влияний этих двух систем на деятельность друг друга. Однако и в этой схеме не уточняется роль каждой из перечисленных структур: ведь сказать, что система «анализирует важнейшие стимулы», явно недостаточно без уяснения принципов этого анализа. Нам кажется весьма спорным и отнесение миндалины к системе анализаторных структур, поскольку миндалина очень тесно связана с функционированием гипоталамуса.
В своих последних работах А. Р. Лурия [1978] описал три основных функциональных блока мозга: 1) блок регуляции тонуса и бодрствования (ретикулярная формация и лобные отделы коры); 2) блок приема, переработки и хранения информации (первичные, вторичные и третичные зоны коры); 3) блок программирования, регуляции и контроля деятельности (префронтальные отделы лобной коры). При анализе схемы функциональной организации мозга, предложенной А. Р. Лурия, возникает целый ряд вопросов. Какую роль в этой организации играют остальные субкортикальные образования, кроме ретикулярной формации? Каким образом функции «блоков» связаны между собой, то есть как процессы приема и переработки информации влияют на программирование деятельности, деятельность - на прием, переработку и хранение информации, первое и второе - на регуляцию тонуса и т. д., и т. п. Впрочем, схема А. Р. Лурия органически вытекает из его общих нейропсихологических представлений, откуда практически исключены потребности, мотивы и эмоции.
Что касается разделов учебников и руководств, посвященных описанию морфологического субстрата потребностей и эмоций, то они, как правило, состоят из простого перечисления эффектов, наблюдающихся при повреждении или электрической стимуляции передних отделов новой коры, гиппокампа, миндалины и гипоталамуса без указания на принципиальные особенности функций каждого из перечисленных образований, на специфику их «вклада» в организацию поведения. Мы полагаем, что причиной подобного положения вещей является отсутствие такой общей концепции интегративной деятельности мозга, которая могла бы быть приурочена к его анатомическому строению.
Наш собственный подход к проблеме, о которой идет речь, определила информационная теория эмоций, согласно которой потребности, действия и эмоции не только представляют самостоятельные, не отождествляемые друг с другом феномены высшей нервной (психической) деятельности, но с необходимостью должны иметь собственный морфофизиологический субстрат. Подобная точка зрения подтверждается результатами многих экспериментов.