

Модуль 1
Основи біохімії. Травлення та вплив харчових речовин на організм людини
Лекція 1
Тема: «Вступ. Харчування людини медико-біологічна та соціально-економічні проблеми»
План:
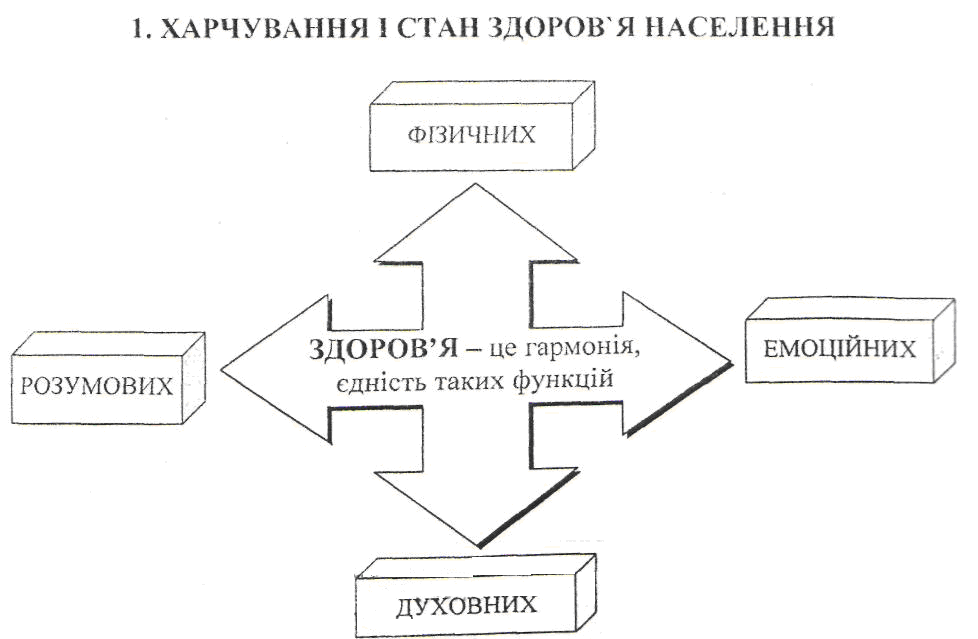
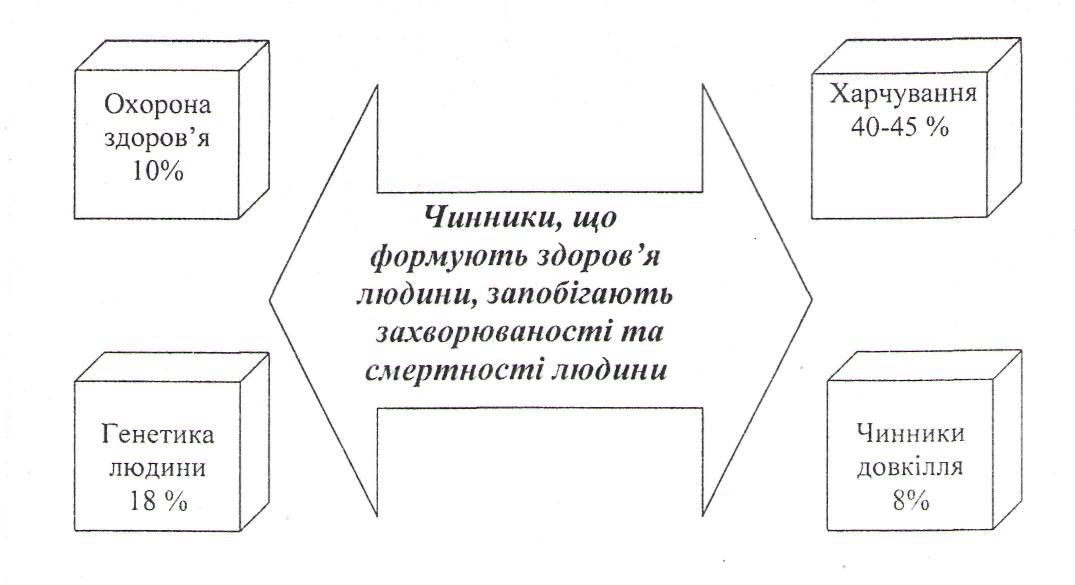
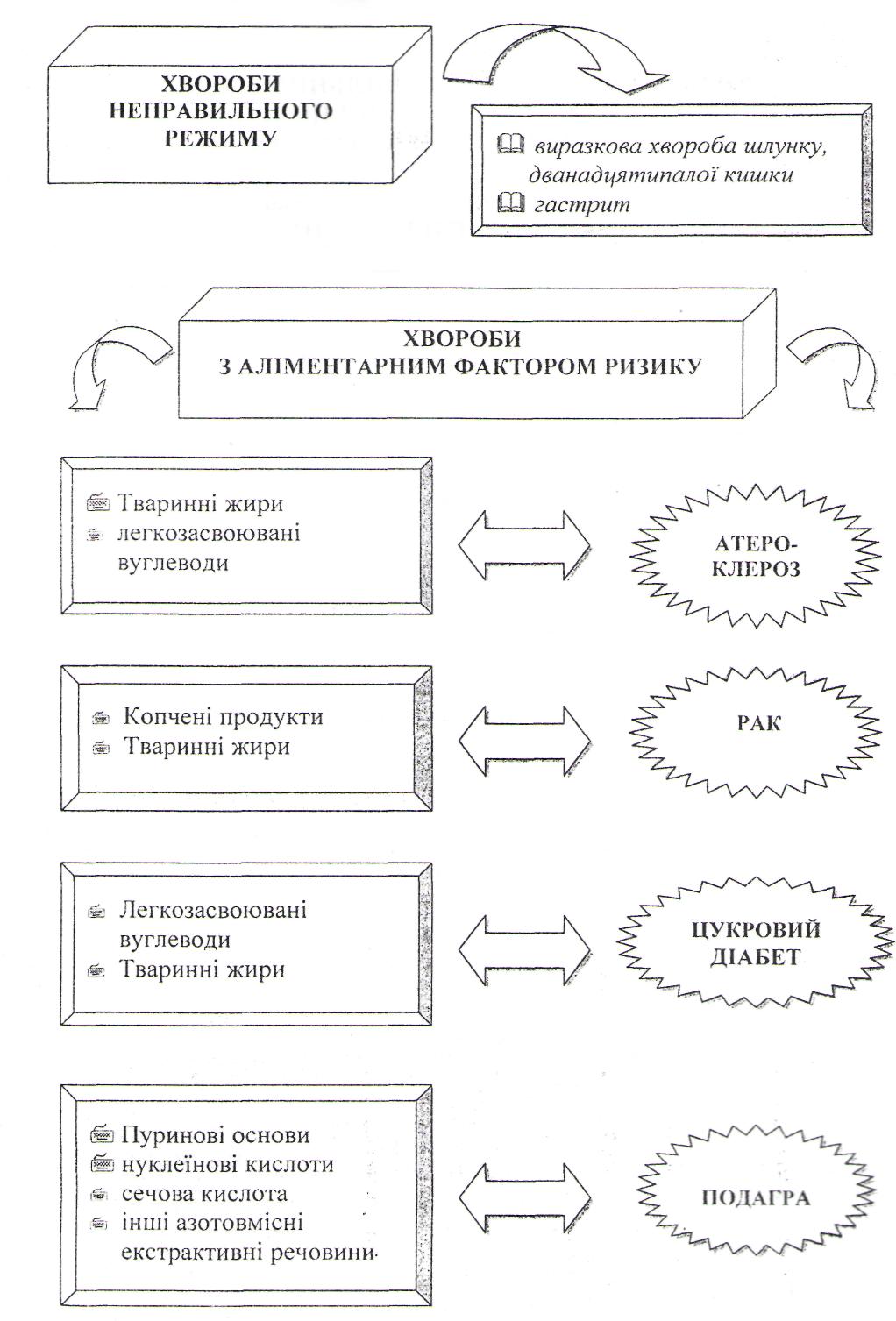
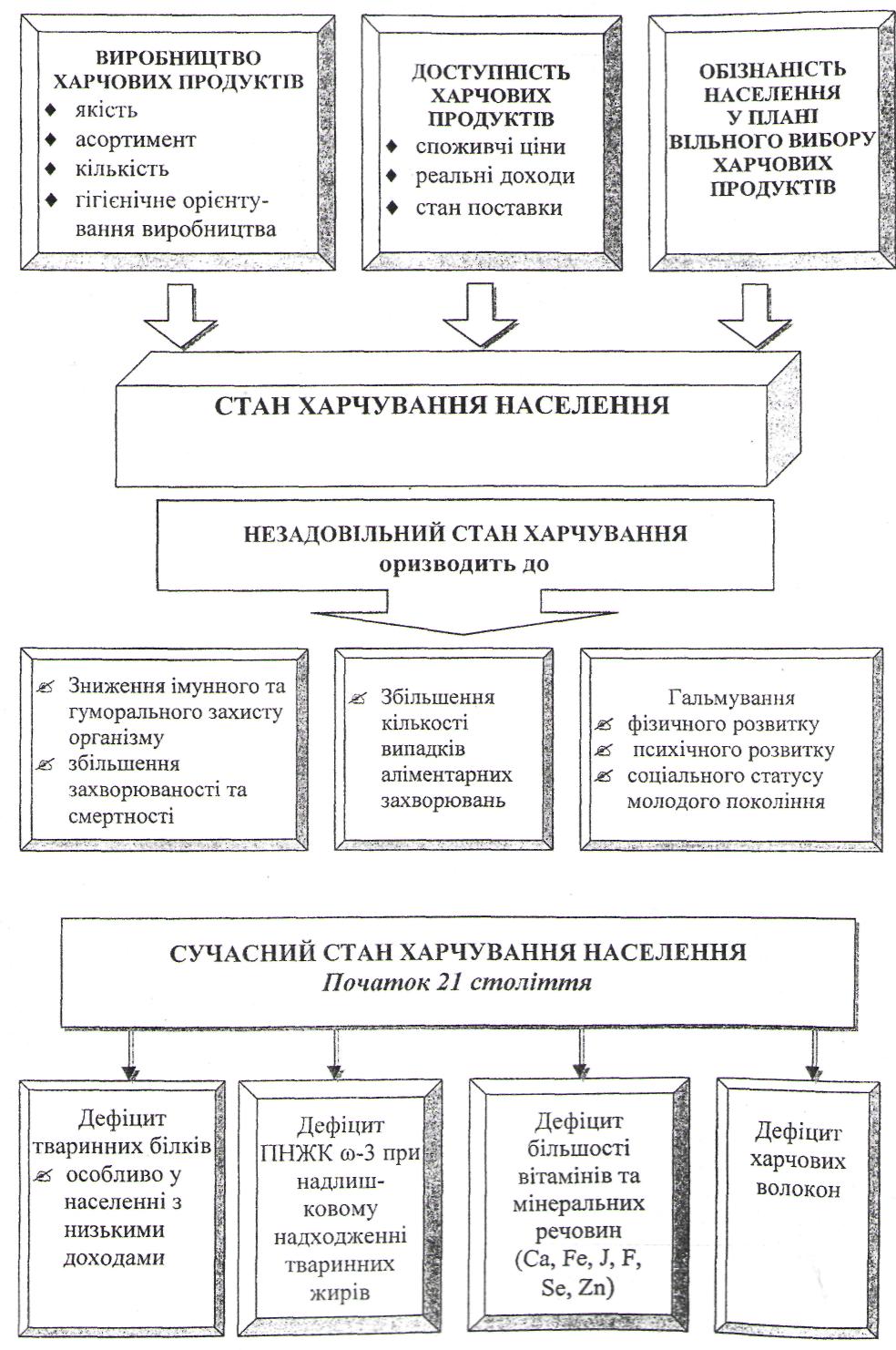
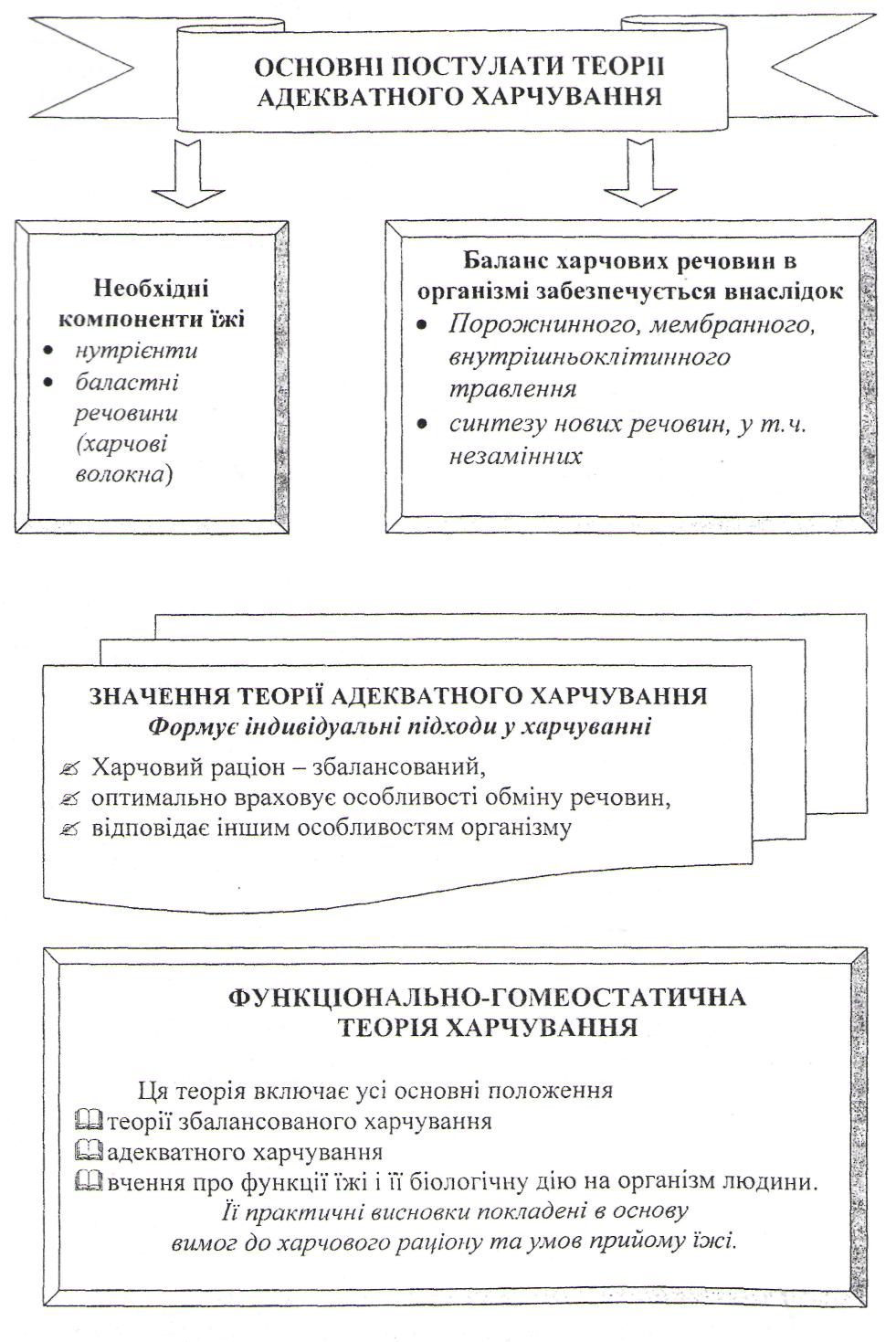
Харчування і стан здоров`я населення.
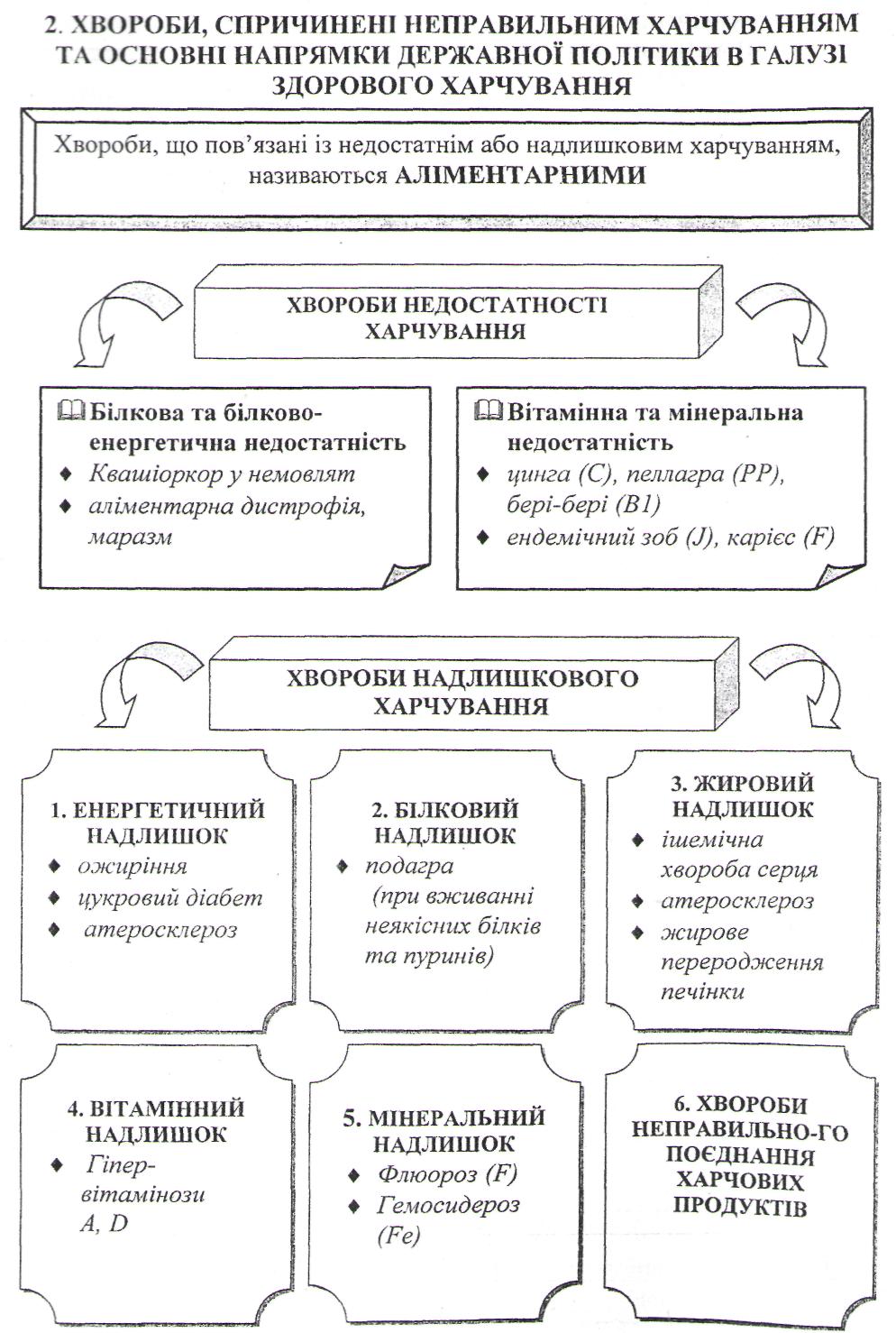
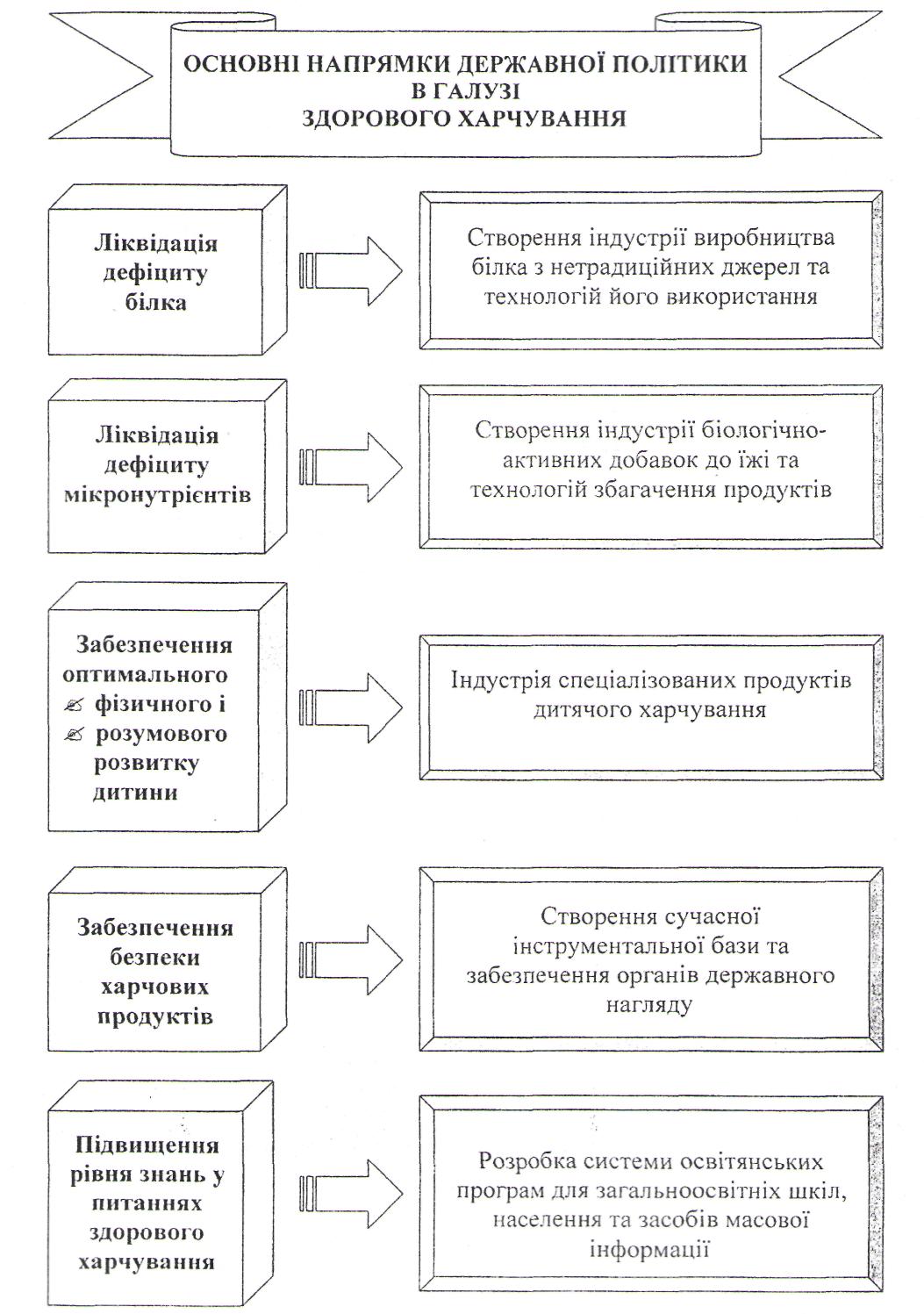
Хвороби, спричинені неправильним харчуванням та основні напрямки державної політики в галузі здорового харчування.
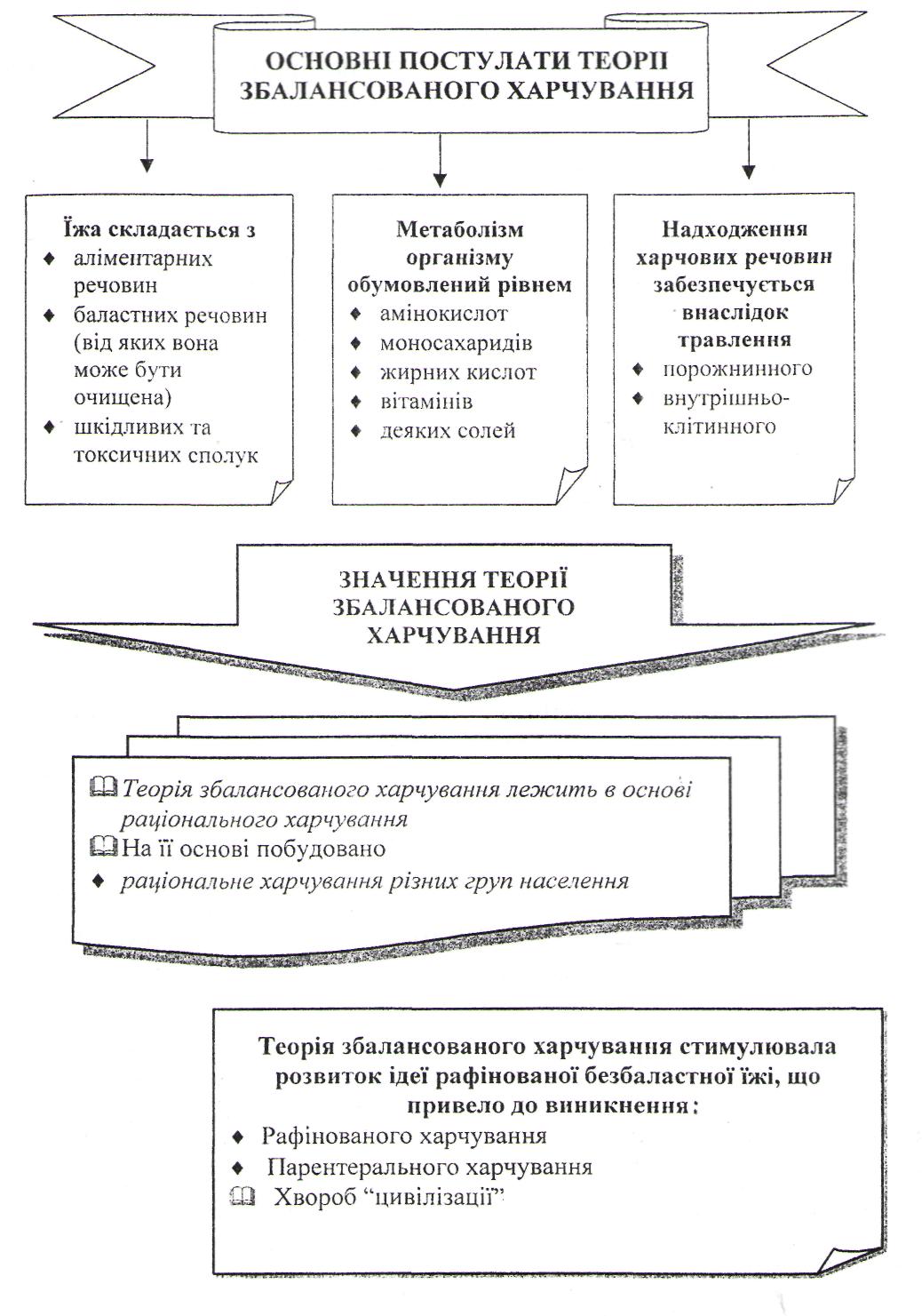
Теорії харчування та їх внесок у розвиток науки про харчування.
Функції їжі та фактори їх забезпечення.
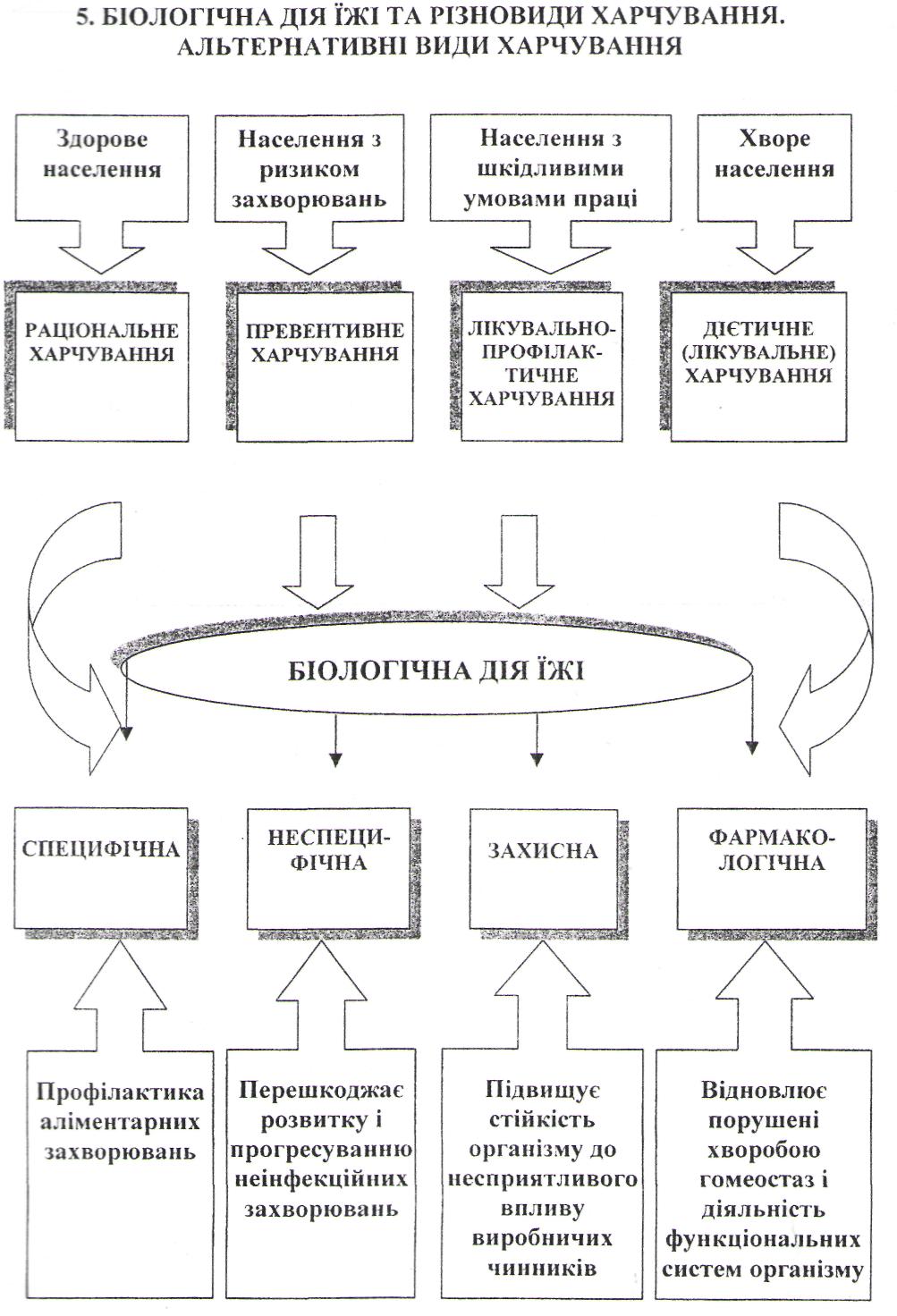
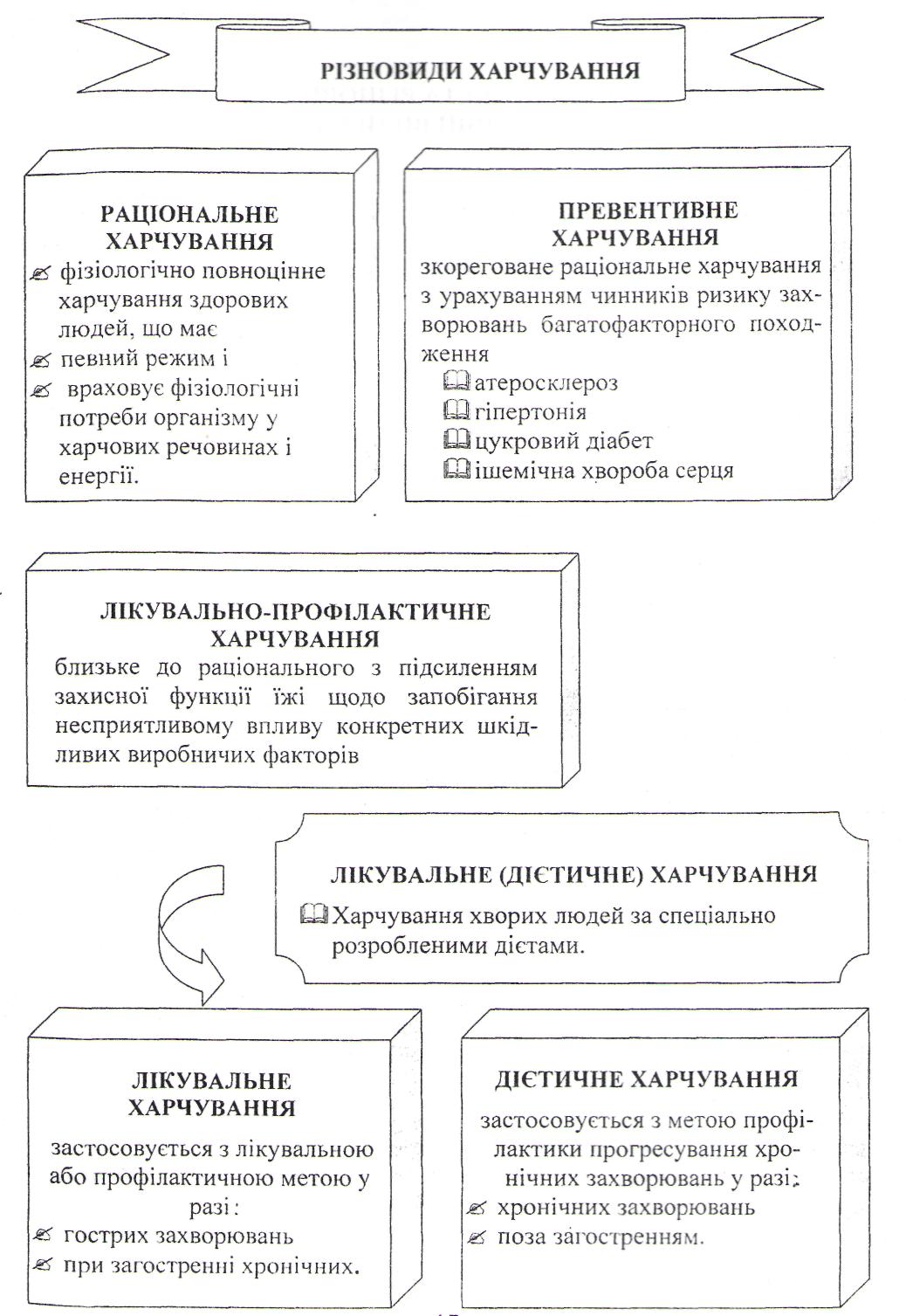
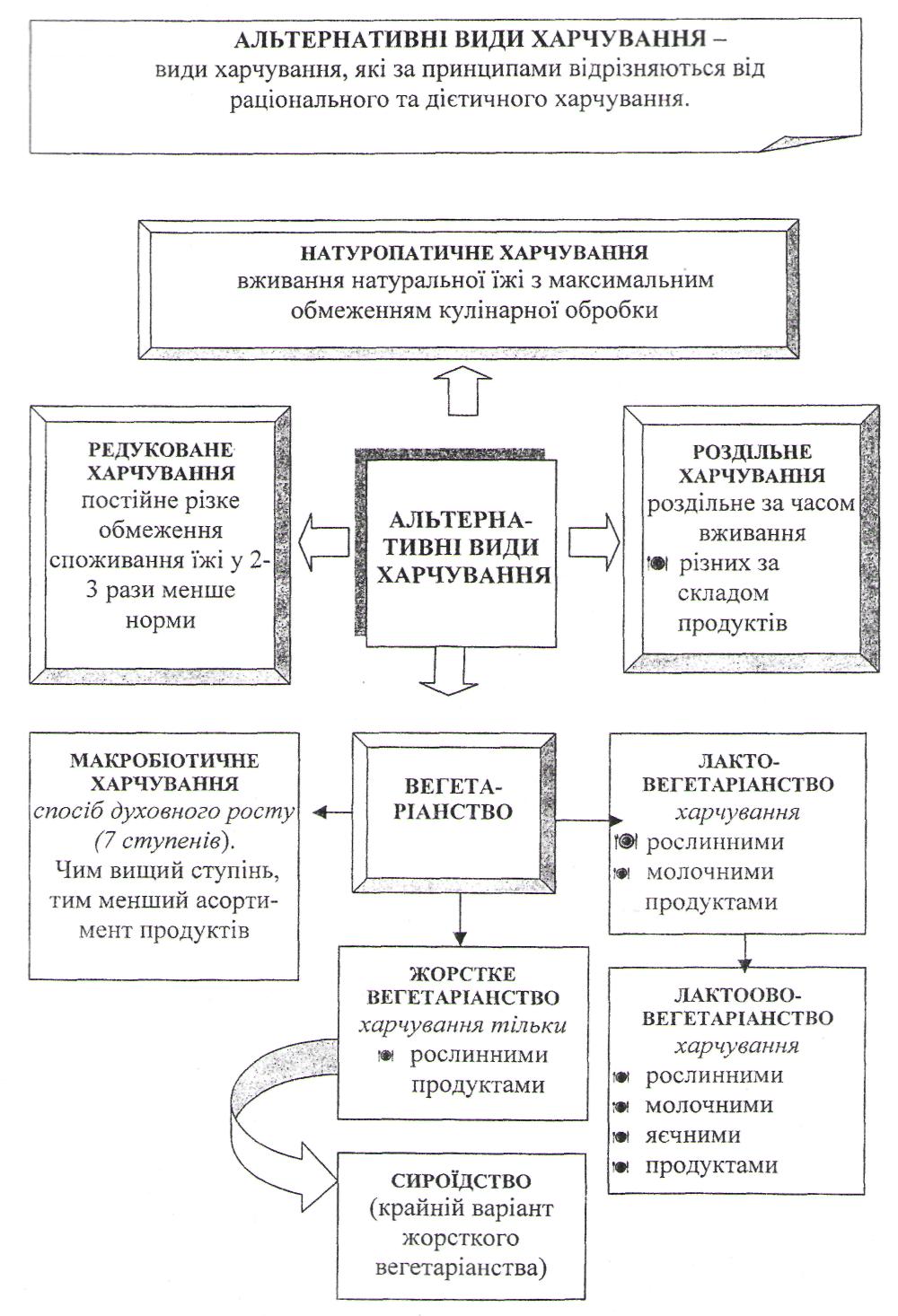
Біологічна дія їжі та різновиди харчування. Альтернативні види харчування.
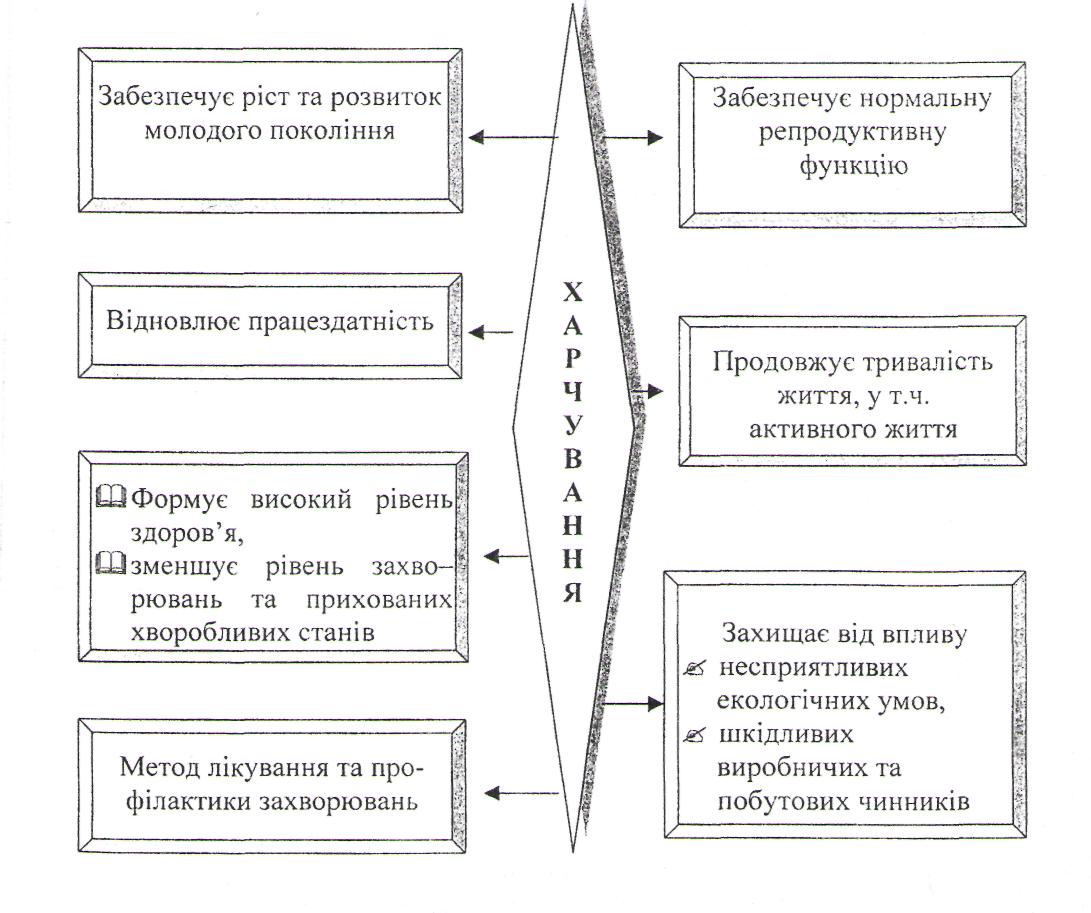



Функції їжі та фактори їх забезпечення.
1. Пластична функція. Її виконання забезпечується білками, ліпідами, вуглеводами, мінеральними речовинами, водою. Найбільш виражена пластична функція притаманна білкам, оскільки вони містяться у всіх органах і тканинах. Ліпіди виконують пластичну роль, оскільки входять до складу клітинних мембран, сполучної тканини, головного й спинного мозку. Таку саму роль відіграють деякі вуглеводи, наприклад, мукополісахариди, що входять до складу сполучної тканини, зв'язок, хрящів.
Пластичну функцію виконують також мінеральні речовини (Са, Р, Мg) у кістках, зубах. Ге входить до складу гемоглобіну і міоглобіну, І2— доскладу гормонів щитовидної залози, Мп необхідний для синтезу кісткової тканини, Си - структурна частина ферментних систем. Пластичну функцію забезпечують м'ясо і м'ясопродукти, риба і рибопродукти, молочні, яєчні продукти, що містять ці речовини.
Крім поживних речовин, у процесі асиміляції беруть участь також вітаміни, мінеральні речовини, гормони й інші біологічно активні сполуки, продукти життєдіяльності мікрофлори кишечника і чужерідні для організму речовини (ксенобіотики).
Збереження незмінності внутрішнього середовища є найважливішою умовою нормального обміну речовин в організмі. Навіть у разі випадкового вибору харчових продуктів, коли кількість і співвідношення нутрієнтів варіюють в значних межах, склад поживних речовин, що надходять у внутрішнє середовище, мало змінюється. У тонкій кишці поряд з транспортуванням речовин з її порожнини в кров існує постійний і спрямований у протилежний бік потік - із крові в порожнину.
Збільшення вмісту якого-небудь компоненту в раціоні позначається на всмоктуванні не лише цього, але й інших компонентів. Так, підвищення концентрації вуглеводів збільшує всмоктування всієї решти речовин.
Більше 80% маси амінокислот, щодня використовуваних організмом для синтезу білків, надходить не з їжею, а звільняється під час реакцій гідролізу власних білків.
Жоден харчовий продукт не містить усі без винятку життєво важливі компоненти, і лише їх різноманітний асортимент може повністю забезпечити організм необхідними нутрієнтами.
У нинішніх екологічних умовах, крім традиційних компонентів, у продуктах містяться різні домішки, до яких організм еволюційно не пристосувався. Наприклад, хімічні речовини, вживані в сільському господарстві для знищення бур'янів, шкідників, з їжею надходять в організм людини.
Вода, що входить до складу всіх органів і тканин, у,значній кількості утримується в крові. Зменшення її вмісту призводить до збільшення в'язкості крові, при цьому «розпалюється» питний центр, що відчувається як спрага. Організм хворобливо реагує на обезводнення.
Біорегуляторна (каталітична) функція. Здійснюється завдяки тому, що в їжі містяться попередники біологічно активних речовин. У регуляції метаболізму беруть участь амінокислоти (АК), вітаміни, поліненасичені жирні кислоти (ПНЖК) та ін. Так, з деяких амінокислот утворюються гормони, наприклад, із тирозина - йодвмісні гормони щитовидної залози, гормон наднирковвиків - адреналін і норадреналін - медіатор симпатичної нервової системи. Попередниками гормонів є деякі пептиди, зокрема, інсуліну (гормону підшлункової залози). Біологічно активними речовинами-ре- гуляторами можуть бути деякі ліпіди, зокрема, гормони кори надниркових залоз, які є похідними стеридів.
Каталітична роль їжі забезпечується і завдяки тому, що в харчових продуктах містяться вітаміни. Вони входять до складу ферментів, що знаходяться в тканинах живих організмів. Наприклад, вітамін РР присутній в анаеробній дегідрогеназі, вітамін В2 - в аеробній дегідрогеназі, вітамін В6 - складова частина ферментів- трансамінів, що здійснюють перенесення активних груп. Вітамін С бере участь в окиснювально-відновних процесах. Вираженими біо- регуляториими властивостями володіють різні овочі, фрукти, ягоди, яйця та ін., багаті на вітаміни.
3.Пристосовно-регуляторна функція. Цю роль їжа виконує завдяки харчовим волокнам, воді та іншим компонентам, що здійснюють регуляцію діяльності функціональних систем організму, найважливішими з яких є системи харчування, виділення і терморегуляції. Так, наприклад, харчові волокна регулюють моторну функцію кишечника, беруть участь у формуванні калових мас. Багаті на них хліб житній і пшеничний з борошна грубого помелу, висівки, крупи, картопля, овочі, фрукти.
4.Імунорегуляторна функція. Виявляється в дії на імуно-ком- петентні клітини, які впливають на здатність організму протистояти дії різних ушкоджувальних факторів. Вона залежить від якості харчування, особливо його білкового і вітамінного складу, вмісту ПНЖК (сімейства омега 3,6) і мікроелементів (Ре, Си, І2 та ін.).
5. Реабілітаційна функція. Полягає в зміні властивостей і хімічного складу раціону харчування з метою прискорення процесу одужання, запобігання рецидивів і переходу хвороби з гострої стадії в хронічну. Для цього використовуються різні групи дієтичних проектів, а саме - продукти з низьким вмістом натрію, з модифікова- им вуглеводним компонентом, зі зниженим вмістом жирів і поліп- іеним їх складом, зі зниженою енергетичною цінністю та ін.
6.Енергетична функція. Забезпечується завдяки компонен- ам, що містяться в їжі і в разі розщеплювання яких у тканинах рганізму виділяється енергія. Найбільша її кількість утворюєть- я під час розщеплювання засвоєних вуглеводів, ліпідів, органіч- гих кислот, етанолу. Менше значення як джерело енергії мають іілки. Надзвичайно важливе для збереження здоров'я дотримання іідповідності між кількістю тієї енергії, що надходить разом з їжею »організм людини, і тієї, що витрачається нею.
7.Сигнально-мотиваційна функція. Здійснюється смаковими і екстрактивними речовинами, що регулюють харчову мотивацію, тобто підтримують її на певному рівні. Основоположник наукової гігієни в Росії Ф.Ф. Зрисман писав: „Без смакових речовин в їжі ми померли б з голоду, але не від того, що їжа погано засвоюється, а від того, що ми швидко відмовилися б від усякої їжі". До смакових речовин належать приправи - оцет, гірчиця, кухонна сіль,цибуля, часник, кріп, селера, петрушка, лавровий лист, кардамон. Вони містять різні ефірні олії, органічні кисл ристі речовини, мінеральні елементи, вітаміни та інші спс надають їжі специфічного смаку й аромату.
Лекція 2.
Тема: «Травлення. Процес травлення живильних речовин в організмі людини».
У процесі травлення в шлунково-кишковому тракті ссавців три основних компоненти їжі - вуглеводи, жири і білки піддаються ферментативному гідролізові, розпадаючись при цьому на складові будівельні блоки, з яких вони утворені. Цей процес необхідний для утилізації харчових продуктів, оскільки клітки, що вистилають кишечник, здатні всмоктувати в кровоток тільки відносно невеликі молекули. Наприклад, засвоєння полісахаридів і навіть дисахаридів стає можливим тільки після їхнього повного гідролізу травними ферментами до моносахаридів. Аналогічним образом білки і ліпіди також повинні бути гідролізовані до блоків, з яких вони побудовані.
На рис. 1 приведена схема травної системи людини.
 Рисунок
1. – А. Шлунково-кишковий тракт людини
Рисунок
1. – А. Шлунково-кишковий тракт людини

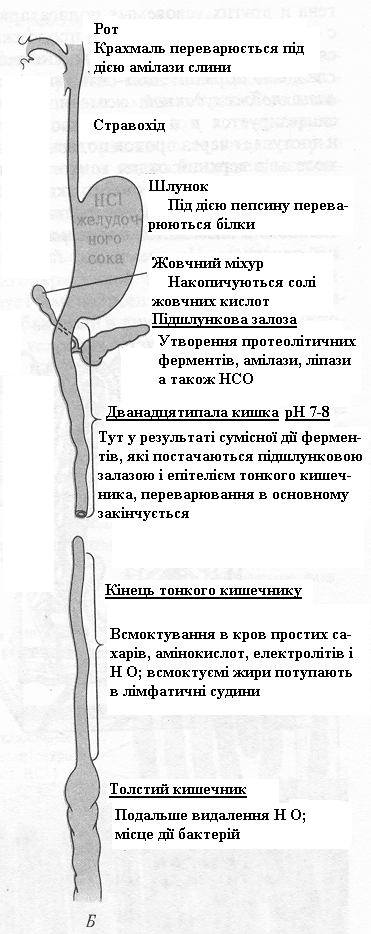
Рисунок 1. – Б. Послідовні етапи переварювання і всмоктування
Процес травлення починається з ротової порожнини і шлунка, тоді як кінцеві етапи переварювання всіх основних компонентів їжі й усмоктування в кров складових їхніх структурних блоків відбуваються в тонкому кишечнику. Анатомічно тонкий кишечник добре пристосований для виконання цієї функції, оскільки він має дуже велику площу поверхні, через яку відбувається усмоктування. Тонкий кишечник характеризується не тільки великою довжиною (4-4,5 м), але також наявністю на його внутрішній поверхні безлічі складок з великою кількістю пальцеподібних виступів, називаних ворсинками. Кожна ворсинка покрита епітеліальними клітками, що несуть численні мікро ворсинки (рис. 2).
 Рисунок
2. – А. Ворсинки слизової тонкого
кишечнику; видно як велика площина,
через яку всмоктуються продукти
травлення;
Рисунок
2. – А. Ворсинки слизової тонкого
кишечнику; видно як велика площина,
через яку всмоктуються продукти
травлення;
Рисунок 2. – Б. Мікрофотографія ворсинок, яка одержана за допомогою скануючого електронного мікроскопу; В,Г. Мікро- фотографія відповідно поздовжнього і поперечного зрізів ворсинок одержана за допомогою трансмісійного електронного мікроскопу
Ворсинки створюють величезну поверхню, через яку продукти переварювання швидко транспортуються в епітеліальні клітки, а з них у капіляри кровоносної системи й у лімфатичні судини, розташовані в стінці кишечнику. Площа поверхні тонкого кишечнику людини складає ~ 180 м2, тобто лише деяким менше ігрової площадки тенісного корту.
У мікроворсинках утримуються пучки актинових мікрофиламентів, з'єднаних у підставах мікроворсинок з мережею міозинових ниток. Ця система ниток забезпечує хвилеподібні коливання мікроворсинок, завдяки яким відбувається місцеве перемішування і краще усмоктування переварених живильних речовин.
а. Переварювання вуглеводів
У людини з вуглеводів переварюються в основному полісахариди - крохмаль і целюлоза, що утримуються в рослинній їжі, і глікоген, що утримується в їжі тваринного походження.
Крохмаль і глікоген цілком розщеплюються ферментами шлунково-кишкового тракту до складових їхніх структурних блоків, а саме вільної D-глюкози. Цей процес починається в роті під час пережовування їжі завдяки дії ферменту амілази, виділюваного слинними залозами. Амілаза слини гідролізує багато хто з (1→4) глікозидних зв'язків у крохмалі й у глікогені. При цьому утвориться суміш, що складається з мальтози, глюкози й олігосахаридів. Коли ми жуємо сухарі або крекери, вони стають поступово солодше, оскільки несмачний крохмаль, що утримується в них, піддається ферментативному гідролізові з утворенням цукрів. Переварювання крохмалю, глікогену й інших засвоюваних полісахаридів з утворенням D-глюкози продовжується і завершується в тонкому кишечнику, головним чином, під дією амілази підшлункової залози, що синтезується в підшлунковій залозі і надходить через протоку підшлункової залози у верхній відділ тонкого кишечнику. Цей відділ тонкого кишечнику з найбільш високою травною активністю називається дванадцятипалою кишкою.
Целюлоза в більшості ссавців не піддається ферментативному гідролізові і не використовується через відсутність ферментів, здатних розщеплювати β(1>4)-зв'язки між послідовними залишками D-глюкози в целюлозі. Разом з тим неперетравлена целюлоза з рослинної їжі створює ту масу (називану іноді «клітковиною» або «грубим кормом»), що сприяє нормальній перистальтиці кишечнику. У жуйних тварин целюлоза піддається переварюванню, але не прямим шляхом, а під дією бактерій, що знаходяться в їхньому рубці (шлунку). Ці бактерії гідролізують целюлозу до D-глюкози і далі сброжують D-глюкозу до лактату, ацетату
і пропіонату, що всмоктуються і надходять у кров. Далі лактат і пропіонат у печінці жуйних перетворюються в цукор крові.
Гідроліз дисахаридів каталізують ферменти, що знаходяться в зовнішньому краї епітеліальних кліток, що вистилають тонкий кишечник. Сахароза, або очеретяний цукор, гідролізується з утворенням D-глюкози і D-фруктози під дією сахарози, називаної також інвертазою; лактоза гідролізується до D-глюкози і D-галактози під дією лактази, називаної також β-галактозидазою; у результаті гідролізу мальтози під дією мальтази утворюються дві молекули D-глюкози. Нагадаємо, що багатьом представникам азіатських і африканських рас у дорослому стані властивий нестерпність лактози, обумовлена зникненням у їхньому тонкому кишечнику лактазної активності, що малася в грудному і дитячому віці. У людей з нестерпністю лактози цей цукор залишається в кишечнику в нерозщепленому виді і частина його піддається сброжуванню під дією мікроорганізмів. Це викликає діарею й утворення газів у кишечнику В епітеліальних клітках, що вистилають тонкий кишечник, D-фруктоза, D-галактоза і D-манноза частково перетворюються в D-глюкозу. Суміш усіх цих простих гексоз поглинається епітеліальними клітками, що вистилають тонкий кишечник, і доставляється кров'ю в печінку.
Лекція 3.
Тема: «Переварювання жирів та білків».
Білки їжі розщеплюються ферментами в шлунково-кишковому тракті до складових їхніх амінокислот. Білки, що надходять у шлунок, стимулюють виділення гормону гастрину, що у свою чергу викликає секрецію соляної кислоти обкладними клітками залоз слизуватої шлунка (рис.3), а також пепсиногену головними клітками.
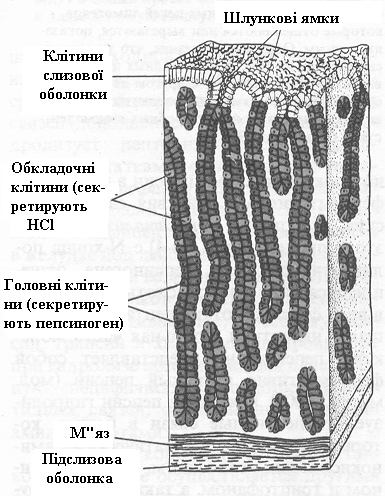
Рисунок 3. – Залози слизової шлунку. Обкладні клітини секретують HCl під дією гормону гастрину, який продуцюється епітеліальними клітками при надходженні у шлунок білків. Головні клітини секретують пепсиноген
Шлунковий сік має рН від 1,5 до 2,5. Завдяки такій кислотності він діє як антисептик, убиваючи більшість бактерій і інших кліток. Крім того, в умовах низького рН шлункового соку глобулярні білки піддаються денатурації, їхні молекули розвертаються і внаслідок цього внутрішні пептидні зв'язки поліпептидних ланцюгів стають більш доступними для ферментативного гідролізу. Пепсиноген (мол. маса 40000), що є неактивним попередником ферменту, або зимогеном, перетворюється в шлунковому соку в активний пепсин у результаті ферментативної дії самого пепсину, тобто шляхом автокаталізу. У ході цього процесу (рис.4)
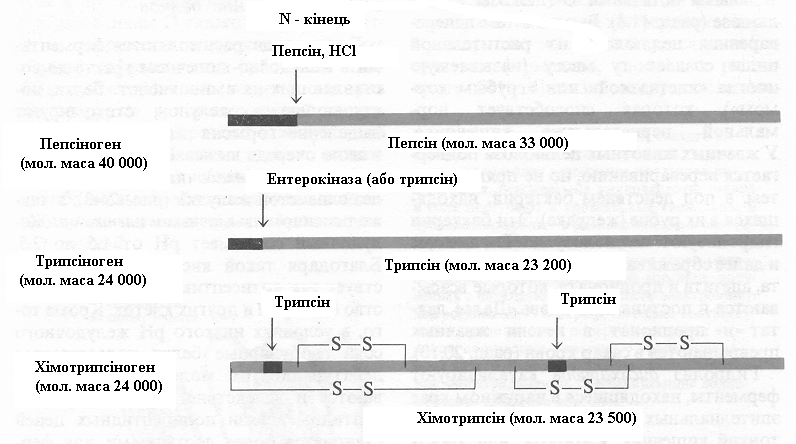 Рисунок
4. – Активація зимогенів пепсину, трипсину
і хімотрипсину. На діаграмах показані
ділянки зимогенів, які підлягають
протеолізу, в результаті якого звільнюються
активні ферменти (показані сірим). Ті
фрагменти поліпептидних ланцюгів
зимогенів, котрі відщепляються або
вирізаються, показані чорним.
Рисунок
4. – Активація зимогенів пепсину, трипсину
і хімотрипсину. На діаграмах показані
ділянки зимогенів, які підлягають
протеолізу, в результаті якого звільнюються
активні ферменти (показані сірим). Ті
фрагменти поліпептидних ланцюгів
зимогенів, котрі відщепляються або
вирізаються, показані чорним.
З N-кінця поліпептидного ланцюга пепсиногену від-щеплюються 42 амінокислотних залишки у виді суміші коротких пептидів. Інша частина молекули пепсиногену що залишається інтактною являє собою ферментативно активний пепсин (мол. маса 33000). У шлунку пепсин гідролізує по пептидним зв'язкам у білках, які утворені ароматичними амінокислотами - тирозином, фенілаланіном і триптофаном, а також поруч інших (табл. 1); у підсумку з довгих поліпептидних ланцюгів утвориться суміш більш коротких пептидів.
Як тільки кислий вміст шлунка попадає в тонкий кишечник, у ньому під впливом низького pН починається секреція гормону секретину, що надходить у кров. Цей гормон у свою чергу стимулює виділення з підшлункової залози в тонкий кишечник бікарбонату, що приводить до нейтралізації HCl шлункового соку. B результаті pН різко зростає від 1,5-2,5, до ~ 7. B тонкому кишечнику перева-рювання білків продовжується. Надходження амінокислот у дванадцятипалу кишку викликає звільнення гормону холецистокініну, що стимулює секрецію декількох ферментів підшлункової залози з оптимумом рН близько 7.
Таблиця 1. - Ферменти, що беруть участь у переварюванні білків, і їхня специфічність по відношенню до пептидних зв'язків, утворених різними амінокислотними залишками
Пепсин, Tyr, Phe, Тrр,
а також Leu, Glu, Gln
Трипсин Lys, Arg
Хімотрипсин Туr, Phe, Трr
Карбоксипептидаза Послідовне відщіплення
С-кінцевих залишків
Амінопептидаза Послідовне відщіплення
N-кінцевих залишків
Три з них - трипсин, хімотрипсин і карбокси-пептидаза - виробляються екзокринними клітками підшлункової залози (рис.5) у виді ферментативно неак-тивних зимогенів - відповідно трипсиногену, хімотрипсиногену і npoкарбоксипептидази.
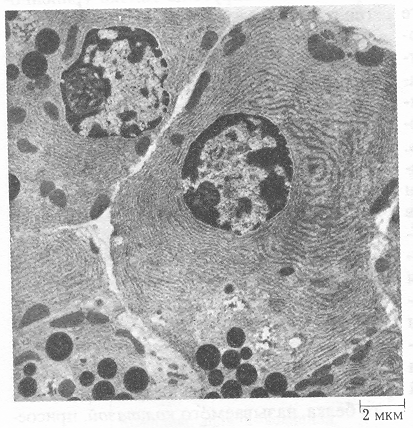
Рисунок 5.- Екзокринні клітини підшлункової залози.
Цитоплазма клітин цілком заповнена шорсткуватим ендоплазматичним ретикулумом. Рибосоми на його мембранах синтезують поліпептидні ланцюги зимогенів багатьох травних ферментів. Зимогени накопичуються у вакуолях, що перетворюються зрештою в зрілі зимогенні гранули. При стимуляції клітки її плазматична мембрана зливається з мембраною, яка оточує зимогенні гранули; останні видні в нижній частині рисунка у виді темних часток сферичної форми. Вміст гранул вивільняється в просвіт протоки (світла область у нижній лівій частині рисунка) за допомогою екзоцитоза. Окремі протоки в остаточному підсумку ведуть у загальну протоку підшлункової залози і далі в тонку кишку.
Завдяки синтезові протеолітичних ферментів у виді неактивних попередників екзокринні клітини не піддаються руйнуванню цими ферментами. Потрапивши в тонкий кишечник, трипсиноген перетворюється на активну форму - трипсин (рис.4) - під дією ентерокінази, спеціалізованого протеолітичного ферменту, який секретується клітинами кишкового епітелію. Вільний трипсин у міру свого утворення також бере участь у каталітичному перетворенні трипсиногену на трипсин. Утворення вільного трипсину обумовлено відщіпленням гексапептидази від N-кінця поліпептидного ланцюга трипсиногену. Як ми вже бачили, трипсин гідролізує пептидні зв'язки, утворені за участю карбонільних груп лізину й аргініну (табл.1).
Молекула хімотрипсиногену являє собою один поліпептидний ланцюг з декількома внутрішньо-ланцюговими дисульфідними зв'язками. Потрапивши в тонкий кишечник, хімотрипсиноген перетворюється на хімотрипсин під дією трипсину, що розриває довгий специфічність по відношенню до пептидних зв'язків, утворених різними амінокислотами.
Деградація коротких пептидів у тонкому кишечнику здійснюється іншими пептидазами. До них відноситься в першу чергу карбоксипептидаза - цинквмісний фермент, синтезований у підшлунковій залозі у виді неактивного зимогену прокарбоксипептидази. Карбоксипептидаза послідовно відщеплює від пептидів С-кінцеві залишки. Тонкий кишечник секретує також амінопептидазу, яка відщеплює від коротких пептидів один за іншим N-кінцеві залишки (табл.1). B результаті послідовної дії цих протеолітичних ферментів і пептидаз перетравлювані білки в остаточному підсумку перетворюються на суміш вільних амінокислот, що далі транспортуються через епітеліальні клітини, які покривають тонкі кишки. Вільні амінокислоти проникають у капіляри ворсинок і переносяться кров'ю в печінку.
B шлунково-кишковому тракті людини не усі білки переварюються цілком. Більшість тваринних білків майже цілком гідролізуються до амінокислот, однак ряд фібрилярних білків, наприклад кератин, переварюється тільки частково. Багато білків рослинної їжі, зокрема білки зерен злаків, не цілком розщеплюються в силу того, що білкова частина насінь і зерен покрита не перетравлюваною целюлозною оболонкою (лушпайкою).
Відомо рідке захворювання cmeamopея (завзятий понос), при якому ферменти кишечнику не здатні переварювати визначені водорозчинні білки зерна, зокрема гліадин, що ушкоджує епітеліальні клітки кишечнику. Зрозуміло, що з їжі таких хворих варто виключити зернові продукти. Іншим захворюванням, зв'язаним з відхиленням поліпептидний ланцюг хімотрипсиногену в двох місцях, вистригаючи діпептиди (рис.4). Три фрагменти, що утворилися з вихідного ланцюга хімотрипсиногену, утримуються, однак, разом за допомогою перехресних дисульфідних зв'язків. Хімотрипсин гідролізує пептидні зв'язки, утворені залишками фенілаланіну, тирозину і триптофану (табл.1). Отже, трипсин і хімотрипсин розщеплюють поліпептиди, що утворилися в шлунку під дією пепсину, на пептиди меншої величини. Цей етап переварювання білків протікає з дуже високою ефективністю, оскільки пепсин, трипсин і хімотрипсин виявляють під час гідролізу поліпептидних ланцюгів різну від норми активності протеолітичних ферментів травного тракту, є гострий панкреатит. При цьому захворюванні,
18 обумовленому порушенням процесу виділення соку підшлункової залози в кишечник, попередники протеолітичних ферментів (зимогени) перетворюються на відповідні каталітично активні форми занадто рано у середині кліток підшлункової залози.
B результаті ці могутні ферменти впливають на тканину самої залози, викликаючи глибоке і дуже хворобливе руйнування органу, що може привести до смертельного результату. B нормі зимогени, які виділені підшлунковою залозою, не активуються доти, поки не потраплять у тонкий кишечник. Підшлункова залоза захищається від самопереварювання і іншим шляхом: у ній синтезується особливий білок - специфічний інгібітор трипсину. Оскільки вільний трипсин активує не тільки трипсиноген і хімотрипсиноген, але також і зимогени двох інших травних ферментів - прокарбоксипептидазу і проеластазу, інгібітор трипсину успішно запобігає передчасне утворення вільних протеолітичних ферментів у клітках підшлункової залози.
Переварювання жирів
П ереварювання
триацилгліцеролів (нейтральних жирів)
починається в тонкому кишечнику, куди
з підшлункової залози надходить зимоген
проліпаза.
Тут
проліпаза перетворюється на активну
ліпазу, що у присутності жовчних
кислот (див.
нижче) і спеціального білка, називаного
коліпазою,
приєднується
до крапельок триацилгліцеролів і
каталізується гідролітичне відщіплення
одного або обох крайніх жирнокислотних
залишків з утворенням суміші вільних
жирних кислот у виді їх Na +
- або К+-солей
(милів) і 2-моноацилгліцеролів (рис.6).
ереварювання
триацилгліцеролів (нейтральних жирів)
починається в тонкому кишечнику, куди
з підшлункової залози надходить зимоген
проліпаза.
Тут
проліпаза перетворюється на активну
ліпазу, що у присутності жовчних
кислот (див.
нижче) і спеціального білка, називаного
коліпазою,
приєднується
до крапельок триацилгліцеролів і
каталізується гідролітичне відщіплення
одного або обох крайніх жирнокислотних
залишків з утворенням суміші вільних
жирних кислот у виді їх Na +
- або К+-солей
(милів) і 2-моноацилгліцеролів (рис.6).
Невелика кількість триацилгліцеролів залишається при цьому негідролізованим.
Мила, що утворилися, і нерозщеплені ацилгліцероли емульгуються у виді дрібних крапельок під дією перистальтики (перемішуючі рухи кишечнику), а також під впливом солей жовчних кислот і моноацилгліцеролів, що є амфіпатичними сполуками і тому функціонують як детергенти. Жирні кислоти і моноацилгліцероли з цих крапельок поглинаються кишковими клітками, де з них в основному знову синтезуються триацилгліцероли. Далі триацилгліцероли проникають не в капіляри крові, а в невеликі лімфатичні судини кишкових ворсинок - лактеалі (інакше хільозні судини). Лімфа, що відтікає від тонких кишок, після переварювання жирної їжи нагадує по виду молоко через достаток зважених у ній хіломікронів - дрібних крапельок емульгованих триацилгліцеролів діаметром близько 1 мкм. Хіломікрони мають гідрофільну оболонку, що складається з фосфоліпідів і спеціального білка, що утримує хіломікрони в зваженому стані. Хіломікрони проходять через грудну протоку в підключичну вену (рис.7).
Емульгуванню і переварюванню ліпідів у тонкому кишечнику сприяють солі жовчних кислот. Солі жовчних кислот людини - це в основному глікохолат натрію і таурохолат натрію, обидві вони є похідними холевої кислоти (рис. 8), що кількісно переважає серед чотирьох основних жовчних кислот, які присутні в організмі людини.
Солі жовчних кислот є могутніми емульгаторами; вони надходять з печінки в жовч, що виливається у верхній відділ тонкого кишечнику. Після завершення усмоктування жирних кислот і моноацилгліцеролів з емульгованих крапельок жиру в нижньому відділі тонкого кишечнику відбувається зворотне усмоктування також і солей жов-чних кислот, що сприяли цьому процесові. Вони повер-таються в печінку і використовуються вдруге. Таким чином, жовчні кислоти постійно циркулюють між печінкою, і тонким кишечником (рис. 7).
Рисунок 7. – Схематичне зображення основних етапів
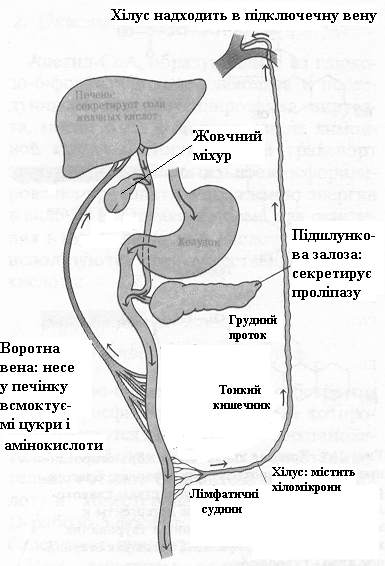
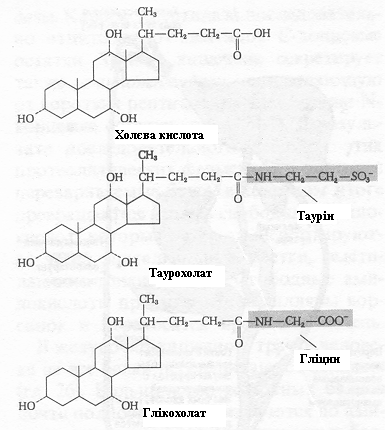
Рисунок 8. – Холева кислота та її коньюговані форми – таурохолат і глікохолат.
Жовчні кислоти виграють винятково важливу роль у засвоєнні не тільки триацилгліцеролів, але і взагалі всіх жиророзчинних компонентів їжі. Якщо жовчні кислоти утворюються або секретуються в недостатній кількості, як це має місце при ряді захворювань, то непереварені і не- поглинені жири з'являються в калі. При цьому погіршується усмоктування жиророзчинних вітамінів A, D, E і K і може виникнути харчова недостатність вітаміну A.
Лекція 4.
Тема: « Роль печенкі у процесі травлення»
Печінка здійснює обробку і розподіл живильних речовин. За винятком більшої частини триацилгліцеролів, живильні речовини, поглинені в кишковому тракті, надходять безпосередньо в печінку - основний центр розподілу живильних речовин у хребетних. Тут цукор, амінокислоти і деякі ліпіди піддаються подальшим перетворенням і розподіляються між різними органами і тканинами. Подивимося, як же відбувається інтеграція шляхів метаболізму основних живильних речовин у печінці. B печінці мається п'ять шляхів метаболізму цукрі. Велика частина спожитої вільної D-глюкози в печінці фосфорілюється за допомогою ATP з утворенням глюкозо-6-фосфату. Поглинені в тонкому кишечнику D-фруктоза, D-галактоза і D-манноза також перетворюються на D-глюкозо-6-фосфат у результаті ферментативного процесу, розглянутого раніше. D-глюкозо-6-фосфат лежить, таким чином, на перехресті всіх шляхів перетворення вуглеводів у печінці. Метаболізм цієї сполуки в печінці може здійснюватися по п'ятьох основних напрямках, і вибір якого-небудь одного з них залежить від щогодиних і навіть щохвилиних мінливих «попиту та пропозиції» (рис. 9).
а. Перетворення в глюкозу крові
Глюкозо-6-фосфат може дефосфорилюються під дією глюкозо-6-фосфатази з утворенням вільної D-глюкози, що надходить у кровоток і доставляється в інші тканини. Цей шлях перетворення глюкозо-6-фосфату має першорядне значення, оскільки концентрація глюкози в крові повинна підтримуватися на досить високому рівні, необхідному для забезпечення енергією мозку й інших тканин.
Рисунок 9. – Шляхи перетворення глюкозо-6-фосфату у печінці
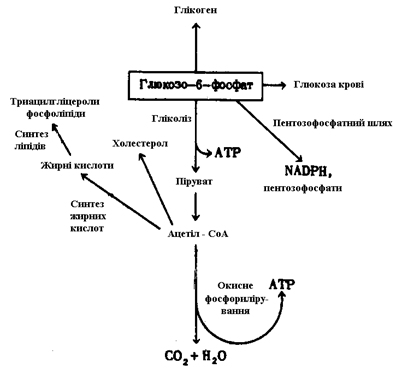
б. Перетворення в глікоген
Tа кількість глюкозо-6-фосфату, що не була використана для негайного перетворення на глюкозу крові, перетворюється на глікоген у результаті послідовної дії фосфоглюкомутази і глікоген-синтази.
в. Перетворення на жирні кислоти і холестерол
Надлишок глюкозо-6-фосфату, не використаного для утворення глюкози крові або глікогену печінки, розпадається в ході гліколізу і наступної дії піруватдегідрогенази до ацетил-СоА, що перетворюється на малоніл-СоА і далі на жирні кислоти. Жирні кислоти йдуть на утворення триацилгліцеролів і фосфоліпідів, що частково експортуються в інші тканини, куди їх переносять ліпопротеіни плазми. Визначена частка ацетил-СоА в печінці йде на синтез холестерола.
г. Окисний розпад до CO2
Ацетил-СоА, що утворився з глюкозо-6-фосфату в ході гліколізу і наступного декарбоксилювання пірувату, може бути окислений у циклі лимонної кислоти. Наступний транспорт електронів і окисне фосфорилювання приводять до накопичення енергії у виді ATP. Однак у нормі для окислювання в циклі лимонної кислоти в печінці використовуються переважно жирні кислоти.
д. Розпад по пентозофосфатному шляху
Глюкозо-6-фосфат служить субстратом пентозофосфатного шляху, у ході якого утворюються: 1) NADPH - відновник, необхідний для відбудовних етапів біосинтезу жирних кислот і холестеролу і 2) D-рібозо-5-фосфат - попередник біосинтезу нуклеотидів.
Під дією різних регуляторних ферментів, а також гормонів у печінці відбувається розподіл потоків глюкозних залишків по переліченим вище шляхах у залежності від співвідношення між потребами організму і надходженням вуглеводів з їжею.
Амінокислоти, що всмокталися в кишечнику і надійшли потім у печінку, також мають кілька основних шляхів метаболізму (рис. 10).
Рисунок 10. – Шляхи перетворення амінокислот в печінці
а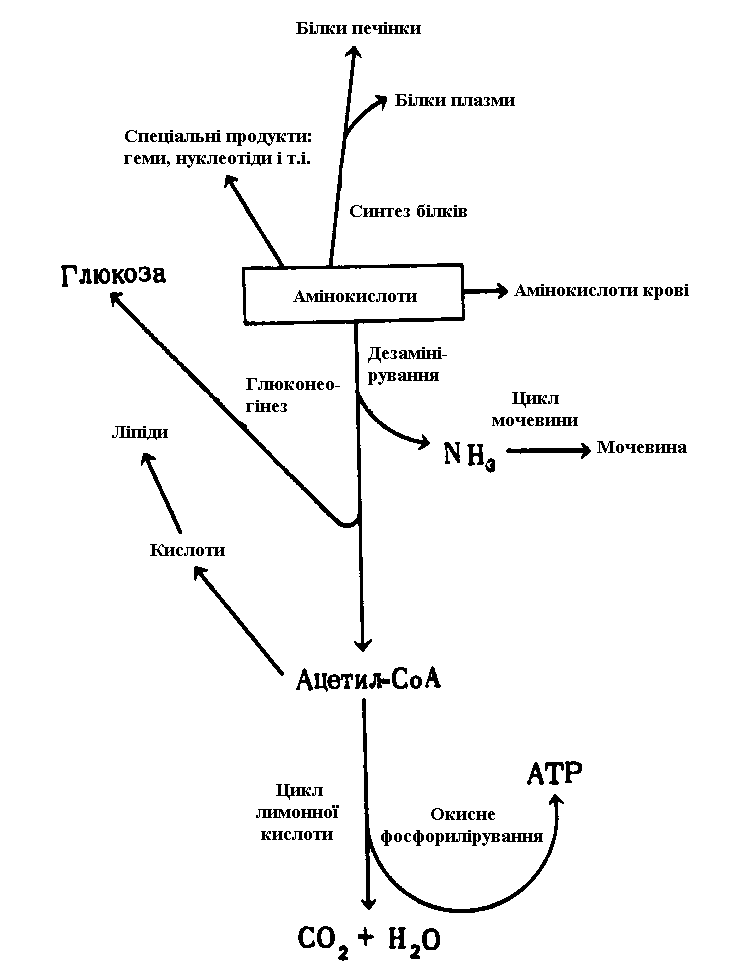 .
Транспорт в інші тканини
.
Транспорт в інші тканини
Амінокислоти з печінки можуть надходити в систему кровообігу й у такий спосіб доставлятися в інші органи, а там використовуватися як будівельні блоки для біосинтезу тканинних білків.
б. Біосинтез білків печінки і плазми крові
Білки печінки піддаються постійному відновленню, причому для них характерна дуже висока швидкість обороту із середнім періодом напівжиття усього лише в кілька днів. Крім того, саме в печінці синтезується більшість білків плазми крові.
в. Дезамінування і розпад
Tі амінокислоти, що не були використані в печінці або в інших органах для біосинтезу білків, піддаються дезамінуванню і розпадаються з утворенням ацетил-СоА і проміжних субстратів циклу лимонної кислоти. Останні можуть перетворитися на глюкозу і глікоген шляхом глюконеогенезу. Ацетил-СоА або піддається окислюванню в циклі лимонної кислоти з накопиченням енергії, що запасається у формі ATP, або перетворюється на ліпіди, що, як було описано вище, відкладаються в запас. Аміак, що вивільняється при розпаді амінокислот, перетворюється на продукт екскреції - сечовину в ході циклу, що протікає в печінці циклу сечовини.
г. Участь у циклі глюкоза - аланін
Печінка бере участь також у метаболізмі амінокислот, що надходять час від часу з периферічних тканин. Через декілька годин після кожного прийому їжі з м'язів у печінку надходить аланін; у печінці він піддається дезамінуванню, а піруват що утворився у результаті глюконеогенезу перетворюється на глюкозу крові. Глюкоза повертається в кістякові м'язи для заповнення в них запасів глікогену. Одна з функцій цього циклічного процесу, називаного циклом глюкоза-аланін, полягає в тому, що він зм'якшує коливання рівня глюкози в крові в період між прийомами їжі. Відразу після переварювання й усмоктування вуглеводів їжі, а також після перетворення частини глікогену печінки в глюкозу, в кров надходить достатня кількість глюкози. Але у період, що передує черговому прийомові їжі, відбувається частковий розпад м'язових білків до амінокислот, що шляхом переамінування передають свої аміногрупи на продукт гліколізу піруват з утворенням аланіну. Таким чином, у виді аланіну в печінку доставляється і піруват, і NH3. B печінці аланін піддається дезамінуванню, піруват що утворився перетворюється на глюкозу, що надходить у кров, а NH3 включається до складу сечовини і виводиться з організму. Виниклий у м'язах дефіцит амінокислот надалі після їжі заповнюється за рахунок усмоктуваних амінокислот їжі.
д. Перетворення на нуклеотиди й інші продукти
Амінокислоти служать попередниками в біосинтезі пуринових і піримідинових основ нуклеотидів, а також у синтезі деяких спеціалізованих речовин, зокрема порфиринів, гормонів і інших азотвмісних сполук. Ці шляхи перетворень амінокислот підсумовані на рис. 10.
Існує п'ять шляхів перетворення ліпідів
Як показано на рис. 11, для жирних кислот, що входять до складу ліпідів, також характерно кілька шляхів метаболізму в печінці.
Рисунок 11. – Обмін жирних кислот в печінці
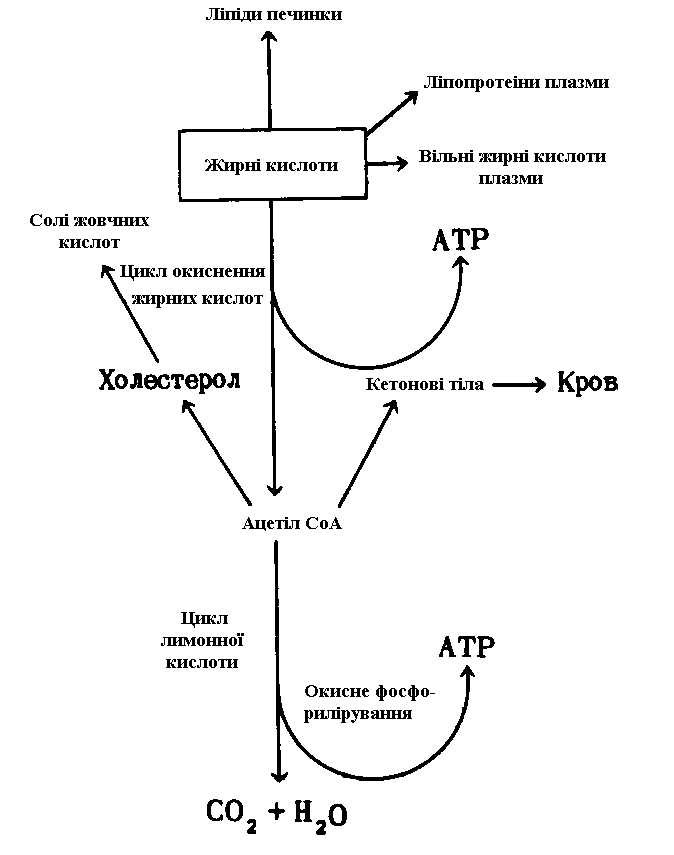
а. Окислювання до CO2 з утворенням ATP
Вільні жирні кислоти піддаються активації й окислюванню з утворенням ацетил-СоА і ATP. Далі ацетил-СоА окислюється в циклі лимонної кислоти, і в ході окисного фосфорилювання утвориться ATP. Жирні кислоти служать основним субстратом енергетичного обміну в печінці.
б. Утворення кетонових тіл
Надлишок ацетил-СоА, вивільнений при окислюванні жирних кислот і не використаний печінкою, перетворюється на кетонові тіла - ацетоацетат і D-β-гідроксибутират, - які переносяться кров'ю в периферічні тканини, де використовуються для окислювання в циклі лимонної кислоти. Кетонові тіла можна розглядати як транспортну форму ацетильних груп. Їхній внесок в енергетику периферічних тканин може бути дуже значним: у серце, наприклад, вони задовольняють до однієї третини енергетичних потреб.
в. Біосинтез холестеролу і жовчних кислот
Частина ацетил-СоА, утвореного з жирних кислот (і з глюкози), служить основним попередником у біосинтезі холестеролу, що у свою чергу є попередником жовчних кислот, необхідних для переварювання й усмоктування жирів.
г. Біосинтез ліпопротеінів плазми крові
Жирні кислоти служать також попередниками в синтезі ліпідної частини ліпопротеінів плазми крові. Ліпопротеіни функціонують як переносники ліпідів у жирову тканину, де останні накопичуються у виді триацилгліцеролів.
д. Утворення вільних жирних кислот плазми крові
Вільні жирні кислоти зв'язуються із сироватковим альбуміном і далі доставляються кров'ю в серце і кістякові м'язи; ці органи поглинають і використовують жирні кислоти в якості основного «палива».
Таким чином, метаболізм печінки має виняткову гнучкість і широкий діапазон. Печінка добре пристосована для виконання функції «розподільного центру» в організмі. Вона забезпечує доставку необхідних кількостей живильних речовин в інші органи, згладжує коливання в обміні речовин, обумовлені нерівномірністю надходження їжі, а також здійснює перетворення надлишку аміногруп у сечовину й інші продукти, що виводяться нирками.
Крім перетворення і розподілу вуглеводів, жирів і амінокислот у печінці активно протікають процеси ферментативної детоксикації сторонніх органічних сполук, наприклад лік, харчових добавок, консервантів і інших потенційно шкідливих речовин, що не мають харчової цінності. Детоксикація звичайно полягає в тому, що відносно нерозчинні сполуки піддаються ферментативному гідроксилюванню, у результаті чого вони стають більш розчинними, легше розщеплюються і виводяться з організму.
Лекція 5.
Тема: «Роль крові у процесі травлення, транспорт живильних речовин в організмі людини.»
Через кров здійснюється взаємозв'язок обміну речовин різних органів тіла. Кров переносить живильні речовини з тонкого кишечнику в печінку й інші органи, а відпрацьовані шлаки транспортує в нирки для наступного виведення їх з організму. Кров доставляє також кисень від легень до тканин і переносить CO2, що утворюється як кінцевий продукт подиху тканин, у легені. Крім того, гормони, що функціонують як хімічні посередники, транспортуються кров'ю від ендокринних залоз до специфічних органів-мішеней. Хімічний склад крові дуже складний, тому що вона містить велике число різноманітних живильних речовин, проміжних продуктів обміну, відпрацьованих шлаків і неорганічних іонів, але саме це і дозволяє їй координувати взаємодію процесів метаболізму в різних органах і поєднувати них у єдину систему.
Об’єм крові в кровоносній системі дорослої людини складає приблизно 5-6 л. Майже половина цього об’єму приходиться на частку еритроцитів (червоних кров'яних кліток), значно менша частина - на частку лейкоцитів (білих кров'яних кліток) і тромбоцитів (рис. 20). Рідка частина крові, плазма, складається на 90% з води і на 10% з розчинених речовин. Понад 70% сухого залишку плазми складають білки плазми (табл. 3). Приблизно 20% приходиться на частку органічних проміжних продуктів обміну (метаболітів), що транспортуються від одних органів до іншим, і на кінцеві продукти обміну - сечовину і сечову кислоту, що, потрапляючи в нирки, виділяються з організму із сечею (табл. 4).
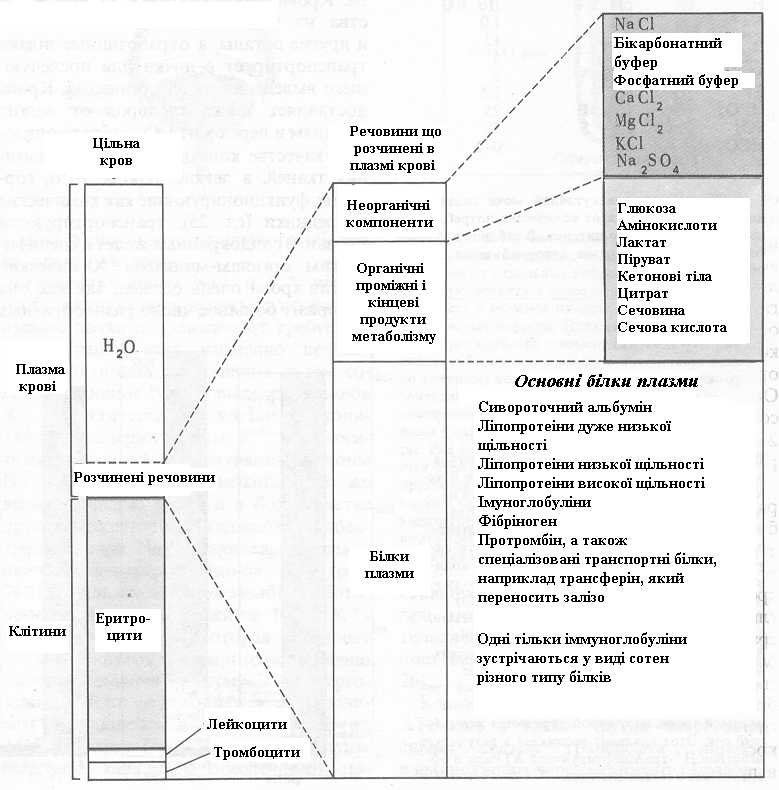 Рисунок
20. – Склад крові. Цільну кров розподіляють
шляхом центрифугування на плазму і
клітини
Рисунок
20. – Склад крові. Цільну кров розподіляють
шляхом центрифугування на плазму і
клітини
Таблиця 3. – Основні фракції білків плазми
Білок |
Кількість мг/100 мл |
Молекулярна маса |
Функція |
Сироватний альбумін |
3500-4500 |
66000 |
Регуляція об’єму крові; транспорт жирних кислот |
α1-Глобуліни |
300-600 |
40000 -60000 |
Транспорт ліпідів тироксину, гормо нів корки наднир-ників |
α2-Глобуліни
|
400-900
|
100000-400000
|
Транспорт ліпідів, міді |
β-Глобуліни |
600-1100 |
110000-120000 |
Транспорт ліпідів, заліза, гемів, імунна активність (антитіла) |
γ-Глобуліни |
700-1500 |
150000-200000 |
Більша частина циркулюючих антитіл |
Фібріноген |
3000 |
340000 |
Попередник фібріну кров’яних згустків |
Протромбін |
100 |
69000 |
Попередник тро-мбіну, необхідно-го для згортання крові |
Загальний вміст білків плазми – 7000-7500 мг/100 мл. В плазмі крові знаходиться велике різноманіття білків; в таблиці наведені лише основні їх класи.
Інші 10% сухого залишку плазми складають неорганічні солі. Ha рис. 20 перелічені основні речовини, розчинені в плазмі крові людини.
За нормальних умов концентрації ряду компонентів у крові можуть коливатися у визначених межах у залежності від характеру харчування, а також від того, у який момент часу узята кров для аналізу. Так, концентрація глюкози в крові досягає максимуму відразу ж після їжі, особливо якщо їжа була багата вуглеводами, а через кілька годин цей показник може бути нижче середнього рівня. Аналогічним чином, концентрація хіломікронів у крові піддається коливанням у проміжках між прийомами їжі.
Вміст окремих компонентів у плазмі крові підтримується на відносно постійному рівні за допомогою різних регуляторних систем.
Таблиця 4. – Концентрації основних небілкових органічних компонентів плазми крові
Компонент |
Межі коливань у нормі, мг/100 мл |
Азотвмісні речовини: |
|
Сечовина |
20-30 |
Амінокислоти |
35-65 |
Сечова кислота |
2-6 |
Креатинін |
1-2 |
Вуглеводи |
|
Глюкоза |
70-90 |
Фруктоза |
6-8 |
Органічні кислоти |
|
Кетонові тіла |
1-4 |
Лактат |
8-17 |
Піруват |
0.4 – 2.5 |
Цитрат |
1.5 – 3.0 |
Ліпіди (повністю зв’язані з білками - α- і β-глобулінами) |
|
Загальна кількість ліпідів |
300-700 |
Триацилгліцероли |
80-240 |
Холестерол і його ефіри |
130-260 |
Фосфоліпіди |
160-300 |
Кров переносить великі об’єми кисню
Нормальний дорослий чоловік у стані повного спокою споживає близько 375 л чистого кисню на добу, що еквівалентне кількості кисню, яка міститься у 1900 л повітря. При сидячій роботі потреба в кисні зростає принаймі вдвічі. У тренованого спортсмена під час змагань по бігу або плаванню швидкість споживання кисню перевищує рівень спокою іноді в 10 разів. B табл. 5 наведені відносні кількості кисню, споживані основними органами тіла дорослої людини в стані спокою і при великому м'язовому навантаженні.
Таблиця 5. - Споживання кисню (у відносних величинах) основними органами тіла людини (дорослого чоловіка)
|
Спокій |
Легка фізична робота |
Важка фізична робота |
Кістякові м'язи |
0.30 |
2.05 |
6.95 |
Органи черевної порожнини |
0.25 |
0.24 |
0.24 |
Серце |
0.11 |
0.23 |
0.40 |
Нирки |
0.07 |
0.06 |
0.07 |
Мозок |
0.20 |
0.20 |
0.20 |
Шкіра |
0.02 |
0.06 |
0.08 |
Інші |
0.05 |
0.06 |
0.06 |
Разом: |
1.00 |
2.90 |
8.00 |
Як ми вже бачили, велика частина кисню переноситься гемоглобіном еритроцитів. Ці клітки утворюють найважливішу «тканину» тіла людини, оскіль-ки їхній загальний об’єм складає 3 л y дорослої людини а загальна вага майже дорівнює ваги печінки. Еритроцити являють собою дуже дрібні дегенеративні клітки, позбавлені ядра, мітохондрій і інших внутрішньоклітинних органел. B їхньому власному обміні кисень не використовується. Відносно невелика кількість необхідного їм ATP виробляється в ході гліколітичного перетворення глюкози, що надходить із крові, у лактат. Основна функція еритроцитів - транспорт O2 з легень у тканини й участь у транспорті CO2 із тканин у легені. 35% маси еритроцита приходиться на гемоглобін, що складає 90% загальної кількості білка, що міститься в клітці.
Гемоглобін - переносник кисню
Гемоглобін складається з двох - і двох β-поліпептидних ланцюгів і чотирьох гемових груп, кожна з яких приєднана до поліпептидного ланцюга. Одна гемова група оборотно зв'язує одну молекулу молекулярного кисню. Оскільки гемоглобін міститься в еритроцитах у великій кількості, у 100 мл цільної крові ссавця при повному насиченні киснем транспортується приблизно 21 мл газоподібного кисню. Зв'язування кисню гемоглобіном залежить від чотирьох факторів: 1) парціального тиску O2; 2) pН; 3) концентрації 2,3-бісфосфогліцерату і 4) концентрації CO2. Ha рис. 21 наведені криві насичення гемоглобіну киснем. Сигмоідна форма цих кривих свідчить про те, що зв'язування першої молекули кисню підвищує здатність решти субодиниць гемоглобіну зв'язувати інші три молекули кисню. При подальшому зростанні парціального тиску кисню криві виходить на плато, що відповідає повному насиченню гемоглобіну киснем, тобто зв’язуванню чотирьох молекул O2 кожною молекулою гемоглобіну. Оборотне зв'язування кисню гемоглобіном супроводжується вивільненням протонів відповідно до рівняння:
HHb+ + O2 ↔ Hb2 + H +
Отже, з ростом pН рівновага повинна зрушуватися вправо і при даному парціальному тиску зв'язування кисню збільшиться. Навпаки, при зниженні pН зв'язування кисню зменшиться.