

Загальна характеристика процесу навчання.
Габітуація і сенситизація – найпростіші форми навчання.
Облігатне і факультативне навчання.
Опосередковане навчання (приклади опосередкованого навчання в роботі Б.П. Мантейфель «Опосредованное обучение» / Додаток 3) та його біологічне значення.
Навички і методи їх вивчення.
Форми навчання, які не пов’язані з підкріпленням.
Дресирування.
Наслідування.
Завдання для самостійної роботи по підготовці до заняття:
Опрацювати літературу за питаннями плану семінару.
Підготувати короткий конспект-відповідь на питання семінару.
Порівняльна характеристика понять «інстинкт», «рефлекс» та «розум» в роботі А.И.Северцова «Эволюция психики» (Додаток 2).
Скласти глоссарій: безумовний рефлекс, умовний рефлекс, імпринтинг, габітуація, навичка, облігатне навчання, факультативне навчання.
Література.
Правоторов Г.В. Зоопсихология для гуманитариев. М.: 2005, - Ст. 36, 40, 45 - 64, 81 -104, 281 – 301.
Фабри К.Э. Основы зоопсихологии. М.: Изд-во Моск. ун-та, 1993. –
Ст. 41 – 87.
Мак-Фарленд Д. Поведение животных. Психобиология, этология и эволюция. М.: Мир, 1988. Ст. 324 – 341.
Дьюсбери Д. Поведение животных. М.: Мир, 1981. Ст.182 -199.
З.А.Зорина, И.И.Полетаева. Элементарное мышление животных. М.: Аспектпресс. 2003 г. – c. 64 – 105.
Теми рефератів
Імпринтинг та його роль в формуванні різних форм поведінки.
Опосередковане навчання та його роль в формуванні поведінки.
Фізіологічні механізми розвитку інстинктивної поведінки.
Основні методи вивчення інстинктів.
Неасоціативне навчання і його роль в формуванні поведінки.
Формування поведінки тварин за Л.В. Крушинським.
Ф. Скіннер та його вклад в науку про поведінку.
Тестові завдання
1.Хто є автором методики «інструментального навчання»?
а) Л.В.Крушинський,
б) Б.Ф.Скіннер,
в) Е.Торндайк,
г) І.П.Павлов.
2. Що таке «фіксований комплекс дій»?
а) цілісний акт поведінки, який формується в результаті інтеграції умовних і безумовних рефлексів, співвідношення яких фіксовано не чітко,
б) зовнішні подразники, які в своїй сукупності складають «пускову ситуацію» для здійснення інстинкту,
в) багато актова поведінка, яка побудована із окремих унітарних реакцій, пов’язана з забезпеченням основних біологічних потреб,
г) видоспецифічні, вроджені, шаблонні рухові акти.
3. Ключовий подразник означає:
а) цілісний акт поведінки, який формується в результаті інтеграції умовних і безумовних рефлексів, співвідношення яких фіксовано не чітко,
б) зовнішні подразники, які в своїй сукупності складають «пускову ситуацію» для здійснення інстинкту,
в) багатокомпонентна поведінка, яка побудована із окремих унітарних реакцій, пов’язана з забезпеченням основних біологічних потреб,
г) видоспецифічні, вроджені, шаблонні рухові акти.
4. Автором «Гідравлічної моделі інстинкту» є:
а) Н.Тінберген,
б) Ч.Дарвін,
в) К.Лоренц,
г) А.А.Ухтомський.
5. Вкажіть, що означає поняття «опосередковане навчання»:
а) індивідуальний досвід, який необхідний для виживання всім представникам даного виду, незалежно від умов життя особини,
б) всі форми індивідуального пристосування до особливостей тих конкретних умов, в яких живе дана особина,
в) миттєва фіксація в пам’яті об’єктів інстинктивних поведінкових актів, яка здійснюється в чітко визначені критичні періоди онтогенезу,
г) навчання методом наслідування.
6. «Комплекс фіксованих дій» представляє:
а) результат інструментального навчання,
б) спадковий руховий стереотип,
в) знаковий стимул,
г) механізм за фіксування.
7. Вкажіть, який з наведених термінів не має відношення до процесу експериментального утворення умовного рефлексу:
а) умовний рефлекс,
б) реакція слідування,
в) гальмування,
г) слиновиділення.
8. Яка з характеристик властива виключно такому феномену, як «імпринтинг»:
а) відбувається на ранніх стадіях онтогенезу,
б) визначає наступний вибір статевого партнера,
в) забезпечує контакт з матір’ю,
г) можливо лише впродовж «чуттєвого періоду».
9. Відмітьте слово, або словосполучення, яке, на Ваш погляд, є «зайвим» в даному ряду:
а) ключовий подразник,
б) етологія,
в) інстинкт,
г) метод проблемних кліток.
10. Вкажіть, яку роль в еволюції відіграють інстинкти:
а) сприяють формуванню в онтогенезі видоспецифічної поведінки,
б) сприяють пристосуванню до епізодичних змін зовнішнього середовища,
в) сприяють пристосуванню до періодичних змін зовнішнього середовища ,
г) сприяють пристосуванню до раптових змін зовнішнього середовища.
11. Відмітьте слово, або словосполучення, яке, на Ваш погляд, є «зайвим» в даному ряду:
а) дзвінок,
б) башта мовчання,
в) П.К.Анохін,
г) собака.
12. Відмітьте слово, або словосполучення, яке, на Ваш погляд, є «зайвим» в даному ряду:
а) умовний рефлекс,
б) метод спроб та помилок,
в) Б.Ф.Скіннер,
г) звикання.
13. Вкажіть, яку роль в еволюції відіграє облігатне навчання:
а) сприяють формуванню в онтогенезі видоспецифічної поведінки,
б) сприяють пристосуванню до епізодичних змін зовнішнього середовища,
в) сприяють пристосуванню до періодичних змін зовнішнього середовища ,
г) сприяють пристосуванню до раптових змін зовнішнього середовища.
14. Вкажіть, яка з форм навчання відноситься до «неасоціативного навчання»:
а) імпринтинг,
б) опосередковане навчання,
в) факультативне навчання,
г) звикання.
15. Відмітьте слово, або словосполучення, яке, на Ваш погляд, є «зайвим» в даному ряду:
а) К.Лоренц,
б) гідравлічна модель,
в) ключовий подразник,
г) ієрархічна теорія інстинкту.
16. Вкажіть, які із компонентів всього комплексу поведінкових реакцій не обумовлені спадково:
а) інстинкти,
б) навчання,
в) елементарна мислительна діяльність,
г) імпринтинг.
17. Встановіть правильну послідовність подій в ланцюгу поведінкових дій тварини:
а) релізер,
б) потреба,
в) завершальний акт,
г) специфічна мотивація,
д) пошукова поведінка,
ж) фіксований комплекс дій,
з) анти драйв.
18. Вкажіть, який із видів навчання відноситься до категорії «неасоціативного»:
а) диференціювальне навчання,
б) латентне навчання,
в) навчання «вибір за зразком»,
г) установка на навчання.
19. Факультативні форми навчання:
а) потребують співпадання в часі (асоціації),
б) залежать від стимулів (подразників), їх інтенсивності та послідовності,
в) не потребують асоціацій в часі,
г) є специфічними для кожної особини.
20. Відмітьте слово, або словосполучення, яке, на Ваш погляд, є «зайвим» в даному ряду:
а) звикання,
б) проблемний ящик,
в) Б.Ф.Скіннер,
г) асоціативне навчання.
21. Факультативне навчання характерно:
а) для раннього етапу онтогенезу,
б) для більш пізніших етапів онтогенезу,
в) для всього онтогенезу,
г) не пов’язано з етапами онтогенезу.
22. Облігатне навчання характерно:
а) для раннього етапу онтогенезу,
б) для більш пізніших етапів онтогенезу,
в) для всього онтогенезу,
г) не пов’язано з етапами онтогенезу.
23. Вкажіть, який із методів навчання може використовуватися для оцінки когнітивних здатностей тварин:
а) навчання в Т-подібному лабіринті,
б) утворення умовних рефлексів слиновиділення,
в) утворення рухових умовних рефлексів,
г) диференціювальне навчання.
24. Вкажіть, хто є автором методики «інструментального навчання»:
а) Л.В.Крушинський,
б) Б.Ф.Скіннер,
в) Е.Торндайк,
г) І.П.Павлов.
Додаток 2 а.Н. Северцов Эволюция и психика
< ... > Существуют два способа приспособления организмов к изменениям окружающих условий: 1. наследственные изменения организации, способ, посредством которого достигаются весьма значительные количественно приспособленные изменения строения и функций животных; способ весьма медленный, посредством которого животные могут приспособиться только к очень медленно протекающим и весьма постепенным изменениям среды; 2. способ не наследственного функционального изменения строения, посредством которого животные могут приспособляться к незначительным, но быстро наступающим изменениям окружающих условии. И в том, и в другом случае строение организмов изменяется. Оба эти способа приспособления существуют и у животных, и у растений. Кроме них существуют еще два способа приспособления, которые встречаются только у животных и которые мы могли бы обозначить как способы приспособления посредством изменения поведения животных без изменения их организации. Они являются для нас особенно интересными и этот вопрос приводит нас к рассмотрению различных типов психической деятельности животных в широком смысле этого слова. Мы знаем три основных типа психической деятельности у животных, а именно рефлекторную деятельность, инстинктивную и деятельность, которую мы условно обозначим как "деятельность разумного типа". Само собой разумеется, что я здесь рассматриваю этот вопрос о психической деятельности животных не как психолог, и соединяя эти три типа (рефлекс, инстинкт и " разумный тип") в одну общую группу, хочу только выразить, что здесь мы имеем деятельности одного порядка. Термином рефлекс мы обозначаем наследственные, однообразные, правильно повторяющиеся целесообразные, т. е. приспособительные реакции организма на специфические раздражения. Обыкновенно говорят, что рефлекторные действия отличаются машинообразностью, определение, которое только до известной степени точно, так как далеко не всегда одна и та же реакция следует за одним и тем же раздражением. Рефлекторная деятельность является наследственной, т. е. молодое животное начинает производить те же рефлекторные действия, которые производили его родители, без всякого предварительного обучения, вполне правильно. Точно так же как и рефлекторная деятельность, инстинктивная деятельность является целесообразной, наследственной и до известной степени машинообразной, но отличается от рефлекторной своей гораздо большей сложностью. Здесь мы обычно находим длинный ряд сложных целесообразных действий, являющихся ответом на определенное внешнее раздражение. "Деятельность разумного порядка" является так же целесообразной, но в отличие от предыдущих типов психической деятельности не наследственной и не машинообразной. Наследственной является способность к деятельности данного типа, но не самые действия, и животные являются наследственно весьма различными в этом отношении: одни способны к сложным действиям "разумного" порядка, другие к весьма элементарным, но самые действия не предопределены наследственно и в индивидуальной жизни не являются готовыми, как рефлексы и инстинкты: для производства определенного действия требуется определенная выучка. Далее эти действия не являются "машинообразными": за определенным раздражением могут следовать весьма разнообразные действия. Сопоставляя эти три типа приспособительной деятельности животных, мы видим вполне ясно, что мы можем распределить их по основному сходству между ними на две группы: к одной будут относиться рефлексы и инстинкты, которые отличаются друг от друга только количественно, к другой - действия "разумного типа": первые наследственны (как действия), не требуют выучки и машинообразны, вторые не наследственны, требуют выучки и в общем не машинообразны. Совершенно ясно, что при сравнении с приспособительными изменениями строения животных, инстинкты и рефлексы будут соответствовать наследственным изменениям строения органов, действия "разумного" типа - функциональным изменениям органов. Рефлексы свойственны всем животным и в общем хорошо известны: на них я не буду останавливаться, и приведу некоторые примеры, поясняющие биологическое значение инстинктов с одной стороны, действий "разумного" типа - с другой. В различных группах животных преимущественное значение имеет либо тот, либо другой тип деятельности. Суживая нашу задачу и принимая в соображение только метамерных билатерально симметричных животных, мы находим, что в типе членистоногие преимущественное значение приобрела деятельность типа инстинкта, в типе хордат - психика "разумного" типа, мы говорим, конечно, только о преимущественном значении, а не об исключительном, так как несомненно и у членистоногих психика "разумного типа" играет известную, хотя и второстепенную роль (мы говорим главным образом о высших представителях этого типа, насекомых, психика которых относительно хорошо изучена и у высших хордат, т.е. позвоночных существуют сложные инстинкты, как например строительные инстинкты птиц и т.д. В типе членистоногих мы видим постепенное повышение инстинктивной деятельности, причем у высших представителей типа, у насекомых, инстинкты сделались необычайно высокими и сложными, и достигли высокой степени совершенства, вследствие чего сознательная психика если не атрофировалась, то во всяком случае отступила на задний план. Напомню необычайно сложные строительные инстинкты общественных насекомых, соты пчел, гнезда муравьев и термитов и т.д. Сложность и постоянство (машинообразность) инстинктов здесь очень ясны, точно так же, как и удивительная целесообразность. В качестве примера возьмем одну из роющих ос, сфекса, и посмотрим, в каких действиях выражается её инстинкт заботы о потомстве. Сначала оса роет норку, сообщающуюся узким коридором ее ячейкой, в которой откладывается добыча, служащая пищей будущей личинке; затем этаа добыча (сверчок) отыскивается и после некоторой борьбы крайне своеобразным способом и делается неподвижной и беспомощной: сфкс перевертывает сверчка на спину, придерживает его лапками и своим жалом колет его три совершенно определенные места, а именно в три передних нервных ганглия брюшной, нервной цепочки: в результате добыча остается живой, но парализованной, так что не может двигаться. После этого сфекс приносит к норке, кладет у входа, влезает в норку, вылезает из нее, втаскивает туда добычу, откладывает в совершенно определенное место eе тела яйцо, из которого впоследствии вылупится личинка и наконец заделывает ячейку; затем в другую ячейку откладывается другая добыча, совершенно так же парализованная и т.д. Целесообразность этого инстинкта поразительна, слабая личинка снабжается свежей пищей, которая не портится в течение всей жизни личинки, и вместе с тем добыча неподвижна в такой степени, что не может сбросить с себя личинку-хищника, питающегося ее телом. Аналогичных примеров того же инстинкта можно привести немало для других насекомых. Весьма интересно, что этот очень сложный вид инстинктивных действий является вполне наследственным: животное производит их без всякой выучки, без всяких изменений из поколения в поколение с замечательной правильностью.
Машинообразность инстинктивных действий, отличающих их от действий, которые нам приходится отнести к типу "разумных " особенно бросается в глаза при так называемых ошибках инстинктов, когда правильный ход процесса вследствие каких либо причин нарушается и окончание его становится вполне бесполезным и бесцельным, и тем не менее животное заканчивает его по раз навсегда установленной рутине. Если у роющей осы, принесшей парализованную добычу, в то время как она осматривает норку, утащить добычу, то она некоторое время ищет ее, но затем успокаивается и заделывает пустую норку совершенно так же, как будто там была положена добыча, что вполне бесцельно. Пчелы имеют обыкновение исправлять поврежденные ячейки сотов, причем производят ряд сложных действий; при нормальных условиях эти действия вполне целесообразны, но пчелы проделывают те же действия, когда ячейка повреждена наблюдателем и пуста и действия совершенно бессмысленны. Таких примеров можно привести много и они показывают, что инстинкты суть приспособления видовые, полезные для вида в такой же степени, как и те или другие морфологические признаки и столь же постоянные. Если мы будем следить за последовательным изменением инстинктов в течение жизни насекомого или паука, то оказывается, что целый ряд инстинктов сменяет друг друга и что каждый инстинкт соответствует организации и образу жизни животного в определенный период жизни особи: при этом каждый инстинкт определенного периода жизни является готовым, действует определенное время и сменяется новым, таким же совершенным, когда изменяется при индивидуальном развитии организация и образ жизни особи, например, когда личинка превращается в куколку и т.д. Мы видим таким образом, что инстинкты суть приспособления, во многих случаях очень сложные и биологически весьма важные, что приспособления эти являются вполне стойкими, т. е. повторяются у каждой особи неизменно из поколения в поколение. Мы выше поставили вопрос о том, к какой из перечисленных нами категорий приспособлений рефлексы и инстинкты относятся? Мы видим, что наш ответ на этот вопрос был правилен и что он вытекает из самого характера инстинктов и рефлексов, а именно из того, что и те и другие наследственны; при этом весьма существенно, что наследственным признаком является не способность к действиям определенного типа, а самые действия с их типичными чертами, т. е. последовательностью определенных движений, их характером и т.д. В виду того, что и инстинкты, и рефлексы являются приспособлениями наследственными, они эволюционируют точно так же, как и прочие наследственные признаки, т. е. крайне медленно и постепенно, посредством суммирования наследственных мутаций инстинктов. Таким образом, эти чрезвычайно важные для организма приспособления суть приспособления к медленно протекающим изменениям внешней среды и о них мы можем сказать то же, что сказали о наследственных морфологических изменениях организма, количественно они могут быть очень велики, но протекают очень медленно и поэтому не могут иметь значения для животных, когда последние подвергаются относительно быстрым неблагоприятным изменениям среды. Иной характер имеют психические свойства организмов, которые мы относим к категории "разумных". Здесь наследственной является только известная высота психики и способность к определенным действиям, но самые действия не предопределены наследственно и могут быть крайне разнообразными. При этом эти сложные действия не являются готовым ответом на определенные внешние раздражения или внутренние состояния организма, как в случаях инстинктов и рефлексов: каждая особь выучивается им заново в зависимости от тех более или менее своеобразных условий, в которых она живет, чем достигается необыкновенная пластичность этих действий, громадная по сравнению с инстинктами. У читателя может возникнуть вопрос о том, существуют ли в действительности у животных действия, которые мы могли бы отнести к категории "разумных"? Во избежание недоразумение предупреждаю, что я употребляю этот термин только с классификационной точки зрения, чтобы отличить известную категорию действий животных от других, а именно, от тех, которые мы охарактеризовали терминами инстинкт и рефлекс. Я здесь совершенно не вдаюсь в вопрос о том, обладают ли животные (и сети обладают, то в какой степени) самосознанием, способны ли животные к абстракции и т.д. Как пример самой низкой степени процессов психических этой категории, мы можем привести так называемые условные рефлексы: животное приучается реагировать постоянно и до известной степени машинообразно на раздражение, на которое оно нормально этим способом совершенно не реагировало: например, у него начинает выделяться слюна, когда оно слышит определенный звук, или при ином раздражении, при котором нормально слюна не выделялась, и таким образом устанавливается новый рефлекс: этот рефлекс отличается от обычного типа рефлексов тем, что он не наследствен и приобретен животным в необычно короткое с эволюционной точки зрения время. В искусственных условиях условные рефлексы могут быть нецелесообразны с биологической точки зрения, но по аналогии мы имеем полное основание думать, что вполне целесообразные условные рефлексы, имеющие приспособительный характер, могут устанавливаться и в естественной обстановке животных и что здесь они имеют весьма большое биологическое значение. Мы знаем, например, что некоторые дикие животные, например, птицы и млекопитающие, живущие на уединенных островах и не знавшие человека, при первом появлении его ведут себя как ручные животные и не боятся человека, не убегают от него и т.д.; при повторном появлении его и после того, как они испытали неудобства и опасности, проистекающие от присутствия этого нового для них существа, они начинают пугаться и убегать: установился новый условный рефлекс. Между очень простыми условными рефлексами и несравненно более сложными действиями, которым животные выучиваются, и в которые несомненно входит элемент разумности, существует полный ряд постепенных переходов. Близка к условным рефлексам и способность диких и домашних животных к дрессировке, т.е. к приобретению новых навыков: мы преимущественно знаем эту способность по домашним животным: и благодаря ей животное может производить крайне сложные и весьма целесообразные (конечно, с человеческой точки зрения) действия. Охотничья собака приучается ложиться и вставать по команде, идет на свисток к хозяину, идет по его команде в определенном направлении, знает классические слова "тубо" и "пиль", подает дичь и т.д.. Многие комнатные собаки, например, пуделя, проделывают гораздо более сложные веши, отворяют и затворяют двери, приносят определенные вещи, снимают с хозяина шляпу, лают по команде, ходят за покупками и аккуратно приносят их. Всякий знает удивительные вещи, которые проделывают дрессированные лошади, свиньи и другие животные в цирке. Весьма интересно, что к дрессировке способны не только домашние, но и дикие и прирученные животные. Как известно, слоны, обезьяны, даже такие крупные хищники, как львы, тигры, медведи, поддаются дрессировке и после выучки у искусного дрессировщика проделывают удивительные штуки, носят поноску, прыгают через кольца, маршируют и т.д. В известных отношениях эти сложные действия близки к простым условным рефлексам, о которых мы только что говорили, но вместе с тем отличаются от них тем, что, во-первых, они несравненно сложнее, во вторых, тем, что в них, несомненно, до известной степени входит тот элемент, который мы у человека относим к категории разума. Конечно, я этим не хочу сказать, что животное понимает мысли и цели человека, который с ним имеет дело, т.е. что собака, которую охотник заставляет идти у ноги, понимает, что он боится, что она спугнет дичь и т.д. Но всякий, кому приходилось дрессировать собаку или лошадь, знает, что одна из главных трудностей дрессировки состоит в том, чтобы добиться, чтобы животное поняло то, что от него требуют. Сказать, что мы имеем здесь дело только с условным рефлексом - едва ли можно. В разбор этого, уже психологического, вопроса я вдаваться не буду, да он для нас и неважен. Для нас интересно, что как у домашних, так и у диких животных при известных условиях (приручении и дрессировке) устанавливаются в сравнительно короткое время и простые, и очень сложные и длинные ряды новых действий, которых животное в обычной обстановке не производит и без этой выучки не способно произвести. По аналогии мы имеем полное право заключить, что высшие позвоночные (птицы и млекопитающие) и в естественной обстановке могут приобретать новые привычки и навыки, вызывающие ряды сложных действий уже биологически целесообразных. Тут может появиться сомнение в том, возможна ли такая выучка (дрессировка) без дрессировщика. Наблюдения над домашними животными, а отчасти и над дикими, устраняет это сомнение: мы знаем, что птицы и млекопитающие сами, без дрессировки, выучиваются новым для них и сложным действиям. Собаки и кошки выучиваются отворять двери, доставать еду из тех мест (шкафов, полок), куда она спрятана и т.д. Молодого шимпанзе (которого исследовала Н.Н. Коте), когда он разыгрался, заманили в клетку с двумя дверцами, одной закрытой, другой открытой и для приманки положили в нее грушу; бегая, он немного приоткрыл закрытую дверцу, потом быстро вбежал в открытую дверь, схватил грушу и выскочил в другую дверцу клетки. Одно время у меня жил попугай, который сам выучился отворять дверцу своей клетки, запертую на задвижку. Подобные примеры показывают, что животные, по крайней мере высшие позвоночные, способны вырабатывать новые и целесообразные способы поведения вполне самостоятельно. Примеры, которые мы приводили до сих пор, касаются прирученных животных; является вопрос - происходит ли тоже и в естественной обстановке, т.е. способны ли дикие животные вырабатывать под влиянием изменений внешней среды новые способы действия и новые привычки приспособительного характера? Аналогия с прирученными животными является сильным аргументом в пользу этого предположения, но и кроме этой аналогии имеются, как мне кажется, указания на то, что такие привычки действительно вырабатываются. Трудность решения этого вопроса заключается в значительной степени в том, что наблюдение животных в их естественной обстановке всегда затруднительно, и что нам приходится в данном случае пользоваться материалом, доставляемым нам путешественниками, коллекционерами, охотниками и т.д., к наблюдениям которых зоопсихологи, особенно современные, склонны относиться крайне скептически. Принимая, что осторожность по отношению к достоверности сообщаемых сведений, конечно, здесь необходима в той же степени, как и по отношению ко всяким другим наблюдениям биологического характера, как, например, относительно времени гнездования, перелета и т.д. я думаю, что мы свободно можем, в виду особенности постановки нашего вопроса, пользоваться этими данными. Дело в том, что зоопсихологи совершенно справедливо относятся скептически к толкованиям, даваемым наблюдателями действий животных, когда, например, согласованные действия общественных насекомых приписываются их взаимной симпатии, когда говорят об особенной сообразительности пчел или муравьев и о разумности постройки гнезд птиц или жилищ бобров и т.д.; но для нас не интересен вопрос о том, что чувствуют те или иные из рассматриваемых нами животных при своих действиях, что они думают, словом, о чисто психологической стороне их деятельности (поэтому мы и употребляем такие неопределенные термины, как действия "типа разумных"): мы ставим вопрос о том, в какой мере я настолько скоро способны высшие животные изменять характер своих приспособительных действий при изменении внешних условий. Трудность проверки относительно диких животных состоит в том, что нам приходится принимать в соображение только те стороны их поведения, которые касаются несомненно новых для них условий существования: таким образом отпадает целый ряд проявлений их психической деятельности, в которых мы могли бы заподозрить существование уже установившихся привычек и инстинктов, например, их поведение при ловле привычной добычи, способы спасания от привычных врагов и т.д. Принимая во внимание это ограничение, мы тем не менее находим ряд примеров, которые показывают нам, что высшие позвоночные приспособляются к несомненно новым для них условиям. Рузвельт в своем путешествии по Африке приводит факт, что слоны изменили свое поведение с тех пор, как за ними стали охотиться охотники с дальнобойными винтовками: они перестали пастись в открытой местности, где к ним охотник может подкрасться издали и использовать свое дальнобойное оружие, а стали держаться в лесу, где их отыскать гораздо труднее и где дальнобойное оружие не представляет преимуществ; охота за ними стала гораздо труднее и истребление приостановилось. Интересно, что носороги, гораздо более тупые, не приобрели этой привычки и поэтому усиленно истребляются. Это изменение у слонов произошло очень быстро, в течение одного поколения, так что о наследственном изменении инстинкта здесь говорить нельзя. Совершенно аналогичное изменение в повадках произошло у бизонов в Канаде: они тоже под влиянием преследования из степных животных сделались лесными, и тоже в короткое время. С рассматриваемой точки зрения весьма характерным является отношение диких животных к различного рода ловушкам; здесь животное сталкивается с совершенно новыми для него опасностями, которые подготовляет ему человек и изменение его поведения, после сравнительно немногих опытов, является весьма показательным. Песцы, которым клали приманку, соединенную шнуром с настороженным ружьем, первоначально ее хватали и погибали, но весьма скоро стали прорывать ход в снегу и схватывать приманку снизу, так что выстрел не попадал в них и они благополучно утаскивали добычу. В качестве аналогичного примера упомяну о так называемых, "контроблавах" на оленей: когда в данной местности произведено несколько облав, то поведение оленей изменяется, и они, вместо того, чтобы бежать от шума, производимого загонщиками, на стоящих тихо и спрятанных охотников, начинают бежать на шум, т. е. на загонщиков и прорываются через их линию и таким образом уходят. Это становится настолько постоянным, что стрелкам приходится становиться позади загонщиков, тогда олени нарываются на них. Некоторые интересные случаи такого изменения поведения были сообщены мне нашим известным орнитологом, проф. П.П. Сушкиным; и ввиду авторитетности наблюдателя я их здесь приведу. Если коллекционер сторожит хищную птицу у гнезда с птенцами (П.Л Сушкин наблюдал это относительно соколов), то старые птицы, заметив охотника, не подлетают к гнезду и держатся от него на почтительном расстоянии; при этом птенцы, сидя без корма, голодают и пищат. В этом случае иногда старые птицы, принося пишу для птенцов, не опускаются с ней в гнездо, а пролетая высоко над ним, т.е. вне выстрела охотника, бросают добычу в гнездо; конечно, она далеко не всегда падает птенцам, но все-таки иногда попадает и съедается. Тут мы видим ряд сложных действий явно приспособительного характера, которые едва ли можем истолковать иначе, как употребляя такие термины как сообразительность, смекалка и т.д. Другой случай тоже весьма характерен: если ворона пытается утащить птенца из выводка домашних уток, то сначала она просто бросается на утят и иногда ей удается схватить утенка и утащить его; но если старая утка отбила нападение и повторные попытки нападения не удаются (старые утки защищают птенцов весьма ожесточенно, и собирают утят под себя), то ворона начинает сильно кричать и обыкновенно на крик прилетает другая ворона и атака возобновляется вдвоем: одна из ворон нападает на утку и дразнит ее, стараясь отвлечь от утят, а другая держится в стороне и пользуется моментом, когда утка занята дракой с се компаньонкой, чтобы схватить утенка и утащить его. По мнению П.П. Сушкина, факт, что первоначально атака производится одной птицей и только в случае неудачи другая призывается на помощь, показывает, что мы имеем здесь не постоянный инстинкт, а ряд индивидуальных действий приспособительного характера. Оценивая теоретическое значение только что приведенных примеров, мы должны обратить внимание на кратковременность того периода времени, в течение которого вырабатывается изменение поведения) животных: здесь мы имеем развитие психической деятельности, совершенно отличной от инстинктивной и, наоборот, весьма похожей: на сообразительность человека, где после нескольких попыток выбирается наиболее целесообразный метод поведения. Если мы сопоставим все сказанное относительно только что рассмотренного нами типа действий высших животных, то мы можем сделать несколько небезынтересных для эволюционной теории выводов. 1. У высших позвоночных животных широко распространены действия, которые в отличие от наследственных рефлексов и инстинктов мы имеем полное право отнести к типу, который мы обозначим условным термином "разумный"; в низшей форме эти действия подходят под тип простых условных рефлексов; у более высоко стоящих животных они усложняются настолько, что приближаются к действиям, которые мы у человека обозначаем, как произвольные и разумные действия. 2. В отличие от инстинктов, эти действия не наследственны и этим отличаются от инстинктов и рефлексов, наследственными признаками являются здесь не самые действия, как таковые, а только некоторая высота психической организации (способности к установке новых ассоциаций и т.д.). С биологической точки зрения, т. е. с точки зрения приспособляемости животных, мы имеем здесь фактор чрезвычайной важности, биологическое значение которого до сих пор не было достаточно оценено: значение его состоит в том, что он в весьма значительной степени повышает пластичность животных по отношению к быстрым изменениям среды. При изменении внешних условий животное отвечает на него не изменением своей организации, а быстрым изменением своего поведения и в очень большом числе случаев может приспособиться к новым условиям весьма скоро. Чтобы оценить значение этого фактора (мы говорим именно о биологическом значении его) нам надо принять в соображение факт, что многие органы высших животных являются органами с полиморфными функциями. Мы знаем, что весьма многие органы животных, имеющие отношение к внешней среде, способны к довольно разнообразным функциям. Это положение касается прежде всего органов движения: мы видим, например, что конечности высших позвоночных ; способны к перемене функции без всякой перемены строения. Крылья крупных птиц и служат для полета, но в случае надобности птица ими пользуется как органами нападения и обороны: орлы наносят крыльями сильные удары и сбрасывают при случае добычу со скал ударом крыла. Задние лапы птиц с служат не только для передвижения по земле (первичная функция задней конечности), но и для обхватывания веток при сидении (на них, для схватывания, перенесения добычи при полете, для нападения и защиты. То же можно сказать и о передних лапах многих млекопитающих, которые служат для бегания, для лазания, для плавания, и в качестве органов обороны и т.д. Даже ноги копытных животных, гораздо более специализированные, чем пятипалые конечности, служат и в качестве органов передвижения, и в качестве органов обороны. Напомню о необычайно разнообразных функциях всех четырех конечностей обезьян и лемуров; о разнообразных функциях рта и зубов очень многих млекопитающих и клюва некоторых птиц, например, попугаев; о необычайном многообразии действий, которые может произвести своим хоботом слон и т.д. Даже кишечный канал способен к довольно разнообразным функциям: мы знаем, что многие животные, нормально питающиеся определенной пищей, как, например, млекопитающие, у которых о роде пищи, которой они питаются, можно судить по строению зубов, переходят в случае нужды к другому типу пиши, например, от мясной к растительной, или от мясной к питанию насекомыми, и свободно переваривают эту пищу. Я не буду останавливаться на подробном перечислении таких примеров органов с полиморфными функциями; всякий читатель, несколько; знакомый с биологией, их легко подыщет сам. Для меня важно только отметить, что у многих высших животных (мы их здесь и имеем главным образом в виду) существует полиморфизм функций экзосоматических, органов: другими словами, данное животное может употреблять один и тот же орган, имеющий отношение к окружающей среде, для нескольких, часто весьма непохожих друг на друга функций. Мы можем спросить себя, от чего зависит тот факт, что животное в известный момент своей видовой жизни вдруг станет употреблять данный орган для функций, для которых его предки этого органа не употребляли? Если мы примем в расчет не только быстроту и легкость изменения поведения животных при наличности того типа психики, который мы обозначили как "разумный", но и полиморфность функций органов, то нам станет понятно все громадное значение этого типа психической деятельности как фактора приспособления. Мы привели ряд примеров более или менее сложных и целесообразных изменений поведения животных при соответственных изменениях условий существования; мы можем представить себе, что изменения эти могут быть еще более значительными, если животные будут при этом пользоваться своими органами для несколько иных целей, чем они ими пользовались раньше: тогда изменения поведения могут привести к весьма значительным изменениям в образе жизни животного. Например, наземное животное может сделаться путем описанного нами активного приспособления из бегающего лазающим или роющим без изменения своей организации, т. е. в весьма короткий промежуток времени. Прибавим к сказанному еще некоторые соображения относительно деятельности "разумного" типа: наблюдая жизнь высших животных, у которых эта психика развита до известной степени высоты, мы видим, что всякое такое животное живет в обычное время среди условий, хотя и в достаточной мере сложных, но в общем повторяющихся; оно имеет дело с определенными условиями неорганической природы, определенным типом растительности данной местности, определенной и в общем знакомой ему фауной, конечно, в той мере поскольку эта фауна, т.е. другие животные, касаются его в качестве конкурентов, врагов, добычи или полезных для него животных. Для того чтобы выжить при этих данных и определенных условиях требуется определенная высота "разумной" психики, и в среднем животные, приспособленные к данным условиям, ею и обладают. Но если условия резко и быстро изменятся в неблагоприятную сторону, т. е. если появится новый и опасный враг (мы берем этот грубый пример для наглядности), то данному виду придется приспособляться к этим новым условиям посредством быстрого изменения определенных сторон своего поведения. Естественно думать, что при этих условиях выживут и приспособятся, т. е. окажутся способными быстро и целесообразно изменить свое с поведение и выработать новые привычки, особи с потенциально более высокой психикой, т.е. животные наиболее умные и наиболее способные: говоря метафорически выживут "изобретатели " новых способов поведения. Другими словами, при эволюции этим путем повышается потенциальная психика, причем дело здесь идет уже о наследственном повышении психических способностей данного типа. Этот процесс, как и другие наследственные процессы, идет, само собой разумеется, очень медленно. Когда животное приспособилось к наступившим изменениям и установилось некоторое новое состояние равновесия, так сказать, некоторая рутина жизни, то эта интенсивная изобретательность, игравшая большую роль в период сильного изменения условий, не требуется, и животное ее может обычно не проявлять, но способность к ней, так сказать, некоторый "запасной ум" в психике животного сохраняется, и при случае, т. е. при наступлении нового изменения условий, может проявиться. Таким образом мы приходим (я отношусь к этому предположению только как к гипотезе) к заключению, что высшие позвоночные животные (птицы и млекопитающие) в общем умнее, чем это кажется при наблюдении их при обычных условиях их жизни. Мне кажется, что опыты дрессировки диких животных (особенно таких, от которых по условиям их существования трудно ждать высокой психики, как, например, тюлени или морские львы) вполне подтверждают эту гипотезу о "запасном уме" млекопитающих. Может быть, эта гипотеза могла бы оказаться полезной при суждении об исключительных проявлениях ума животных, которые мы имеем у лошадей Кралля, собак проф. Циглера и т.д. Вдаваться в разбор этих вопросов, требующих специально психологического разбора (от которого я в настоящей статье сознательно уклоняюсь), я не буду. В предыдущем мы сделали некоторую попытку разобрать способы, посредством которых животные приспособляются к различным изменениям среды, и пришли к выводу, что способов этих два, причем каждый из них может в свою очередь быть подразделен на две категории: первый тип составляют наследственные изменения, которые являются способом, посредством которого животные приспособляются к очень медленным, но вместе с тем и очень значительным изменениям среды. Посредством наследственного изменения изменяются: а) организация животных и вырабатываются те бесчисленные приспособительные изменения, которые нам известны на основании данных палеонтологии и сравнительной морфологии, и б) рефлексы и инстинкты животных, причем изменяется наследственно самое поведение животных; в некоторых случая это изменение поведения происходит без изменения строения органов, в других сопровождал его, т. к. эволюция нового, активного, а частой пассивного органа всегда требует изменения поведения животного. Ко второму типу приспособления относятся ненаследственные приспособления, которые в свою очередь являются приспособлениями к быстрым, хотя и не особенно значительным изменениям в условиях существования животных; сюда относятся: а) те изменения строения, которые мы, за неимением лучшего термина, обозначили как функциональные изменения строения животных и б) изменение поведения животных, происходящее без изменения их строения под влиянием тех психических процессов, которые мы отнесли к разумному типу. Отметим, что в основе и та и других приспособлений лежит, в конце концов, наследственное изменение: способность животных к приспособительным реакциям на раздражения, получаемые из внешней среды, весьма различна у различных животных и мы имеем полное основание думать, что если не сама реакция, то способность к ней наследственна и эволюционирует по типу наследственных изменений. Напомню о различиях в способности к регенерации у различных животных, относительно которых мы с большой вероятностью можем сказать, что они произошли от общих предков. Тоже самое можно сказать о психических действиях разумного типа: самые действия не наследственны, но способность к ним является наследственной и соответственно этому эволюционирует очень медленно. Указанное распределение можно выразить, следовательно, такой классификационной схемой: I. Наследственные приспособления к очень медленным изменениям среды: 1. Наследственные изменения строения животных. 2. Наследственные изменения поведения без изменения строения {рефлексы и инстинкты). II. Ненаследственные приспособления к сравнительно быстрым изменениям среды: 1. Функциональные изменения строения животных. 2 Изменения поведения животных "разумного" типа. Мы видим таким образом, что существует несколько отличных друг от друга способов приспособления животных к окружающей среде, посредством которых они приспособляются к изменениям протекающим с различной скоростью. Эти типы приспособления до известной степени независимы друг от друга, т.е. в одних эволюционных рядах сильнее развиты одни, в других другие. Я не буду подробно разбирать здесь значения наследственных (I, 1) и ненаследственных (ІІ, 1) приспособительных изменений строения животных, и только коротко остановлюсь на эволюции двух остальных типов приспособления посредством изменения действий и поведения животных. Если мы возьмем членистых и членистоногих животных, начиная от аннелид и кончая насекомыми и пауками как высшими представителями, то мы видим, что здесь прогрессивно развивалась деятельность рефлекторно-инстинктивного типа, так что у высших представителей членистоногих, насекомых и пауков, инстинкты достигают высокой степени сложности и совершенства. У многих общественных и одиноких насекомых и очень многих пауков мы должны признать, что психическая деятельность этого типа достигает необычной высоты, сложности и целесообразности: напомню строительные инстинкты пауков, общественные и строительные инстинкты насекомых, инстинкты заботы о потомстве у rex и других и т.д. В каждом из таких инстинктов мы имеем длинный ряд очень точно регулированных и строго повторяющихся действий, которые при обычных условиях существования представляют самые удивительные примеры приспособления животных к совершенно определенным условиям существования. Но даже у тех форм членистоногих, у которых инстинкты достигли высокой степени совершенства, психическая деятельность того типа, который мы обозначили как разумный, стоит относительно весьма низко. Приспособление, посредством перемены способа действий и выучки у них по-видимому играет весьма небольшую роль. Поскольку мы можем судить, эволюция приспособлений при помощи изменения поведения животных здесь пошла в сторону прогрессивного развития наследственно фиксированного поведения (инстинкта). В другом ряду билатерально симметричных животных, а именно у хордат, мы видим что, эволюция пошла в направлении прогрессивного развития психики "разумного типа", т. е. наследственно не фиксированных действий Нельзя сказать, чтобы инстинктов в этом ряду не было, но в общем они гораздо менее сложны и менее распространены, чем у высших членистоногих, и к ним постоянно примешиваются действия "разумного типа"; это мы видим даже в тех случаях, когда мы имеем дело со сложными инстинктами высших позвоночных, как, например, с инстинктом постройки гнезд птиц или заботы о детенышах у амфибий, птиц и млекопитающих. Если же мы возьмем тот тип психической деятельности, который мы обозначим термином "разумный", то в ряду позвоночных, мы в общем видим, что он развивался прогрессивно: у рыб и амфибий он. поскольку мы можем судить, сводится к сравнительно простым условным рефлексам, значительно сложнее он у рептилий, и достигает своего высшего развития у птиц, с одной стороны, у млекопитающих - с другой. И у тех и у других приспособления посредством изменения поведения в течение индивидуальной жизни имеют громадное биологическое значение и позволяют высшим представителям этих двух групп быстро приспособляться к весьма разнообразным условиям и к весьма быстро наступающим изменениям в последних. Последнее особенно ясно видно, когда животным приходится приспособляться к изменениям, вносимым в их жизнь человеком. Наибольшее значение приспособлений этого тепа мы, конечно, видим при эволюции человека, где они несомненно играли первенствующую роль. Можно сказать, что благодаря развитию сознательно-разумной психики, способность непосредственных предков человека и самого человека к приспособлению повысилась в невероятной степени и что именно благодаря этой способности человек и занял, не только в ряду млекопитающих, но и в ряду всех животных, доминирующее положение; он может приспособляться в чрезвычайно короткое с эволюционной точки зрения время решительно ко всяким изменениям и условиям существования. Может быть, было бы интересно сравнить с этой точки зрения способы приспособления животных и человека к изменениям внешних условий: при таком сравнении мы видим, что относительное значение отдельных факторов приспособления, которые мы только что рассмотрели, весьма различно в разных группах животных и у человека. Представим себе, что какое-нибудь млекопитающее животное переселяется из теплого климата в холодный и приспособляется к новым условиям существования. Обычно мы видим, что у него вырабатываются новые приспособления, т. е. что организация его путем наследственного изменения весьма сильно изменяется и что поэтому самый процесс переселения есть процесс весьма медленный: общие покровы животного изменяются таким образом, что они делаются способными защитить животное от холода, соответственно этому происходит целый ряд изменений во внутренних органах, часто изменяется окраска животного и т.д. Аналогичные изменения мы видим, когда млекопитающее из лесного делается степным, когда оно переменяет пищу и т, п. Даже такие незначительные различия, как питание травой и питание ветками деревьев, сопровождаются изменениями строения: мы знаем, например, что у обыкновенного носорога на верхней губе существует пальцевидный придаток для захватывания веток, которого нет у белого носорога, питающегося травой. Всякому известны удивительные приспособления в языке и лапах дятлов, выработавшиеся как приспособления к сравнительно незначительному изменению в образе жизни и способе питания: лазанию по стволам деревьев и добыванию насекомых и их личинок из под коры и из щелей последних. Напомню, что эти примеры относятся и к птицам и к млекопитающим, у которых приспособление посредством изменения поведения играет большую роль. Обращаясь к человеку, мы видим, что соответствующие и даже гораздо большие изменения в образе жизни, переселения в совершенно иной климат, весьма значительные изменения в способе литания и т.д. не отразились на организации человека, и к таким весьма значительным с биологической точки зрения изменениям человек приспособлялся только изменениями своего поведения и своих привычек. Переселяясь в холодный климат, человек не изменяет своей организации, но изменяет свою одежду, свое жилище и т, д. При встрече с новым и опасным врагом он не вырабатывает новых органов нападения и зашиты, рогов, клыков, чешуи и т.д. но изобретает новый способ борьбы, новое оружие. Другими словами, человек, начиная с очень ранней стадии своей эволюции, начинает заменять новые органы новыми орудиями. Там, где животное для приспособления к новым условиям существования вырабатывает новые особенности строения, требующие громадных промежутков для своей эволюции, человек изобретает (при той же организации) новые орудия, которые практически заменяют ему органы, одежду, согревающую его, огонь для варки пиши; каменный топор, увеличивающий силу его удара, копье, позволяющее поражать врага на расстоянии, лук и стрелы, увеличивающие это расстояние и т.д. и т.д. Благодаря членораздельной речи человек приобрел способность быстро передавать новое изобретение или, с нашей точки зрения, новое приспособление, другим людям, чем увеличилась легкость обучения; слово, песня и затем письмо фиксировали всякое новое изобретение, сделали возможным его передачу из поколения в поколение, и облегчили его усовершенствование и т.д. Я не буду разбирать этой стороны эволюции человека в деталях, так как это выходит далеко за пределы моей задачи: сказанного достаточно, чтобы показать, что тот гип деятельности, который мы у животных обозначали как "разумный" и который у человека уже в полной мере заслуживает этого имени, был у человека необычайно важным фактором прогрессивной эволюции. Главное значение этого фактора заключается в том, что он до крайних пределов повысил способность человека к приспособлению, сделав его существом в самой высокой степени пластичным по отношению к изменениям среды. Мы видели, что этот фактор действует в весьма значительной степени и у других позвоночных и может быть корней его приходится искать очень глубоко среди предков позвоночных: высокого развития он достигает только у высших позвоночных и в конце концов у человека. Эволюция "приспособлений посредством изменения поведения без изменения организации" пошла в дивергируюших направлениях по двум главным путям и в двух типах животного царства достигла своего высшего развития. В типе членистоногих прогрессивно эволюционировали наследственные изменения поведения, инстинкты и у высших представителей их, у насекомых, мы находим необыкновенно сложные и совершенные, приспособленные ко всем деталям образа жизни инстинктивные действия. Вся жизнь общественного насекомого введена в строгие рамки, подчинена строго определенной рутине. Каждый повторяющийся случай обыденной жизни муравья или паука служит стимулом, вызывающим к деятельности определенную, в большинстве случаев весьма совершенную, инстинктивную реакцию: все правили поведения наследственны и даны раз навсегда. Но этот сложный и совершенный аппарат инстинктивной деятельности является вместе с тем крайне громоздким: если происходит изменение в условиях среды, то изменение деятельности, посредством которого животное может приспособиться к новым условиям (если оно к ним приспособляется этим путем, а не развитием новых органов), совершается необыкновенно медленно, так что к быстрым изменениям животное этим путем приспособиться не может. Таким образом, мы здесь имеем тип животных очень совершенных, с высоко стоящей психикой, но у которых пластичность организации не превышает пластичности, достигаемой посредством наследственного изменения организации. В типе хордат эволюция пошла по другому пути, инстинктивная деятельность не достигла очень большой высоты (так же как у членистоногих деятельность разумного типа), но зато приспособление посредством индивидуального изменения поведения, деятельность разумного типа стала развиваться прогрессивно и в высокой степени повысила пластичность организмов: над наследственной приспособляемостью появилась целая надстройка индивидуальной приспособляемости поведения. У человека эта надстройка достигла максимальных размеров и благодаря этому человек является существом, приспособляющимся к любым условиям существования, создающим себе, так сказать, искусственную среду, - среду культуры и цивилизации: с биологической точки зрения мы не знаем существа, обладающего большею способностью к приспособлению, а, следовательно большим количеством шансов на выживание в борьбе за существование, чем человек.
Додаток 3
Б. П. Мантейфель опосредованное обучение
Печатается по изданию: Б. П. Мантейфель. Экология поведения животных. М.: 1980. Изд-во: «Наука». Стр. 83-113.
Почти у всех видов млекопитающих и птиц, а также у многих видов рыб имеет место явление, которое мы называем опосредованным обучением: ото взаимообучение животных, приобретение ими при общении новых элементов поведения, повышающих устойчивость, «надежность» популяции в борьбе за существование. Опосредованное обучение обычно происходит на основе врожденной способности животных к подражанию, часто подкрепляется определенной сигнализацией и закрепляется памятью. Поэтому условные рефлексы, приобретаемые в результате опосредованного обучения, физиологи нередко называют имитационными условными рефлексами (Айрапетянц, Герасимов, 1965; Воронин, 1977). Хотя сам факт опосредованного обучения у позвоночных общеизвестен, он подтвержден экспериментальными данными еще весьма недостаточно. В. Я. Кряжев [1928, 1929, 1935, 1955] в экспериментах над млекопитающими четко доказал, что животные исключительно легко образуют условнорефлекторные связи в условиях общения. Он пишет, что условные рефлексы при общении животных вырабатываются в 2 — 3 раза быстрее, чем в одиночестве, что связано с подражанием. Самое главное значение опосредованного обучения заключается в том, что при этом необходимый для жизни животного в тех или иных условиях внешней среды набор приобретаемых элементов поведения создается без непосредственного воздействия на данное животное безусловного раздражителя. Это очень важно при жизни в естественных условиях. Животное получает, например, оборонительные реакции к данному хищнику, не испытав на себе его прямого воздействия, без болевого раздражителя. Ему достаточно лишь видеть, как хищник схватил их соседа по группе, а в ряде случаев наблюдать, как соседи избегают то или иное опасное животное, или просто воспринимать их сигналы опасности. То же можно сказать и в отношении ядовитой пищи: необязательно животным самим испытывать ее ядовитое действие. Вполне достаточно понаблюдать, что едят или чего избегают его более «опытные» соседи. При получении опыта индивидуальным путем каждое животное должно на себе непосредственно испытать вредное действие хищника или ядовитой пищи, что обычно в природе влечет за собой гибель животного. В группе этот опасный безусловный раздражитель заменен примером более опытных особей, а также их сигналами, передающими информацию о надвигающейся опасности. Таким путем необходимые особенности поведения (опыт) передается в общении с себе подобными в естественных условиях с неизмеримо меньшим числом ошибок, а следовательно, и с меньшей потерей численности популяции. В этом, на наш взгляд, заключается прежде всего огромное адаптивное значение опосредованного обучения. Полностью можно согласиться с Л. А. Орбели [1949], который указывал, что имитационное поведение — «это главный охранитель вида», так как «громадное преимущество заключается в том, что «зрители», присутствующие при акте повреждения члена их же стада или их сообщества, вырабатывают рефлекторные защитные акты и таким образом могут в будущем избежать опасности». О механизмах подражания писал Л. Г. Воронин [1957]. Следует заметить, что прекрасный американский натуралист — Э. Сетон Томпсон [1957] пишет, что у каждого дикого животного есть три источника познания. Первый источник — это опыт предков, инстинкт, переданный ему по наследству. Этот опыт накопился у целого ряда поколений за долгие века борьбы с опасностями. Второй источник познания — пример родителей и других взрослых животных того же вида. Детеныш перенимает все обычаи и повадки своего племени. Третий источник познания — собственный опыт. Наследственный инстинкт не всегда помогает животному, так как он недостаточно изменчив и подвижен, а условия жизни постоянно меняются. Пример взрослых тоже не может научить детеныша всему необходимому. А третий источник знания плох тем, что личный опыт всегда приобретается слишком опасным путем. Конечно, написанное относится в основном лишь к млекопитающим и птицам. Можно думать, что Э. Сетон Томпсон приуменьшает при этом значение опосредованного обучения, но в основном с ним следует согласиться. Можно говорить о двух типах опосредованного обучения, постоянно переплетающихся и дополняющих друг друга: обучение в несемейных группах животных и обучение в семейных группах.
Групповое обучение в несемейных группах
Обучение в несемейных группах (кратко — групповое обучение) происходит в стаях и стадах животных, в их временных скоплениях, в их территориальных группах и сообществах. Значение этих групп, и в частности стай, как мы видели выше, весьма многогранно. В данном случае нас интересует «обучающее» значение стаи. Пожалуй, лучше всего оно изучено на рыбах. Еще в опытах Уэлти (Weltty, 1934) было показано, что золотые рыбки (Caras-sius auratus (L.)) научаются проплывать лабиринт значительно быстрее, если они перед этим видели, как это делают другие, уже обученные рыбы. Весьма интересные опыты провели Е. М. Богомолова с сотрудниками [1958] в лаборатории Л. Г. Воронина. Разделив аквариум на две части прозрачной перегородкой, они вырабатывали у рыб (карасей и карпов), помещенных в одном отсеке, условные пищевые двигательные рефлексы (положительный на зеленый свет и дифференцировочный — на красный свет). Условные рефлексы у этих рыб, которых назвали «актерами», проявились закрепились достаточно быстро. Но самое интересное, что у другой группы рыб, названной «зрителями», которые никакого пищевого подкрепления не получали и могли лишь видеть, как его за стеклом получали «актеры», также выработались довольно прочные положительные условные рефлексы на зеленый свет. Эти рефлексы во времени имели волнообразный характер и затухали примерно на девятом-десятом повторений. Представляет интерес и тот момент, что в том случае, если «зрители» видели только выработку условного рефлекса (хватание бусинки) у «актеров», но лишены были возможности видеть акт пищевого подкрепления (он производился в данном случае за непрозрачной перегородкой), то выработать у них «подражательный» рефлекс не удавалось. И. В. Орлов [1958] в лаборатории Э. Ш. Айрапетьянца также отмечает факт стойкости условных имитационных рефлексов, выработанных у карпов. Широкие опыты по образованию имитационных рефлексов у стайных морских рыб в возрасте 1 — 2 года (треска — Gadus morhua morlma L., сайда — Gadus virens L., пикша — Gadus aeglefinus L.) провел В. В. Герасимов [1962, 1964а, 19646, 1965, 1967а, 19676] в аквариуме, разделенном стеклянной перегородкой. Он обучал «рабочую особь» («актера») переходить из одной части аквариума в другую под действием электрического тока. При этом быстро вырабатывалась оборонительная условнорефлекторная реакция ухода от лампочки белого цвета. У «подражающей» рыбы («зритель») также вырабатывался такой же оборонительный условный рефлекс, хотя в ее отсеке электрический ток отсутствовал. Для нее подкреплением являлась двигательно-оборонительная реакция «актера». Следует отметить, что условный рефлекс у «зрителя» вырабатывался значительно быстрее (в 2 — 6 раз), чем у «актера» под непосредственным воздействием электрического тока. Этот имитационный условный рефлекс отличается большой устойчивостью. После двух-трехкратного гашения оказалось достаточным дать два подкрепления (наблюдения за реакцией «актера»), чтобы имитационный рефлекс не только восстановился, но стал бы практически неугасаемым. Он не угасал после 200 — 300 и более предъявлений и даже после перерыва в 1 — 3 месяца. Следует отметить, что в опытах В. В. Герасимова имитационный рефлекс вырабатывался лишь в том случае, если «актеры» и «зрители» не различались ни в видовом отношении, ни размерами. Кроме того, была установлена прямая зависимость между количеством «актеров» и прочностью вырабатываемых имитационных рефлексов у «зрителя». Этот же исследователь установил, что значительную роль в выработке имитационных рефлексов играет передний мозг изучаемых рыб. Попытки выработки двигательно-оборонительных имитационных условных рефлексов у таких типично нестайных рыб, как маслюк (Pholis gannelus L.), бельдюга (Zoarces vivipa-rus (L.) и бычок керчак (Myoxocephalus scorpius L.), оказались безуспешными. Эти рыбы, не имея рефлекса подражания, оказались неспособными к общению и опосредованному обучению (Герасимов, 1964). Следует заметить, что молодь большинства видов рыб ведет стайный образ жизни. Это несомненно связано с тем, что именно в ювенильный период, с одной стороны, происходит наиболее сильная элиминация популяции вида, а с другой — формируются основные характерные черты видового поведения и животные приобретают весь основной набор элементов поведения, необходимый в их дальнейшей жизни. Прежде всего это относится к оборонительному поведению, предохраняющему популяцию от врагов. Именно поэтому Т. С. Лещева в нашей лаборатории обратила особое внимание на опосредованное образование оборонительных рефлексов у молоди целого ряда видов рыб, держащихся стаями [Лещева, 1968, 1971, 1972, 1974, 1975, 1976]. Она проводила эксперименты над стайной молодью таких карповых рыб, как плотва (Rutilus rulilus L.), лещ (Abramis brama (L.)), ry-стера (Blicca bjorkna (L.)), и некоторых других, изучая образование оборонительных реакций в стайках по отношению к различного рода хищникам. В качестве хищников в опытах использовались молодь щуки (Esox lucius L.), ротанов (Percottus glehni Dyb.) и других хищных рыб, а из беспозвоночных — личинки плавунца окаймленного (Dytiscus latissimus). Опыты проводились и по методу «артистов и зрителей» (через стеклянную перегородку) и при непосредственном контакте хищников с подопытной молодью. Последняя методика была наиболее эффективна, поскольку в этом случае «жертвы», находящиеся в стае, имели возможность воспринимать хищника и сигналы схваченных им «жертв» всем комплексом своих рецепторов (включая хеморецепцию), а не только зрением, как в первом случае. В своих экспериментах Лещева прежде всего доказала, что при опосредованном обучении большое значение имеет возраст подопытной молоди рыб. Так, например, у молоди плотвы личинки и ранние мальки (размером от 6 до 13 мм), не образующие стаи, не обучаются или очень слабо обучаются реакциям на хищника. Молодь же (мальки), начинающая образовывать стайку оборонительного типа, хорошо вырабатывает активные оборонительные рефлексы и достаточно способна к опосредованному обучению. Мальки этого возраста, «обученные» на хищника, выедаются им примерно в 2 раза менее интенсивно, чем «необученные». Часто хищник, помещенный в один сосуд со стайкой «обученной» молоди, после нескольких неудачных бросков вообще прекращает свою охоту. Такая стайка обычно держится в противоположной от хищника половине аквариума, не подпуская его на дистанцию его охотничьего броска и выполняя оборонительные маневры при нападении. Можно привести пример из опытов Лещевой [1974] (рис. 1 — 2) по схватыванию хищников (мальком щуки) «обученной» и «необученной» молоди плотвы и верховки. Как видно, за первые 5 мин хищник схватил 20% «обученных» и 40% «необученных», а за 15 мин — 80% «необученных» и около 30% «обученных» мальков. При этом выяснилось, что, чем моложе испытуемая молодь, тем быстрее у нее затормаживается оборонительный рефлекс. Так, например, у молоди плотвы длиной 20 — 25 мм затормаживание началось уже на второй день после обучения, а у молоди длиной 30 — 35 мм — лишь на третий день. При этом Лещева делает заключение, что затормаживание оборонительного рефлекса не снижает его значения для популяции рыб в природе. Угасая у какой-то части рыб в стае, он восстанавливается при столкновении с хищником у другой части. При этом, по всем данным, растормаживание происходит значительно быстрее. Таким образом, у стаи рыб в целом имеется «условнорефлекторный фонд», который в результате подражания является достоянием всей стаи [Радаков, 1970]. Лещева подтвердила это положение рядом опытов. Так, в одном из них она подсадила к 3 малькам плотвы, «обученным» на хищника (личинки жука-плавунца), двух необученных, причем эти двое во всем подражали первым трем и все пять оказались недоступными для хищника, который после безуспешных бросков прекратил охоту. В отличие от стайной молоди рыб опыты Лещевой с головастиками (Rana ridibunda и R. esculenta) показали, что эти животные, держащиеся большими скоплениями, но не имеющие стайности и стайного поведения, не показывали обучения (ни стайного, ни индивидуального) на хищников (ротан и личинка плавунца). Также нестайные мальки гуппи (Lebistes reticulatus (Peters)) длиной 10 мм, выпущенные к хищнику — хромису (Не-nuchromis bimaculatus Gill.), спасались от него индивидуально и соответственно истреблялись значительно интенсивнее. Наши опыты с более взрослой молодью гуппи размерами 15 — 20 мм, посаженными в количестве 10 экз. в аквариум, где содержался взрослый жук-плавунец (в 1957 г.), показали, что гуппи быстро начали держаться оборонительной стайкой. При этом за первые 20 суток жуку удалось съесть только двух гуппи, а остальные 8 стали для него недосягаемыми. За последующие 3 месяца он не смог съесть больше ни одной рыбки, так же как и подсаженных туда дополнительно еще 5 «необученных» гуппи. Более того, гуппи собирались около плавунца, когда его кормили, и выхватывали у него падающие кусочки пищи: червей и мучных червей, но держались очень осторожно и уплывали при каждой попытке нападения. Исходя из всех этих и других экспериментов, мы [Мантейфель, Радаков, 1960; Лещева, 1974] сделали определенный вывод, что количество схваченных из стаи рыб следует рассматривать как «плату» стаи за обучение и что при какой-то определенной оптимальной численности стаи эта «плата» должна быть наименьшей.
 Рис.
1. Результаты опытов по схватыванию
хищником (мальком щуки) обученной (2) и
необученной (1) молоди плотвы (длина 20 —
20 мм) (а) и верховки (длина 30 — 40 мм) (б)
[по Лещевой, 1974]
Рис.
1. Результаты опытов по схватыванию
хищником (мальком щуки) обученной (2) и
необученной (1) молоди плотвы (длина 20 —
20 мм) (а) и верховки (длина 30 — 40 мм) (б)
[по Лещевой, 1974]
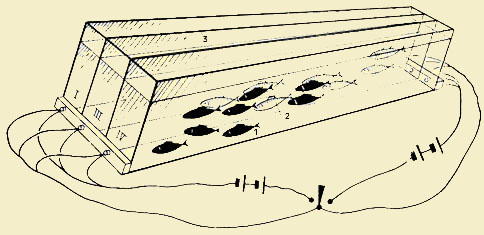 Рис.
2. Схема опыта по опосредованному обучению
молоди методом «актеров и зрителей» в
аквариуме с тремя отсеками [по Лещевой,
1976]
I, II, III — отсеки экспериментального
аквариума; 1 — «актеры» в первом отсеке;
2 — «зрители» во втором отсеке; 3 —
непрозрачная перегородка между вторым
и третьим отсеками
Рис.
2. Схема опыта по опосредованному обучению
молоди методом «актеров и зрителей» в
аквариуме с тремя отсеками [по Лещевой,
1976]
I, II, III — отсеки экспериментального
аквариума; 1 — «актеры» в первом отсеке;
2 — «зрители» во втором отсеке; 3 —
непрозрачная перегородка между вторым
и третьим отсеками
Т. С. Лещева произвела также эксперименты по методике «артисты и зрители», вырабатывая у молоди рыб как пищевые, так и оборонительные имитационные рефлексы. Наиболее интересные из них [Лещева, 1976] были проведены со стайными ак-вариальными рыбами тетрагоноптерусами (Hemigrammus сап-dovittatus Ahl) в аквариуме, разделенном на три отсека (см. рис. 2). В каждый отсек было посажено по 10 рыб. В первом отсеке пищевой рефлекс вырабатывался обычно: сигнальное зажигание лампочки и пищевое подкрепление в заданном месте. Во втором отсеке, отделенном от первого прозрачной перегородкой, «зрители» вырабатывали при этом имитационный пищевой рефлекс, а в третьем отсеке, отделенном сначала от второго непрозрачной перегородкой, затем эту перегородку устраняли и оставляли прозрачную. Таким образом, рыбы второго отсека, выработавшие имитационный условный рефлекс, оказывались для рыб третьего отсека уже в роли «артистов», и у рыб-«зрителей» в третьем отсеке в данном случае выработался имитационный рефлекс второго порядка. Когда их изолировали, они дважды подошли к месту «кормления», а затем при 3-м и 4-м повторении рефлекс угас. Он оказался очень нестойким, поскольку эти рыбы не видели, как происходит подкрепление. У других видов рыб создать имитационный рефлекс в третьем отсеке вообще не удавалось. В августе 1968 г. в старице Москвы-реки плавала стая (шт. 30) головлей длиной 16 — 17 см. Они были очень активны и явно голодны. На удочку, наживленную кузнечиком, бросилась сразу вся стая. Один, схвативший наживку, был вытащен. На брошенную вторично наживку они бросились не сразу, а минут через 5. Третий взял наживку через 15 мин, а затем на протяжении часа ее больше не схватил ни один. Все подходили к наживке и испуганно уплывали прочь. Следовательно, у них оборонительный рефлекс на удочку создался лишь при виде вытаскиваемых из воды сочленов стаи. В связи с этим мы совместно с Д. С. Николаевым на биостанции Кропотово близ р. Оки провели эксперименты над помещенными в большие бассейны стаями пресноводных рыб (голавли, лещи, ерши и некоторые др.). При вылове этих рыб на экспериментальную удочку условный оборонительный рефлекс на нее создавался у стаи в целом примерно после одного — трех наблюдений за тем, как вылавливались особи из их стаи. Это наблюдение за выловом сочленов стаи являлось тем безусловным раздражителем, который сохранялся от одних до нескольких суток, но угаснув, восстанавливался при первом же сочетании, т. е. при наблюдении за выловом. Все эти эксперименты достаточно четко доказывают, что у рыб, находящихся в стае, хорошо образуются как пищевые, так и оборонительные условные рефлексы, что свидетельствует об их способностях к групповому опосредованному обучению. Можно думать, что в стаях птиц и млекопитающих происходит такое же групповое опосредованное обучение. А. И. Ильенко [1965] описывает, как молодые воробьи, вливаясь в стаи несколько более старших воробьев (предыдущего выводка), быстрее «приспосабливаются к условиям существования». В. К. Федоров [1963] отмечает, что подражание в форме «актера» и «зрителя» широко используется разными видами млекопитающих при образовании новых условных связей. В частности, он наблюдал это у лабораторных мышей, причем «зрители» образовывали условный рефлекс на индифферентный раздражитель, подкрепляемый актом еды «актера». Об обучении в стадах копытных много фактов приведено Л. М. Баскиным [1970, 1976], который устанавливает, например, что «формирование поведения оленей, находящихся в стаде, происходит в значительной мере за счет подражания» [1970, с. 9]. В отношении одомашненных животных об этом же пишет Н. М. Носков [1973], который, например, отмечает, что «если теленка, умеющего поедать траву, сено и другие корма, поместить в группу сверстников, не способных это делать, то они, подражая своему „ментору", через несколько дней приобретают те же навыки». И далее: «При появлении в стаде, отаре, табуне одного-двух животных, обладающих способностью преодолевать изгороди, рвы, водные преграды, большинство других в короткий срок научились приемам своих менторов». А. Г. Томилин [1974] отмечает, что у дельфинов превосходно развит рефлекс подражания. Если один дельфин в стайке научился, к примеру, пригибать сеть и таким образом выходить из окружения, этот опыт тут же перенимают другие. Такое подражание значительно облегчает дрессировку дельфинов и сокращает сроки их обучения. Те дельфины, которые долго находились с дрессируемыми сородичами, могли выполнять это действие даже без специальной тренировки. В Калифорнийском океанариуме малая косатка обучилась трюкам только на основе наблюдения за своими компаньонами — афалинами, с которыми работали тренеры. Исходя из изложенного можно считать вполне справедливым замечание И. А. Шилова [1973] о том, что в группе животных открывается возможность использования опыта немногих особей всей группой.
Опосредованное обучение в семейных группах
Это интереснейшее явление — обучение молодых животных их родителями — наиболее сильно развито у птиц и млекопитающих. На него обратил серьезное внимание М. Е. Лобашев [1961, 1963], назвав его термином «сигнальная наследственность». Это явление широко известно, но еще весьма недостаточно исследовано. Оно происходит в результате так называемого биологического контакта поколений [Промптов, 1929] и представляет собой чисто функциональную преемственность приспособительных реакций. При этом предшествующие поколения путем научения передают последующим поколениям накопленную ими информацию и соответствующие особенности поведения. Сами эти особенности не врожденны, не закреплены генетически, но настойчиво передаются потомству в силу наследственного рефлекса подражания родителям или при помощи специальной сигнализации. Происходит как бы передача «по наследству» ненаследственной информации, не входящей в генетическую структуру организма, но через подражание и память настойчиво передающейся из поколения в поколение и в то же время постоянно изменяющейся адекватно изменениям факторов среды с каждым последующим поколением. Можно согласиться с Б. Л. Астауровым [1971], что термин Лобашева «сигнальная наследственность» не совсем удачен. С. Н. Давиденков [1947], говоря о наследственности человека, предлагал наследственностью называть то, что передается из поколения в поколение через половые продукты, а то, что передается посредством обучения, называть социальной преемственностью. Поскольку термин М. Е. Лобашева был предложен для животных, обучение в их семейных группах мы будем называть сигнальной преемственностью. Способность к сигнальной преемственности, широко развитая у высших позвоночных и связанная, конечно, с прогрессивным развитием их нервной системы, чрезвычайно обогатила поведенческие, адаптивные особенности этих животных. Сигнальная преемственность стала здесь как бы дополнительным звеном между врожденными элементами поведения, относительно стабильными, и индивидуально приобретаемыми элементами, чрезвычайно лабильными. Она значительно обогатила и усовершенствовала поведенческий комплекс животных, объединив опыт многих поколений и способствуя образованию у них разнообразной и сложной сигнализации. Мы полагаем, что в начале сигнальной преемственности стоит явление запечатлевания (imprinting), установленное для многих видов позвоночных животных (главным образом для птиц) [Heinroth, 1911; Lorenz, 1935; Hess, 1959, 1972; Schutz, 1965; и др.]. Как пишут Кальтенхаузер и Крушинский [1969], у животных многих видов в онтогенезе имеются определенные «чувствительные периоды», во время которых они научаются реагировать на определенный, строго специфический комплекс раздражителей. При ознакомлении с основной литературой по запечатлеванию можно видеть, что новорожденное животное, когда у него начинают действовать рецепторные системы, запоминает (запечатлевает) в короткий срок и нередко на длительный период окружающую его обстановку и прежде всего своих родителей — их внешний вид, голос, а у млекопитающих — и запах. Кстати, Конрад Лоренц [Lorenz, 1935] ввел термин запечатлевания для обозначедия уникального процесса обучения, когда на ранних стадиях развития выводковые птицы образуют привязанность к внешнему виду матери. Именно запечатлевание родителей и стремление на какой-то срок им подчиняться и подражать создают прочную силу закона для сигнальной преемственности. С физиологической точки зрения запечатлевание подробно разобрано А. К. Понугаевой [1973]. Многие этологи проводили эксперименты, показывая новорожденным птенцам (главным образом утятам) любой движущийся предмет соответствующих размеров (чучело, футбольный мяч, утку другого вида и т. д.), и при этом добивались запечатлевания птенцами этого предмета. В сочетании с рефлексом следования эти птенцы всюду следовали за запечатленным предметом. К. Лоренц произвел запечатлевание на себя выводка инкубаторных гусят, которые потом всюду следовали за ним и сопровождали его в воде при купании. Поскольку запечатлевание запоминается молодым животным весьма надолго, нередко на всю жизнь, при указанных выше экспериментах создавались поведенческие аномалии. Животные считали запечатленный предмет (или человека) не только своей матерью, но и сочленом популяции своего вида. В природе же, как правило, в момент рождения или вылупления около новорожденного оказывается мать или оба родителя. Именно их и запечатлевает (импринтирует) молодое животное в первые часы и даже минуты своей жизни (так называемый чувствительный период). Таким образом, период «запечатлевания» представляется нам очень важным этапом — постнатальным началом сигнальной преемственности у высших позвоночных. Дальше следует целая система воспитания (обучения) этих молодых животных, включающая подражание, следование, целый ряд сигналов, а нередко поощрения и наказания. У некоторых позвоночных животных этот период обучения длится недолго, а у других весьма длительное время. У представителей класса рыб сигнальная преемственность, как правило, отсутствует, хотя, как было показано выше, обучение в стаях («групповое обучение») происходит здесь очень широко. Следует отметить, что у тех немногих видов рыб, для которых установлено семейное поведение (и прежде всего семейства Cichlidae), можно наблюдать элементы сигнальной преемственности, которые несомненно возникли в этом классе животных конвергентно под влиянием специфики взаимоотношений в тропических районах. Для выяснения этого вопроса в нашей лаборатории были проведены эксперименты над семейными группами пятнисто-голубой акары (Aequidens latifrons). А. Д. Мочек [1972 а, 1974, 1976], проводивший эти исследования, пришел к заключению, что мальки акары, держась в семейной группе, лишь на 15 — 17-й день свободного плавания научаются узнавать родителей. До этого они, будучи выпущенными вблизи других взрослых акар, не находящихся в фазе заботы о потомстве, плывут к ним так же, как к родителям. Даже если эти взрослые особи начинают поедать мальков, последние продолжают к ним стремиться, пока не будет уничтожена вся стайка. В специально поставленном эксперименте Мочек выработал оборонительный условный рефлекс у пары акар, готовых к размножению. Этот рефлекс заключался в том, что при зажигании лампочки с одной стороны аквариума рыбы уходили в другую половину аквариума через отверстие в перегородке. После нереста и выклева молоди у родителей этот рефлекс сохранился и автор производил «тренировку» по 20 раз в день. Молодь в силу реакции следования за родителями увлекалась за ними при зажигании лампочки. В результате у мальков выработался аналогичный условный рефлекс, который достигал наибольшей выраженности на 10 — 12-й день после перехода мальков к свободному плаванию: мальки в отсутствие родителей (их отсаживали) убегали от зажигаемой лампочки в другую половину аквариума. В дальнейшем сила рефлекса у мальков несколько угасает, но не исчезает окончательно еще дли тельное время. Спустя 40 — 50 дней после перехода мальков к свободному плаванию, когда семейная группа уже распадается, рефлекс, почти угаснув, проявляется у молоди вновь и с большой силой. Из сказанного А. Д. Мочек [1974] делает вывод, что в классе рыб наблюдаются лишь зачатки «сигнальной преемственности». В классах амфибий и рептилий, где, как указывалось выше, почти отсутствует активная забота о потомстве и соответственно нет семейных групп, не отмечена сигнальная преемственность. Можно лишь предполагать ее элементы у некоторых крокодилов. У представителей класса птиц сигнальная преемственность развита очень сильно. Известно, что почти все виды птиц — и птенцовые и выводковые (кроме так называемых «гнездовых паразитов») воспитывают своих птенцов, обучают их. Это обучение охватывает широкие области жизни: защита от врагов, питание и добывание пищи, полет, ориентация, многие сигналы, особенности пения и т.п. К. Лоренц [1970] описывает особенности обучения птенцов у галок и заключает: «Животное, не осведомленное от рождения инстинктом о своих врагах, получает от более старых и опытных особей своего вида информацию о том, кого и чего следует бояться. Это поистине традиция, передача индивидуального опыта, приобретенных знаний от поколения к поколению». А. Н. Промптов [1956] прекрасно описывает обучение родителями птенцов из отряда воробьиных и приходит к выводу, что «из поколения в поколение передается довольно сложный «арсенал» навыков, составляющих биологические «традиции вида», не являющиеся наследственными, но в большей своей части представляющие как раз самые тонкие «уравновешивания» организма с условиями среды». У выводковых птиц птенцы с первого же дня жизни всюду следуют за матерью, подражая ей, копируя ее движения и подчиняясь ее сигналам. Таким образом, они быстро научаются объектам и способам питания, а также распознаванию своих врагов и способам защиты (затаивания) при тревожных сигналах самки. У птенцовых птиц из трех стадий ювенильной жизни, выделенных Промптовым [1956], видимо можно составить два периода сигнальной преемственности. Первый — начальный период — от вылупления из яйца до покидания гнезда. Это период запе-чатлевания родителей и обстановки. Второй — активный период, когда птенцы оперены, выходят из гнезда, обучаются летать и следуют за родителями, подчиняясь их сигналам. Именно в этот активный период, как отмечает А Н. Промптов, у птенцов образуется огромное количество условных рефлексов и формируются основные черты поведения взрослой птицы. При этом родители, конечно, бессознательно действуют часто по определенным программам. По наблюдениям Т. Л. Бородулиной (1972), выводок поганок, покинув гнездо, чередует плавание и ныряние в воде с обогревом на спине родителей, это делается по программе, определяемой родителями. Птица сбрасывает с себя птенцов в воду и регулирует время их плавания, не давая им вернуться к себе на спину. По мере роста птенцов время их пребывания в воде птица увеличивает. По нашим наблюдениям, самец большой синицы (Parus major) обучал своих летных птенцов маневрировать следующим образом он брал в экспериментальной кормушке кусок пищи и, подлетев к сидящим на ветке птенцам, садился около, а затем летел, маневрируя между ветвями, за ним летела вся стайка птенцов Самец садился и давал кусок первому подлетевшему птенцу. Это повторялось многократно. Самка большого пестрого дятла (Denrocopos major), взяв из той же кормушки кусок хлеба, летела в сопровождении птенца к своей «кузнице», вставляла туда кусок и отлетала в сторону, как бы приучая птенца пользоваться «кузницей». Таких примеров можно привести большое количество. В случае опасности на птенцов действуют сигналы опасности родителей. К. Н. Благосклонов [1976] отмечает, что птенцы мухоловки-пеструшки не боялись ос и пчел в клетке и пытались их клевать. Если же жалящих насекомых впускали в клетку, где, кроме молодых, была и взрослая птица — кормилица, то все птицы, явно испуганные, сбивались в один угол, подальше от опасного насекомого. Очевидно, пишет он, птенцы получали информацию об опасности от взрослой птицы. Многие черты в поведении птиц, которые входят в «видовой стереотип их поведения» [Промптов, 1940] и считались ранее генетически закрепленными, после соответствующих исследований оказываются приобретенными или формирующимися в онтогенезе на базе опосредованного обучения и прежде всего «сигнальной преемственности». Это было установлено у ряда видов птиц для особенностей песни и некоторых акустических сигналов, имеющих в природе определенный видовой стереотип. На этот вопрос было направлено внимание исследователей, которые, казалось бы, получали противоречивые результаты. Так, Ф. Зауер [Saner, 1954, 1955], выращивая птенцов славок в полной звуковой изоляции, пришел к выводу, что их голосовые реакции являются врожденными и для формирования песни им не нужны даже упражнения. Однако наблюдения А. Н. Промптова [1944] и Промптова и Лукиной [1945] показали, что у таких воробьиных птиц, которые отличаются упрощенной песней (зеленушки — Сhо-ris chloris L., овсянки обыкновенные — Emberiza citrinella L., лесные коньки — Anthus triwialis, пеночки-трещотки — Phyllos-copus sibilator Bechst. и пеночки-теньковки — P. collybitus), нормальное формирование песни происходит без влияния со стороны «учителя». Их песни слагаются, несмотря на то, что они никогда не слышали песни самцов своего вида. Однако у большинства видов птиц, обладающих более сложной песней, нормальная песня не может сложиться без имитации песни взрослых самцов своего вида. Для формирования нормальной песни необходимо, чтобы птенец с первых дней вылупления имел возможность слышать поющего неподалеку самца. У выкормышей, выросших в изоляции от птиц, формируется абортивное пение, иногда весьма отличающееся от песни особей своего вида. В отсутствие поблизости поющих самцов ювенильное щебетание сохраняется долго — до трех лет. Промптов [1944], Промптов и Лукина [1945], а также В. Торп [Thorpe, 1954, 1958] делают вывод, что у воробьиных птиц врожденной является лишь основной характер песни, а ее детальные вариации определяются путем имитации поющих особей данного вида. Имитационные способности многих птиц, не говоря уже о таких имитаторах, как попугаи, скворцы и многие другие, широко известны. Любителями, содержащими птиц в клетках, было показано, что пению канарейки обучаются от отца. Лоренц в своей статье, переведенной на русский язык [1969], пишет: «Мой сотрудник И. Николаи обнаружил, что молодые самцы-снегири учатся своей индивидуальной песне только у отца. После того как птенцы оперяются, у них на сравнительно долгий период устанавливаются своеобразные, имеющие явно сексуальный оттенок, отношения с отцом. Молодые снегири припадают перед ним к земле, принимают покорные позы, которые (как и у многих обезьян) напоминают позу самки при спаривании, и вообще они гораздо больше привязаны к отцу, чем к матери. В этот период они и обучаются отцовской индивидуальной песне. У И. Николаи был самец снегирь, выращенный канарейками, и пел он точно так же, как они. Эта «ария кенара» передавалась потомству в чистом виде, без примеси в течение четырех поколений, причем птенцы воспитывались в птичнике, в котором жила масса нормально поющих снегирей. Эксперименты Николая показали также, что молодой снегирь может выучить отцовскую песню только тогда, когда он принимает по отношению к отцу описанную выше позу». К. А. Вилке и Е. К. Вилке [1958], а также Е. К. Вилке [1958, 1959, 1961] провели огромную и необычайно интересную работу по массовому пересаживанию яиц и птенцов одних видов воробьиных птиц в гнезда других видов в природных условиях. Часть выросших окольцованных птенцов в дальнейшем контролировалась на протяжении нескольких лет. Конечно, не у всех этих пересаженных птенцов удалось проследить условия формирования и особенности песни, но весьма интересно, что в ряде случаев из птенцов — самцов в дальнейшем получались как бы «поведенческие гибриды», морфологически они обладали всеми признаками своих основных родителей, а их песни соответствовали песням приемных родителей. Так, одни мухоловки-пеструшки пели, как горихвостки, другие — как большие синицы, а третьи — как пеночки-трещотки. Хотя в природе эти птенцы как в гнездовом, так и в послегнездовом периодах имели возможность слышать песни многих птиц (в том числе и птиц своего вида), но имитировали они, как правило, только тех птиц, которые их кормили, иначе говоря — тех птиц, которых они запечатлевали. Отсюда Е. К. Вилке делает вполне обоснованный вывод, что в формировании песни исследованных певчих птиц решающей оказывается имитация. Этот процесс имитации происходит (по Вилке) главным образом после вылета молодой птицы из гнезда, т. е. в то время, которое мы называем активным периодом сигнальной преемственности. Сложившаяся же в первом году песня в последующие годы не меняется [Вилке, 1961]. Нам тоже неоднократно приходилось наблюдать, как в середине лета старые самцы зябликов летали в сопровождении одного или двух птенцов, которых они кормили. При этом самцы временами пели, а молодые их внимательно слушали. В отношении сигнальных криков (позывов) птиц большинство исследователей считает их полностью врожденными. Однако Е. К. Вилке в своих экспериментах показала, что при производимых пересадках птицы часто перенимали не только песни, но и позывы приемных родителей и использовали их в «правильном смысле» — соответственно ситуации, причем в основном такими имитаторами оказывались самцы. Самки же были менее способны к имитациям и в большинстве случаев пересадок сохраняли крик тревоги своего вида. Из этого Вилке заключает, что сигнальные позывы имеют более определенную врожденную основу, но и они тоже могут имитироваться. Так, она наблюдала самца мухоловки-пеструшки, который вырос в гнезде горихвостки и полностью имитировал ее песню и крик тревоги. Сын этого самца тоже перенял от отца песню горихвостки; крик же тревоги в норме у него был обычный для мухоловки, но изредка он издавал и крик тревоги горихвостки. Даже стереотип гнездования Вилке в своих опытах в какой-то степени постепенно изменяла, подтверждая тем самым вывод Промптова [1938] о том, что довольно устойчивый шаблон выбора места для гнездостроения не является для вида единственно возможным. Он определяется главным образом «традициями» вида — условнорефлекторно. Из сказанного можно сделать предположение, что местные напевы птиц разных регионов представляют собой результат обучения (сигнальная преемственность) и создание местных акустических семейных линий. Этим, вероятно, можно объяснить постепенное исчезновение особых виртуозов-певунов — курских соловьев, которые особенно высоко ценились любителями и вылавливались по показателям их пения. В результате резко ухудшились (с точки зрения любителей соловьев) местные виртуозные напевы курских соловьев. Молодым соловьям не у кого стало учиться. Именно так объясняет данное явление Н. А. Гладков [1970]. Сигнальная преемственность у млекопитающих развита в не меньшей степени. Она, так же как и у птиц, начинается с запечатлевания и реакций следования. Обучение родителями детенышей описано для многих видов. Это — «школы» выдр, где молодые обучаются плавать, ловить рыбу, защищаться; это — обучение в семьях волков, которое длится более года. Особенно интересно, пожалуй, обучение молодых в семьях бурых медведей. Известно, что в восточной части Советского Союза и в Северной Америке медведи занимаются «рыболовством», вылавливая идущих на нерест лососей. В Америке был снят прекрасный фильм, в котором было хорошо видно, как медведи ловят рыбу и как этому обучаются молодые особи. А. Севастьянов [1970] пишет, что камчатские бурые медведи ловят лососевых, подстерегая их с берега и в воде. Они выгоняют лососей на отмель и прижимают здесь. Л. Пашковский и Ф. Картавых [1975] также описывают, как медведи ловят лососевых на Сахалине. При этом они отмечают, что каждый вид лососей (горбуша, сима и кета) имеет собственные особенности поведения. Соответственно у медведей выработались свои навыки для охоты за каждым видом лососей. Сима — стремительная, осторожная рыба, которая днем отстаивается в ямах и идет по реке только в сумерках. Поэтому медведь днем залезает в яму и плавает там, зорко следя за перекатом. Стоит испуганной симе ринуться на перекат, как медведь стремительно бросается за ней и мчится по перекату, нанося удары лапами и хватая рыбу зубами. Горбушу медведь поджидает на перекатах и стремительным ударом оглушает ее и хватает зубами. Кету ловит на нерестилищах в ключах, идя по руслу нерестилища и хватая рыбу зубами. Из сказанного видно, что наши дальневосточные бурые медведи хорошо научились ловить рыбу, что передается у них от поколения к поколению. Томилин [1974] отмечает, что у дельфинов родители обучают детенышей приемам, как ловить и поедать добычу. Шовен [1972] пишет, что ягнята, выкормленные козами, брыкаются задними ногами, как козы (что несвойственно овцам) и впоследствии боятся других овец. Таких примеров можно привести очень много.
Биологическое значение опосредованного обучения
Биологическое значение опосредованного обучения во всем его комплексе очень велико. Оно проникает у птиц и млекопитающих даже в сексуальное и в материнское поведение. Например, как было показано в опытах Харлоу (Harlow M., Harlow Н., 1962) с обезьянами, биологически нормальный самец, выращенный в изоляции от других особей своего вида, достигнув половой зрелости оказывается неспособным к нормальному для вида сексуальному поведению. Самка же, выращенная на искусственном кормлении под искусственной моделью «матери», не может правильно кормить своих детей и заботиться о них, а нередко и уничтожает их. Как пишет по этому поводу И. Кон [1970], кроме инстинкта здесь требуется еще какое-то «научение», осуществляемое в процессе общения с себе подобными. Миссакиан [Missakian, 1969] также отмечает, что резусы (Масаса mulatta), изолированные с детства, сохранили половое влечение и делали попытки к совокуплению, но не осуществляли полового акта. У них также отсутствовала взаимная чистка шерсти, хотя собственный шерстяной покров они чистили. Отсюда автор делает вывод, что изоляция сказывается не на влечении и не на моторике, а на организации общения между особями в процессе осуществления репродуктивных функций [цит. по: П. В. Симонов, 1975]. Самцы ряда уток, выведенные в опытах этологов под утками других видов и подвергшиеся соответствующему запечатлеванию, предпочитают затем в ряде случаев спариваться с самками именно этих, а не своих видов. Именно опосредованным обучением можно объяснить относительно быстрое образование новых навыков у популяции в целом или у части популяции данного вида с резкими изменениями видовых стереотипов поведения. Можно привести несколько примеров. Интересные изменения произошли за последние десятилетия в питании больших пестрых дятлов (Dendrocopos major). Эти птицы, питавшиеся в основном древесными вредителями, частично перешли на выкармливание птенцов в теплые годы главным образом тлями, соскабливая их клювом с листьев и побегов (Благосклонов, 1976). В холодные дождливые годы, когда прекращается размножение тлей, многие дятлы начинают хищничать, разоряя гнезда мелких птиц, преимущественно (но не только) дуплогнездников и, убивая их птенцов, питаются ими сами и кормят своих птенцов. Можно думать, что это явление так или иначе связано с уменьшением в культурных лесах старых деревьев, зараженных вредителями. Во всяком случае, как отмечает К. Н. Благосклонов [1972], такое хищничество больших пестрых дятлов уже давно было известно в Западной Европе. Под Москвой оно было отмечено в начале 60-х годов и развивается все больше. За Уралом оно неизвестно. Хищничество дятлов имеет место только в отношении птенцов семейства воробьиных. На взрослых птиц оно не распространяется. Во всяком случае, пришлось наблюдать, как самец-дятел перетаскал неоперенных птенцов из гнезда дрозда-белобровика, а самку дрозда, которая, защищая гнездо, налетала на дятла, он убил ударом клюва в голову, но есть ее не стал. К. Н. Благосклонов [1972] пишет: «По-видимому, имеет место «научение» дятлов от одного к другому и этой «науке» обучаются все более восточные дятлы». По нашим наблюдениям, не все индивидуумы больших пестрых дятлов проявляют хищные наклонности. Так, в районе Абрамцева (Подмосковье) было отмечено особенно большое хищничество дятла, но после отлова хищничающего самца разорение гнезд почти прекратилось, хотя в этом же районе держались и кормились еще две пары этих птиц. Не проявляла этих наклонностей и оставшаяся самка отловленного дятла. Весьма интересно, что хищное поведение дятлов отразилось на поведении других птиц: мухоловки серые и пестрые и дрозды-белобровики и рябинники начали нападать вблизи своих гнезд на больших пестрых дятлов и стараться прогнать их. Я установил этот факт раньше, чем мне стало известно о хищничестве дятлов. Фишер и Хинде [Fisher, Hinde, 1948] описывают, как большие синицы в Англии научились протыкать клювами картонные крышки у бутылок со сливками, которые торговцы оставляли утром у дверей домов своих клиентов, и выпивать часть сливок. По этому поводу Р. Шовен [1972] писал: «Этот трюк, «изобретенный» отдельными птицами, переняли другие, так что он широко распространился в довольно большом районе, и молочники уже не осмеливались оставлять по утрам молоко у дверей домов. Вероятно, первые синицы научились этому методом проб и ошибок, а остальные — подражая первым». В наших поселках и городах нередко серьезной помехой хозяйкам бывают синицы-«форточницы», научившиеся расклевывать свертки с продуктами, вывешиваемые зимой за окно. Следует вспомнить историю образования хищного поведения у новозеландского попугая кеа (Nestor notabilis). Эта некрупная птица (размером с ворону) живет в горных районах Новой Зеландии и питается обычно растительной пищей и мелкими животными. Однако в связи с развитием овцеводства некоторые кеа начали проявлять хищнические наклонности, нападая на овец и нанося им своим мощным клювом огромные раны в области спины. Овцы от этого погибают и их мясо поедается попугаями. Первые случаи такого хищничества были отмечены в 1868 г. [Иорданский, 1973]. Можно предполагать, что ему предшествовали случаи «чистки» овец попугаями от паразитов. При «чистке» ранки на коже овец кровоточили, и кровь пришлась, вероятно, по вкусу попугаям. Затем хищничество кеа приняло довольно широкие масштабы, причем, видимо, большое значение имело опосредованное обучение. Интересный пример быстрого изменения особенностей поведения грача (Corvus frugilegus L.) приводит Т. Б. Ардамацкая [1967]. Грачи, гнездящиеся в лесах Черноморского заповедника, всегда питались в основном насекомыми — вредителями сельского хозяйства и вдруг у обособленно живущей группы, гнездящейся на островке Соленоозерного участка, стали проявляться хищнические наклонности. Эта группа грачей жила в смешанной колонии с малой белой цаплей. У этих двух видов нередко возникали драки, во время которых разбивались яйца и расклевывались птенцы в гнездах. В 1964 г. грачи начали посещать о-в Орлов в Тендровском заливе и спокойно расклевывать лежащие в гнездах яйца чаек. Особенно большой урон они наносили гнездам черноголовых чаек, которые при тревоге не защищают своих кладок. Так «грачи-хищники», по выражению Т. Ардамацкой, приспособились к добыче высококалорийного и легкодоступного корма. Как отмечает С. В. Мараков [1969], после акклиматизации ондатры на озере Балхаш кабаны в зимнюю пору начали вредить вселенцу, причем отдельные семьи кабанов даже специализируются на разорении ондатровых жилищ и поедании запасов их корма. При этом также можно предполагать роль опосредованного обучения. Киккава [Kikkava, 1969] описывает, что одна небольшая популяция ондатр легко переносила недостаток пищи зимой, запасая с осени кукурузу, которую зверьки таскали с соседнего поля. Эта полезная для популяции особенность поведения передавалась из поколения в поколение путем подражания. Когда после одной исключительно суровой засухи большая часть этой популяции погибла, новое поколение не научилось использовать кукурузу и зимняя смертность этой популяции была очень высокой. В антропогенный период филогенеза происходят особенно быстрые изменения поведенческих адаптации животных. Человек так или иначе все сильнее вмешивается в жизнь животных нашей планеты. Он ускоряет естественный эволюционный процесс и придает ему новые направления [Иоганзен,, 1959]. Одних животных он истребляет, другим изменяет условия жизни, третьих охраняет, одомашнивает и разводит. Очевидно, скорость возникновения морфологических адаптации, если она не подкреплена искусственным отбором, не соответствует темпам развития деятельности человека, поэтому такие морфологические изменения очень редки. Основные адаптации у диких животных к этой деятельности человека развиваются главным образом по линиям общефизиологических (в первую очередь плодовитость) и поведенческих реакций [Мантейфель, 1974]. У животных ветви вторичноротых эти поведенческие адаптации создаются прежде всего в направлении опосредованного обучения (сигнальная преемственность и стайное обучение). Можно привести огромное количество таких примеров поведенческих адаптации к деятельности человека. Как дикие (мизантропные) виды, так и синантропные [условные, частичные и полные, по классификации Ю. А. Исакова, 1968, 1969] во многом адаптировались к человеку, изменили свое поведение соответственно его действиям. Многие преследуемые человеком промысловые животные выработали по отношению к нему сложные оборонительные особенности поведения. Они хорошо учитывают дальность действия современного охотничьего оружия, умело прячутся и запутывают свои следы. Нередко лисы, преследуемые гончими собаками в населенных местностях, используют шоссе и железные дороги для запутывания своих следов, что часто вызывает гибель собак. Волки и лисицы хорошо усвоили опасность капканов и запаха железа; в то же время лисы свободно роются на городских свалках и у туристских становищ среди консервных банок и других металлических предметов. Д. И. Бибиков [1977] указывает, что при сокращении (от охотников) численности волков они делаются чрезвычайно осторожными, воспитывая это качество и в своем потомстве. Во многих местах стая волков обычно уходит от недоеденной добычи за десятки километров, что делает невозможным уничтожение их из засады, а также при помощи капканов и ядов. В тундре при отстреле с самолетов волки ложатся, маскируясь от авиаохотников в тени от неровностей рельефа. В то же время в Кавказском заповеднике волки не боятся людей, следят за туристами и подбирают за ними остатки пищи. Следует заметить, что изменения в поведении позвоночных животных очень четко отражает отношение человека к данным животным. Они не только быстро научаются (опосредованное обучение) бояться человека и не подпускать его на определенное расстояние, но и, наоборот, при хорошем отношении перестают его бояться и даже часто используют для возможности так или иначе получить пищу. При этом оборонительная реакция на человека или на его технику сменяется безразличной или положительной — пищевой. Достаточно вспомнить, как близко подпускают к себе человека и автомашину звери больших африканских заповедников (что описано в ряде книг, например Феликсом Род-ригес Де ла Фуэнтэ [1972]), как роются рядом с гостиницами на помойках больших национальных парков Америки медведи и другие животные. Белки в некоторых наших парках, как, например, в Ботаническом саду в Москве, в садах около Московского государственного университета, а также в садах и даже на улицах Новосибирска, Риги и других городов, подпускают человека к себе и берут пищу из рук. В 1977 г. мне неоднократно приходилось наблюдать, как в саду, неподалеку от Московского государственного университета, белки подходили к людям, брали у них из рук корм и даже вскакивали к ним на руки и на одежду. Также доверчиво вели себя здесь большие синицы, гаички и поползни. При этом решающее значение имеет опосредованное обучение. Для изучения этого явления мною были произведены в районе Абрамцева (Московская область) эксперименты по приручению больших синиц (Parus major). Первоначально были приучены к съемной кормушке пять выводков синиц (несколько десятков экземпляров). Затем кормушка была поставлена на стол и экспериментатор сел рядом, положив руки около кормушки. Синицы быстро (за 4 часа) привыкли и начали брать корм (подсолнухи) рядом с руками. На следующий день кормушку убрали и корм положили прямо на руки. Через час уже пять синиц стали брать корм, садясь на руки. Остальные сидели недалеко на ветвях дерева, наблюдая за действиями наиболее смелых. Спустя 3 часа корм стали брать около десяти синиц. На третий же день, когда экспериментатор появился у стола, к нему навстречу полетело не менее двадцати синиц, которые садились на плечи, руки и голову, требуя еды, и дрались между собой. Среди этих синиц были уже не только молодые, но и старые. Так за 2 дня было приручено несколько десятков синиц, которые и дальше продолжали иметь резко сниженную оборонительную реакцию к человеку, а особенно к данному экспериментатору и подлетали к нему не только в этом месте, но и встречаясь с ним в лесу. Самое интересное происходило тогда, когда у кормушки появлялась новая взрослая синица, не проходившая процесса приручения. Новая синица боялась экспериментатора и при его приближении отлетала в сторону, откуда наблюдала, как ручные синицы садились на человека и брали подсолнухи. Новая синица сначала при этом очень волновалась и издавала крики предупреждения и тревоги, а затем опускалась на ветви, приближаясь к экспериментатору, а через час-полтора вслед за другими подлетала к руке и, не садясь на нее, держась порхающим полетом, схватывала семечко. Вскоре после этого новая синица начинала садиться на руку. Здесь, несомненно, имело основное значение опосредованное обучение и пример соседей, поскольку все попытки приручить таких старых синиц в одиночку не давали успеха. Следует отметить, что на следующее лето у кормушки появлялось несколько прошлогодних прирученных синиц, у которых сначала рефлексы приручения были несколько уменьшены, но очень быстро восстановились. При этом старая самка, гнездившаяся в этом районе три года, очень быстро научилась «выпрашивать» пищу для выкармливаемых ею в дуплянке птенцов: она подлетала к экспериментатору и трепетала крыльями так, как обычно делают самки перед самцом, производя ювенильную сигнализацию. Получив корм (вареное яйцо, сыр), она несла его в дуплянку кормить птенцов. Интересно, что впоследствии обучение птенцов родителями «на человека» не происходило: взрослая синица безбоязненно садилась на руку и брала корм, но летным птенцам не разрешала этого делать и сигналами тревоги заставляла их подниматься на ветви дерева. Таким образом, каждый год приручение молодых синиц приходилось проводить заново, когда семейные группы распадались. Можно предполагать, что для родителей синиц человек продолжал в какой-то степени оставаться потенциальной опасностью в отношении их птенцов. В то же время у прирученной молоди наблюдалась тенденция искать спасения у человека. Так, когда появился ястреб-перепелятник и раздался сигнал опасности, три молодых синицы бросились к сидящему экспериментатору и спрятались у него под ногами. А. Н. Формозов [1972] приводит интереснейшие наблюдения за поведенческими адаптациями животных к человеку. Так, он отмечает, что в 1948 г. в Актюбинской области лисы выходили к железнодорожному полотну неподалеку от тех станций, на которых шла торговля копченой и вяленой рыбой, и ждали, когда из окон проходящего поезда пассажиры выбрасывали рыбные отходы. Он же описывает, как ласточки летают около идущего человека и следуют за повозкой или автомашиной, схватывая насекомых, вылетающих из травы. Подобно этому, сокол-чеглок ловит мелких птиц, выпугиваемых идущим поездом. В. Э. Якоби [1972] отмечает, что ласточки летят за реактивным самолетом, едущим по дорожке, и ловят насекомых, поднятых из травы струей газов двигателя. Следует отметить, что относительно недавно появившаяся техника сначала вызывала резкую оборонительную реакцию у многих животных, но затем в результате опосредованного обучения довольно быстро превратилась не только в индифферентный раздражитель, но в ряде случаев стала служить и положительным пищевым условным раздражителем. Прежняя идиллическая картина пахаря и сопровождающих его грачей теперь заменилась трактором, идущим с плугами. Стрекотание такого трактора привлекает животных, охотящихся за червями, личинками насекомых и мелкими грызунами. За такими тракторами движутся часто целые колонны грачей, галок, ворон, речных и сизых чаек. Его сопровождают пустельги и кобчики, а нередко и азартно охотящиеся за грызунами лисицы. При этом, по нашим наблюдениям, врановые быстро устанавливают, в какой части поля держится наиболее богатая почвенная фауна, и следуют за трактором именно на этом участке. По наблюдениям Э. Н. Головановой и Ю. Б. Пукинского [1971а], грачи в Каракалпакии не следуют за трактором на пахоте, а ожидают полива земель и охотятся за насекомыми, выползающими из заливающей их воды. Э. Н. Голованова [1975] пишет: «Особенно много птиц концентрируется у посевов трав во время покоса. Следом за косилкой двигаются и кормятся грачи, вороны, скворцы, аисты, майны, сизоворонки. Ласточки ловят спугнутых мелких насекомых, а остальные птицы поспешно хватают все живое, что внезапно оказывается на поверхности скошенных трав. Так же как покос, птиц привлекает на посевы трав полив. Грачи, галки, белохвостые пигалицы в Каракалпакии, майны, сороки, сизоворонки и удоды в Туркмении собираются на участках полей, где только начинается полив. Птицы, следуя вдоль края медленно наступающей воды, ловят насекомых, выползающих на поверхность». Во многих местах побережья Балтики, и в частности в г. Таллине, чайки нескольких видов научились питаться отбросами на городских свалках, выхватывая наиболее лакомые куски прямо из-под ковша экскаватора, сгребающего мусор (сообщение В. Э. Якоби). По сообщению В. И. Цветкова, в порту Малокурильска при выгрузке с судов открытых ящиков с сайрой вороны (Corwus corax) и серебристые чайки (Larus argentatus) собираются к транспортерным лентам и спокойно едут на них, поедая сайру из движущихся ящиков. Многие птицы и млекопитающие связали свое поведение с техникой рыбных промыслов. Так, глупыши (Fulmarus glacialis) живут в северных морях в основном за счет отходов тралового лова. Так же ведут себя в различных морях и многие виды чаек [HilJis, 1971], для которых звук работающих на судах лебедок (выборка трала, сетей, кошелькового невода) служит пищевым раздражителем [Мантейфель, 1965]. Эти птицы в массах скапливаются около промышляющих рыболовецких судов. В прекрасном документально-научном кинофильме, снятом из подводного батиплана «Атлант» сотрудником Атлан НИРО В. К. Коротковым в Средиземном море, хорошо видно, как дельфины афалины преследуют идущий под водой трал с уловом и вытаскивают зубами рыбу через ячею его кутка. В. В. Дежкин и С. В. Мараков [1968] пишут, что морские львы (сивучи Eumetopias jubatus) у берегов Камчатки в последнее время привыкли забираться в идущие в воде тралы и пожирать рыбу. Даже будучи вытащенными в трале на борт судна, они ведут себя довольно независимо и прыгают обратно в море. Как пишут эти авторы, «сивучи освоили траловый лов». Интересные изменения происходят в поведении обыкновенного скворца (Sturnus vulgaris). Эта птица, сильно тяготеющая к человеку (частичный синантроп), отличается необычайной пластичностью поведения. Гнезда скворца можно встретить и в дуплах деревьев, скворечниках, под застрехами домов в городах; их обнаруживали также в антенне действующего радиолокатора [Якоби, 1969], в контргрузе работающего экскаватора [Якоби, 1970] и в других местах. В. И. Харченко [1970] отмечает, что с 1965 г. скворцы начали оставаться на зимовку в ряде южных областей СССР. Известно, что большие стаи скворцов остаются на зимовку и в Москве. Харченко описывает, как зимующие скворцы устраивают массовую ночевку на деревьях, растущих на центральной улице Ростова-на-Дону. Тысячи скворцов облепляют здесь ветви деревьев, не опасаясь необычной обстановки этого большого, густонаселенного города. Зимующие скворцы питаются в основном на городских свалках всевозможными отбросами и не упускают возможности поохотиться за мышевидными грызунами. Обычно, пишет Харченко, в такой охоте принимают участие сразу несколько птиц, нанося грызуну удары клювом. Убив мышь, скворцы начинают ее расклевывать. Переход скворцов к такому хищничеству сказывается на их паразитофауне, поскольку на них все в большем количестве встречаются эктопаразиты мышевидных грызунов. Далее Харченко отмечает, что летом при выкармливании птенцов некоторые скворцы, живущие в населенных пунктах Ставрополья, Ростовской области и Калмыкии, добывают пищу не только днем, но и в сумерках и даже ночью, охотясь у зажженных фонарей на слетающихся на свет насекомых. Кроме того, скворцы в этот период часто питаются и обитателями прудов и рек (в Донецке, Ростове-на-Дону) — как насекомыми и головастиками, так и мелкой рыбешкой. При этом, по описанию Харченко, одни скворцы, стремительно пролетая над поверхностью воды, выхватывают из нее всплывших к поверхности мальков; другие же, «как заправские ихтиофаги, на несколько секунд зависают в 3 — 5 м над водой, затем следует стремительный бросок вниз — и вот уже из каскада брызг появляется мокрый скворец с трепещущей серебристой полоской в клюве. Через несколько минут добыча расклевана и скормлена выводку» [Харченко, 1970, с. 99]. Этот же автор отмечает образование больших стай скворцов во время весенней и осенней путин на рыбоприемных пунктах на берегах Азовского и Каспийского морей и на других водоемах Предкавказья. Здесь на рыбоприемных пунктах сотенные стаи кормятся у транспортеров и на грудах наваленной рыбы, расклевывая у рыб брюшко и выклевывая глаза. Для дополнительной характеристики поведенческой пластичности скворца могу привести факт, сообщенный мне в 1973 г. В. М. Никольским: под Керчью в огромном механическом цехе одного из заводов в зимнее время собираются тысячи скворцов. Рядом с цехом находится заводская столовая и рабочие, возвращаясь с обеда, несут им всякую снедь. Скворцы при этом бросаются к людям, садятся к ним на плечи и головы и ведут себя как совершенно ручные птицы. Такая необычайная пластичность поведения обыкновенного скворца позволяет этим птицам широко использовать деятельность человека, в связи с чем численность вида непрерывно растет. Много примеров адаптивного изменения поведения можно видеть в процессе урбанизации лесных птиц, причем, как отмечает А. С. Мальчевский [1969], имеет место особенно жесткий отбор, идущий в направлении формирования пластического поведения особей: в условиях людного парка (речь идет о парке Лесотехнической академии Ленинграда) птицы могут изменять характер своего поведения у гнезда, суточный ритм питания, стереотип гнездования и т. п. К. Н. Благосклонов [1975] в своей интересной статье о птицах большого города также приводит целый ряд примеров таких изменений особенностей поведения, и в частности гнездования птиц в условиях урбанизации. Серые вороны (Corvus cornix L.) раньше гнездились обычно в лесах, где вели себя очень скрытно и осторожно. Теперь же очень часто можно видеть гнезда ворон, находящиеся на деревьях, расположенных на людных улицах и перекрестках г. Москвы (например, на Ленинском проспекте). Не менее интересно происходит освоение городских зданий как гнездовых стаций чаек в Великобритании и Ирландии [Cramp Stanley, 1971]. Еще в 1940 г. гнездование чаек на зданиях в этих странах было почти неизвестно. Учет гнездящихся на зданиях чаек, произведенный в 1969 и 1970 гг., показал, что такое изменение гнездового поведения произошло у пяти видов. Наиболее широко урбанизирована серебристая чайка (L. argenta-tus), у которой учтено более тысячи гнезд на зданиях. Большинство колоний этого вида располагается на крышах домов, лавок, контор. Гнезда устраиваются часто также между колпаками дымовых труб, на опорах различных мостов. Клуша (Larus fuscus) гнездится сходно с серебристой чайкой. Моевка (Rissa tridactyla) начала использовать постройки человека раньше других чаек. Изменению гнездового поведения сопутствует и изменение пищевого поведения: чайки питаются на городских свалках и в портах. Аисты часто стали гнездиться на столбах электропередач. Резко меняется поведение по отношению к технике и у домашних животных. Н.М. Носков [1972] описывает, как за последние 50 лет изменилось отношение домашнего скота к автотранспорту — от панического страха к полностью индифферентному поведению. Он отмечает, что в Кабардино-Балкарии ослята и телята приучились греться группами на шоссе, не обращая внимания на подъезжающие автомобили и создавая тем самым большие помехи для автотранспорта. Таких примеров можно привести очень много.
Практичне заняття №3. Розвиток психіки тварин в онтогенезі.
Мета: вивчення розвитку психічної діяльності, рухової активності, сенсорних можливостей та елементів спілкування у пренатальному періоді; особливостей постнатального розвитку поведінки тварин, пізнавальних аспектів раннього постнатального періоду, формування спілкування, “дослідницької” поведінки та орієнтації. Вивчення особливостей розвитку психічної діяльності в ювенільному періоді, удосконалення рухової активності та формування спілкування в іграх тварин, пізнавального значення ігрової активності та порівняльно-психологічних аспектів ігор тварин і людини.
Форма заняття: семінар.
Обладнання: методичні рекомендації, навчальні посібники.
План заняття
Загальна характеристика сенсорних і рухових реакцій в пренатальному періоді.
а) „Ембріональне навчання” і дозрівання.
б) Особливості пренатального періоду у безхребетних, хребетних,
птахів, ссавців.
в) Значення ембріогенезу для розвитку психічного відображення.
2. Розвиток поведінки в ранньому постнатальному періоді.
а) Розвиток поведінки у зрілонароджених і незрілонароджених тварин.
б) Значення турботи про потомство.
в) Раннє облігатне і факультативне навчання у безхребетних і хребетних.
г) Раннє формування спілкування у нижчих і вищих хребетних.
3. Розвиток в ювенільному (ігровому) періоді.
а) Основні концепції гри у тварин. Концепція К.Е.Фабрі.
б) Гра і спілкування.
в) Пізнавальне значення гри у тварин.
Завдання для самостійної роботи по підготовці до заняття:
Опрацювати літературу за питаннями плану семінару.
Підготувати короткий конспект-відповідь на питання семінару.
Скласти глоссарій: онтогенез, пренатальний період, постнатальний період, ювенільний період, ігрова діяльність, сенсорна депривація, маніпуляційні ігри, локомоторні ігри, трофейні ігри, статеве закарбування, реакція слідування, вроджене впізнання.
Теми рефератів
Розвиток поведінки хребетних в онтогенезі.
Статеве зафіксування та його значення для статевої поведінки хребетних.
Подібність та різниця онтогенезу зріло- та незрілонароджених ссавців.
Навчання в різні періоди онтогенеза.
Типи шлюбних відносин у тварин
Роль акустичної комунікації в статевій поведінці.
Роль зорової комунікації в статевій поведінці.
Типи угруповань тварин.
Гра тварин у природі і розуміння її людиною. Форми, функції, ігрова комунікація.
Порівняльна характеристика гри тварин та дітей.
Література.
Дьюсбери Д. Поведение животных. М.: Мир, 1981.
Правоторов Г.В. Зоопсихология для гуманитариев. Новосибирск: ЮКЗА, 2001.
Фабри К.Э. Основы зоопсихологии. М.: Изд-во Моск ун-та, 1993.
Фабри К.Э. Игра у животных. М.: Знание, 1985. (Новое в жизни, науке, технике. Сер. «Биология», № 8).
Фабри К.Э. Игры животных и игры детей // Вопросы психологии. 1982. № 2. С. 26–34.
Крымов А.А. Проблема игры животных в современной зоопсихологии //Психол. Журнал. 1982.№3. с.132-139.
Хайнд Р. Поведение животных. М.: Мир, 1975.
З.А.Зорина, И.И.Полетаева. Элементарное мышление животных. М.: Аспектпресс. 2003 г.
Леонтьев А.Н. Проблемы развития психики. М., 1965. С. 209–261.
Тестові завдання
1. Вкажіть яку роль в еволюції відіграє вроджене впізнання:
а)сприяє формуванню видоспецифічної поведінки,
б) полегшує пристосування до постійних змін зовнішнього середовища,
в) полегшує пристосування до періодичних змін зовнішнього середовища,
г) полегшує пристосування до раптових змін зовнішнього середовища.
2. Основні періоди онтогенезу тварин:
а) пренатальний,
б) період дитинства,
в) ранній постнатальний ,
г) ювенільний,
д) старість.
3. Представники концепції гри як „виходу надмірної енергії”:
а) К.Гроос,
б) Г.Спенсер,
в) К. Лоренц,
г) К.Ллойд Морган,
д) Д.Моріс,
ж) А.Д.Слонім.
4. Який з перерахованих періодів є ігровим?
а) постнатальний,
б) ювенільний,
в) статевої зрілості,
г) пренатальний.
5. Ігри, при яких мають місце узгоджені дії хоча б двох партнерів:
а) спільні ігри,
б) ритуалізовані форми поведінки,
в) „запрошення до гри”,
г) ієрархічні ігри.
6. Сучасна концепція гри, яка розроблена К.Е.Фабрі:
а) пропонує ендогенну мотивацію ігрової поведінки,
б) стверджує, що гра – це тренування,
в) пропонує синтетичний підхід до розуміння гри,
г) гра – розвиваюча діяльність, яка охоплює більшість функціональних сфер.
7. Ранній постнатальний період:
а) формуються найважливіші життєво важливі взаємовідношення з середовищем,
б) закладаються основи дорослої поведінки,
в) найбільш чутлива фаза для навчання,
г) тварина повністю самостійна.
8. Автор концепції гри як „действия вхолостую»:
а) К.Гроос,
б) Г.Спенсер,
в) К.Лоренц,
г) К.Ллойд-Морган,
д) Д.Моріс,
ж) А.Д.Слонім.
9. Маніпуляційні ігри якісно змінюють у тварин:
а) емоційну сферу,
б) сенсорну сферу,
в) моторну діяльність,
г) перцептивну діяльність.
10. Вирішальний фактор в різноманітності протікання постембріонального періоду:
а) наявність середовища,
б) наявність їжі,
в) освітленість,
г) ступінь зрілонародженості особини.
11. Маніпуляційні ігри:
а) шкодять положенню в ієрархічній групі,
б) ігри молодих тварин з предметами,
в) маніпулювання іншою твариною в угрупованні,
г) розвиваюча діяльність,
д) гальмують розвиток особини,
ж) збагачують рухову активність тварин.
12. Відмітьте слово або словосполучення, яке на Ваш погляд не пов’язане з іншими:
а) пасивно оборонна реакція,
б) ювенільний період,
в) ігрова поведінка,
г) імпринтинг.
13. Система поведінкових зв’язків між особинами в групі, яка регулює їх взаємовідносини:
а) структурованість,
б) інтеграція,
в) ієрархія,
г) угруповання.
14. Ігрова активність пов’язана з інстинктами:
а) вітальними,
б) зоосоціальними,
в) статевими,
г) саморозвитку.
15. Автор концепції гри як „спонтанної” діяльності, яка викликається підпороговими подразниками:
а) К.Гроос,
б) Г.Спенсер,
в) К.Лоренц,
г) К.Ллойд-Морган,
д) Д.Моріс,
ж) А.Д.Слонім.
16. В іграх тварин проявляються особливості:
а) статевої поведінки,
б) поведінки добування їжі,
в) пошукової поведінки,
г) способу життя.
17. У вищих тварин чутливий період для облігатного навчання:
а) ранній постнатальний,
б) ювенільний,
в) пренатальний,
г) адуальний.
18. Представники концепції „гри як практики для дорослої поведінки”:
а) К.Гроос,
б) Г.Спенсер,
в) К.Лоренц,
г) К.Ллойд-Морган,
д) Д.Моріс,
ж) А.Д.Слонім.
19. Ступінь зрілонародження:
а) залежить від рівня філогенетичного розвитку особини,
б) власної ваги,
в) способу життя,
г) власного розміру.
20. Суть концепції гри як”действия вхолостую»:
а) інстинкти, які проявляються за відсутністю релізерів ,
б) удосконалення спадкових форм поведінки,
в) „вакуумна активність”,
г) практика для дорослої поведінки,
д) „спонтанна” діяльність, яка викликається підпороговими подразниками,
ж) фаза розвитку поведінки в стадії формування.
21. Відмітьте слово або словосполучення, яке на Ваш погляд не пов’язане з іншими:
а) олені,
б) імпринтинг,
в) реакція слідування,
г) період новонародженості.
22. Раннє формування просторової орієнтації пов’язано:
а) тільки з облігатним навчанням,
б) тільки з факультативним навчанням,
в) генетичними програмами,
г) з поєднанням облігатного та факультативного навчання.
23. Спадкові програми проявляються в пренатальному періоді:
а) в побудові організму, його органів і систем,
б) в запуску всіх функцій організму,
в) в становленні рефлекторних функцій,
г) готують морфофункційну основу для психічної діяльності в зовнішньому середовищі.
24. Здатність тварини до самостійного виконання життєво важливих функцій після народження:
а) готовність,
б) зрілонародженість,
в) зрілість,
г) метаморфоз.
25. Психічно найбільш розвинені тварини:
а) зазвичай відносяться до зрілонароджених,
б) завжди відносяться до незрілонароджених,
в) не залежать від ступеню зрілонародженості,
г) в більшості відносяться до незрілонароджених.
26. Психічна діяльність ембріона:
а) відсутня,
б) існує в зародковій формі,
в) відношення з зовнішнім середовищем формуються,
г) відношення з зовнішнім середовищем сформовані.
27. Властивості гри:
а) елементи дослідницької активності,
б) існує „центр гри” в мозкових структурах,
в) ігрові дії виконуються багаторазово та невтомно,
г) існують „органи” гри.
28. Гра:
а) не „зразок”, дорослої поведінки, а сама поведінка в процесі становлення
б) формування нових моторних програм поведінки,
в) навчальна форма поведінки,
г) є складним інстинктом саморозвитку з певним енергетичним еквівалентом.
29. Дії тварин, які направлені на забезпечення чи покращення умов виживання і розвитку потомства:
а) виховання,
б) турбота про потомство,
в) годування,
г) ігрова поведінка.
30. Першими у лисенят та барсучат зявляються маніпуляції, які виконуються:
а) передніми кінцівками,
б) ротовим апаратом і головою,
в) однією передньою лапою,
г) задніми кінцівками.
31. Впізнання базується на:
а) таксисах,
б) сигнальному наслідуванні,
в) імпринтингу,
г) умовних рефлексах.
Складний комплекс різних поведінкових актів, який в сукупності складає поведінку молодих тварин на етапі онтогенезу, що передує статевому дозріванню:
а) інстинкт,
б) адаптивна поведінка,
в) ігрова активність,
г) пошукова діяльність.
Практичне заняття № 4. Еволюція психіки.
Мета: ознайомитися з характеристикою нижчого та вищого рівнів елементарної сенсорної психіки, поняттям пластичності поведінки і психічного відображення на вищому рівні елементарної сенсорної психіки. Ознайомитися з характеристикою нижчого та вищого рівнів перцептивної психіки. Порівняльняти особливості психіки на різних еволюційних рівнях.
Форма заняття: семінар.
Обладнання: методичні рекомендації, навчальні посібники.
План заняття
Стадія елементарної сенсорної психіки.
а) Загальна характеристика нижчого рівня елементарної сенсорної психіки;
б) Рухова активність і її просторова орієнтація у найпростіших;
в) Кінези і елементарні таксиси;
г) Проблема пластичності поведінки найпростіших. Явище звикання.
д) Загальна характеристика вищого рівня елементарної сенсорної психіки;
е) Поява нервової системи і основні етапи її розвитку;
є) Рухова активність нижчих багатоклітинних безхребетних;
ж) Зачатки вищих форм поведінки у кільчастих червів;
з) Пластичність поведінки і психічне відображення на вищому рівні елементарної сенсорної психіки.
2. Стадія перцептивної психіки
а) Загальна характеристика нижчого рівня перцептивної психіки;
б) Рухові і сенсорні здібності вищих безхребетних. Особливості будови їх нервової системи і органів чуття.
в) Предметне сприйняття на нижчому рівні перцептивної психіки.
г) Спілкування і групова поведінка у вищих безхребетних; територіальна поведінка;
д) Інстинкт і навчання в поведінці комах;
е) Загальна характеристика вищого рівня перцептивної психіки.
є) Центральна нервова система вищих хребетних;
ж) Локомоція і маніпулювання у вищих хребетних;
з) Сенсорні здібності у вищих хребетних;
и) Спілкування;
і) Ригідність і пластичність в поведінці вищих хребетних.
3. Проблема інтелекту тварин.
а) Передумови і елементи інтелектуальної поведінки тварин;
б) Критерій інтелектуальної поведінки тварин;
в) Форми мислення;
г) Біологічна обмеженість інтелекту тварин.
Завдання для самостійної роботи по підготовці до заняття:
Опрацювати літературу за питаннями плану семінару.
Підготувати короткий конспект-відповідь на питання семінару.
Ознайомитися з роботою Тушмаловой Н.А. Основные закономерности эволюции поведения беспозвоночных. Наводити приклади.
Скласти глоссарій: сенсорна психіка, перцептивна психіка акустична коммунікація, візуальна коммунікація, вроджене впізнання, грумінг, депривація.
Заповнити таблицю: Стадії і рівні психіки та поведінки тварин (за А.Н. Леонтьєвим і К.Е. Фабрі).
Таблиця 1
Стадії і рівні психіки та поведінки тварин (за А.Н. Леонтьєвим і К.Е. Фабрі).
Стадії і рівень психічного відображення, його характеристика |
Особливості поведінки, що пов’язані з даною стадією і рівнем |
Види живих істот, які досягли цього рівня розвитку |
І. Стадія елементарної сенсорної психіки. |
||
А. Нижчий рівень. Примітивні елементи відчуттів. Розвиток подразливості.
|
А. |
А. |
Б. Вищий рівень. Наявність відчуттів. Поява важливого органу маніпулювання – щелеп.
|
Б. |
Б. |
ІІ. Стадія перцептивної психіки. |
||
А. Нижчий рівень. Відображення зовнішньої дійсності в формі образів предметів. Інтеграція, обєднання властивостей в цілісний образ предмета. Головний орган маніпулювання – щелепи.
|
А. |
А. |
Б. Вищий рівень. Елементарні форми мислення (розвязок задач). Формування певної «картини світу».
|
Б. |
Б. |
В. Найвищий рівень. Виділення в практичній діяльності особливої, орієнтовно-дослідницької, підготовчої фази. Здатність вирішувати завдання різними способами. Перенос відомого способу рішення задачі в нові умови. Створення і використання в діяльності примітивних знарядь праці. Здатність до пізнання довкілля незалежно від наявних, біологічних потреб. Врахування причинно-наслідкових звязків між явищами в практичних діях (інсайт). |
В. |
В. |
Теми рефератів
Рівні розвитку психіки.
Вроджені та набуті комунікативні процеси.
Переваги комунікації.
Знаки для понять; можливості та обмеження звукової сигналізації у приматів.
Взаємодія домовного мислення і комунікативних процесів.
Невербальна комунікація у людини.
Навчання вищих мавп «розмовляти».
Джерела мови людини. Мова та пізнання.
Розумні мавпи.
Розвиток поведінки в онтогенезі мавп і людини.
Стадні відношення і способи спілкування мавп.
Маніпулювання мавп з предметами і форми їх діяльності.
Навички та інтелект мавп.
Тестові завдання
Відмітьте слово або словосполучення, яке, на Ваш погляд, є „зайвим” в даному ряду:
а) Д. Фоссі,
б) Д. Шаллер,
в) Д. Гудолл,
г) горила.
Вкажіть, в якому періоді онтогенезу молоді тварини починають грати:
а) ювенільний,
б) період новонародженості,
в) період змішаного вигодовування,
г) період соціалізації.
Відмітьте слово або словосполучення, яке, на Ваш погляд, є „зайвим” в даному ряду:
а) кріт,
б) зір,
в) нюх,
г) дотик.
Відмітьте слово або словосполучення, яке, на Ваш погляд, є „зайвим” в даному ряду:
а) сексуальні ігри,
б) облігатне навчання,
в) статеве зафіксування,
г) зрілонародження.
Яка рухова реакція найпростіших є реакцією на дотик:
а) клінотаксис,
б) фототаксис,
в) геотаксис,
г) тигмотаксис.
У представників якої таксономічної групи можна вперше говорити про утворення умовних рефлексів?
а) кишковопорожнинні,
б) найпростіші,
в) кільчасті черви,
г) плоскі черви.
7. Вкажіть, хто із перерахованих тварин володіє перцептивной психікою?
а)синиця,
б) щука,
в) гідра,
г) слизняк.
8. Перший прояв психіки, який орієнтує організм в середовищі, виконує сигнальну функцію і є показником її виникнення:
а) подразливість,
б) чутливість,
в) реактивність,
г) рефлективність.
9. Вкажіть, психіка якої із перерахованих безхребетних тварин виходить за межі елементарної сенсорної?
а)дощовий черв,
б) виноградний равлик,
в) кальмар,
г) морська зірка.
10. Вкажіть, хто із перерахованих тварин володіє перцептивной психікою?
а) лисиця,
б) окунь,
в) амеба,
г) медуза.
11. Автором цитати „Стадия перцептивной психики характеризуется способностью отражения внешней объективной действительности уже не в форме отдельных элементарных ощущений, вызываемых отдельными свойствами или их совокупностью, но в форме отражения вещей” є:
а) К.Е.Фабрі,
б) А.Н.Леонтьев,
в) Л.Л.Крушинський,
г) Л.Г.Воронін.
12. Навчання найпростіших представляє собою:
а) вироблення умовного рефлексу,
б) асоціативне навчання,
в) неасоціативне навчання,
г) інструментальне навчання.
13. Вкажіть, психіка якої із перерахованих безхребетних тварин виходить за межі елементарної сенсорної?
а) інфузорія-туфелька,
б) медуза,
в) оса,
г) черв поліхета.
Вкажіть, психіка якої із перерахованих безхребетних тварин виходить за межі елементарної сенсорної?
а) морський їжак,
б) „устрица”,
в) мурашка,
г) дощовий черв.
15. Стадія елементарної сенсорної психіки характеризується здатністю відображення зовнішньої об’єктивної дійсності в формі:
а) відображення речей,
б) елементарних відчуттів,
в) елементарної чутливості,
г) предметного сприйняття,
д) відображення між предметних зв’язків і ситуацій.
16. Самим високим рівнем інтелекту із зазначених тварин володіють:
а) кішка,
б) курка,
в) собака,
г) кролик.
17. Вкажіть, хто із перерахованих тварин володіє перцептивной психікою нижчого рівня?
а) золота рибка,
б) віслюк,
в) ворона,
г) лисиця.
18. Вкажіть, хто із перерахованих тварин володіє перцептивной психікою вищого рівня?
а) горобець,
б) жаба,
в) інфузорія-туфелька,
г) дощовий черв.
19. Вкажіть, хто із перерахованих тварин володіє перцептивной психікою вищого рівня?
а) циклоп,
б) щука,
в) щур,
г) собака.
20. Елементарні інстинктивні рухи:
а) кінези,
б) таксиси,
в) тропізми,
г) локомоції.
21. Властивості нижчого рівня перцептивной психіки:
а) переважання жорстких програм,
б) групова поведінка,
в) спілкування,
г) ритуалізація,
д) елементарна розумова діяльність.
22. Перцептивна психіка:
а) психічне відображення в вигляді відчуттів,
б) супроводжується лише інстинктивною діяльністю,
в) сітчаста нервова система,
г) предметне сприйняття.
23. Самим високим рівнем інтелекту із зазначених тварин володіють:
а) білий щур,
б) черепаха,
в) дельфін,
г) собака.
24. Здатність до предметного сприйняття має повний розвиток у:
а) хребетних,
б) комах,
в) головоногих молюсків,
г) земноводних.
Три вершини розвитку перцептивной психіки:
а) хребетні,
б) комахи,
в) головоногі молюски,
г) земноводні.
26. У вищих представників безхребетних зачатки:
а) сприйняття,
б) агресивної поведінки,
в) спілкування,
г) елементарної розумової діяльності.
Література.
Дьюсбери Д. Поведение животных. М.: Мир, 1981.
Правоторов Г.В. Зоопсихология для гуманитариев. Новосибирск: ЮКЗА, 2001.
Фабри К.Э. Основы зоопсихологии. М.: Изд-во Моск ун-та, 1993.
Хайнд Р. Поведение животных. М.: Мир, 1975.
Кликс Ф. Пробуждающееся мышление. М.: Прогресс, 1983.
Леонтьев А.Н. Проблемы развития психики. М.: Изд-во Моск. ун-та, 1972.
Мак Фарленд Д. Поведение животных. Психобиология, этология и эволюция. М.: Мир, 1988.
З.А.Зорина, И.И.Полетаева. Элементарное мышление животных. М.: Аспектпресс. 2003 г.
О.Т. Губко. Основи зоопсихології. К. 2006.
Тушмалова н.А. Основные закономерности эволюции поведения беспозвоночных
Печатается по изд.: Хрестоматия по зоопсихологии и сравнительной психологии. Учебное пособие. 2-е изд. М.: Российское психологическое общество, 1998. С. 30-43. Цит. по изд.: Основные закономерности эволюции поведения беспозвоночных // Физиология поведения. Нейробиологические закономерности. Л.: Наука, 1987. С. 236-265 (с сокр.).
Одной из характерных особенностей современной биологии служит все возрастающий интерес к изучению поведения животных. По сравнению с позвоночными животными, закономерности формирования поведения беспозвоночных значительно менее изучены. Причем сказанное относится прежде всего к изучению их ВНД. Значительное отставание в изучении функциональных механизмов приобретенного поведения у беспозвоночных животных, на наш взгляд, может быть объяснено по крайней мере тремя основными причинами. Во-первых, трудностью содержания многих животных в лабораторных условиях; во-вторых, трудностью создания адекватных методик исследования и методов объективной регистрации поведения; и, в-третьих, тем, что физиологи, занимающиеся исследованием поведения беспозвоночных, далеко не всегда обладают знаниями, позволяющими свободно выбирать необходимые для изучения объекты (Тушмалова, 1973). Заметим сразу, что в филогенетическом ряду беспозвоночных наиболее изученными оказались насекомые и моллюски по сравнению с более "простыми" организмами. Так, для моллюсков и насекомых наиболее актуальной задачей является определение степени сложности механизмов приобретенного поведения, в то время как для низших беспозвоночных необходимо вновь и вновь доказывать, что они способны к накоплению индивидуального опыта — обучению. <...>
О физиологических механизмах поведения простейших
Об условных рефлексах простейших. Рассмотрение оригинальных экспериментальных исследований, посвященных выработке у простейших условных рефлексов, убеждает в чрезвычайно разноречивой трактовке результатов. Обзору таких работ посвящен ряд публикаций, отражающих взгляды биологов зоологического и физиологического профилей (Коган, 1963; Воронин и др., 1967; Серавин, 1969, 1978; Hamilton. 1975; Тушмалова, 1980). Исследователи, иллюстрирующие возможность выработки, условных рефлексов у простейших, чаще всего останавливаются на опытах Н. Н. Тимофеева (1958) и серии работ Гельбера (Gelber, 1956, 1958). Рассмотрим детально эти эксперименты, чтобы показать, что их результаты могут иметь иное объяснение, если учесть некоторые особенности ресничных инфузорий. Н. Н. Тимофеев проводил опыты на инфузориях в камере размером 10x4 мм, разделенной пополам, в каждой из половин были вмонтированы платиновые электроды. Условным раздражителем служил свет, безусловным — электрический ток (50 Гц 1 — 2 В). Выработка условного рефлекса заключалась в том, что инфузории "учились" только в ответ на изолированное действие света не заплывать в ту часть камеры, где давался удар электрического тока. Объясняя результаты опытов автор акцентирует внимание на двигательных реакциях инфузорий, считая, что выработка условных реакций определяется "количеством, а возможно, и качеством самой двигательной активности", не раскрывая этого понятия, однако опыты Н. Н. Тимофеева можно объяснить исключая возможность формирования временных связей. Известно, что многие инфузории в ответ на различные вредные воздействий внешней среды выстреливают трихоцисты1. Эту реакцию можно классифицировать как врожденную, безусловно оборонительную — сигнал "опасности" для других особей. В опытах Н. Н. Тимофеева таким "оборонительным" раздражителем был электрический ток. Формирование "условного рефлекса" во времени (постепенность выработки) на самом деле может быть следствием постепенного диффундирования относительно небольшого количества трихоцист по объёму камеры. Экспериментально показано, что выстреливание трихоцист происходит в ответ на непосредственное действие различных кислот, щелочей, механических воздействий электрического тока (Prosser, Brown, 1967). Таким образом, результаты опытов Н. Н. Тимофеева не могут служить доказательством способности простейших вырабатывать условные рефлексы. Часто как доказательство выработки у простейших условных рефлексов приводят серию работ Гельбера по выработке у инфузорий Paramecium aurelia пищевых условных рефлексов. В связи с принципиальной важностью физиологической оценки этих работ в плане обсуждаемой проблемы, остановимся подробно на рассмотрении результатов этих работ. Опыты проводились следующим образом. Стерильная платиновая проволока длиной 7,6 см с диаметром кончика 0,5 мм располагалась в середине стеклянной экспериментальной лунки под углом 85° к горизонтали. Поднимание и опускание проволоки осуществлялось с помощью специального рубильника. В контрольных экспериментах было показано, что погружение проволоки в нативную культуру2 на 3 мин приводит к скоплению на ней 1 — 2 инфузорий. При обучении проволоку опускали в лунку с голодными инфузориями на 15 с. с интервалами в 25 с (время отсчитывали фотографическим хронометром). Каждое третье опускание проволоки подкреплялось пищей (опускалась проволока с нанесенными бактериями). В контрольных экспериментах не было обнаружено изменений в количестве животных, что исключило возможное объяснение увеличения числа животных у проволоки за счет простого изменения двигательной активности инфузорий. Однако в этой работе не обнаружено воздействия обучения с подкреплением, что, по мнению автора, может быть объяснено слабой или замедленной реакцией животных на эффект опускания проволоки. Хотя такое объяснение автора и не лишено основания, более вероятной причиной нам кажется нарушение процедуры выработки условных рефлексов: слишком большие интервалы между сочетаниями (оптимальные условия частоты вообще специально не изучались) и непериодичность подкрепления (лишь третье предъявление иглы сочеталось с пищей). В дальнейшем Гельбером было показано, что выработанная реакция инфузорий — прилипание "дрессированных" животных к стерильной проволоке — сохраняется до 3 ч. Был продемонстрирован эффект, аналогичный угасанию, у трех групп культур P. aurelia, обученных по описанной методике, через 2 ч проверялось сохранение обучения. Группы перед началом проверки получили разное количество испытаний без подкрепления: группа 1 — 10 испытаний, группа 2 — 5, группа 3 — не получала совсем. Оказалось, что через 2 ч после дачи проб группа 3 показала количество ответов, превышающее остальные группы. Группы без обучения (контрольные) дали "нулевые" ответы. Эти опыты, без сомнения, подтверждают пластичность поведения инфузорий. Однако они, как и предыдущие эксперименты автора, не дают (в силу особенностей используемой методики) ответа на природу физиологических механизмов, лежащих в основе наблюдаемого явления. Отмеченное угасание на самом деле могло развиваться по механизму привыкания к отсутствию пищевого раздражителя, что исключает условнорефлекторную природу явления. В этой связи заслуживают внимания опыты Катца и Детерлайна (Katz, Deterline, 1958), которые использовали методику Гельбера с дополнительными контрольными экспериментами и показали, что после перемешивания жидкости в лунке с бактериями "дрессированные" бактерии не оседают на поверхности стерильной проволоки на дне лунки. В другом опыте авторы на дне лунки с голодными парамециями вносили с помощью платиновой проволоки небольшое количество бактерий. Повторив методику выработки условных рефлексов у инфузорий P. aurelia, они использовали только стерильную платиновую проволоку. Результаты достоверно опровергали опыты Гельбера: были получены такие же данные, как и в случае с проволокой, покрытой предварительно бактериями (инфузории без "обучения" собирались на поверхности стерильной проволоки). Таким образом, результаты опытов Гельбера, на наш взгляд, могут свидетельствовать лишь о способности инфузорий четко реагировать на пищевое раздражение или, возможно, на изменение среды, вызванное присутствием бактерий. Привыкание простейших. У организмов, лишенных нервной системы, могут быть более простые (неусловнорефлекторные) формы приспособительного поведения. Примером элементарного механизма накопления индивидуального опыта служит привыкание (Thorp, 1964). Под привыканием понимают прекращение реакции на постоянно действующий биологический раздражитель. Л. Г. Воронин (1968, 1969, 1972) относит привыкание простейших к несигнальной форме индивидуального приспособления. К одной из первых работ по выработке привыкания у простейших принадлежат опыты Данича (Danisch, 1921) на сувойках (Vortioella nebulivera). Автор исследовал привыкание к механическому раздражению у инфузорий. Критерием привыкания служило прекращение сокращения стебелька сувойки в ответ на раздражение. О динамике привыкания судили по количеству сокращений, необходимых для отсутствия сокращения. В работе получена зависимость скорости выработки привыкания от силы механического раздражения — привыкание вырабатывалось тем быстрее, чем меньше была сила используемого механического раздражителя. Позднее закономерности привыкания у простейших изучал Кинастовский. Им было подробно изучено влияние механического раздражения на сокращение инфузорий (Kinastowski, 1963). Автор в качестве механических раздражителей использовал свободно капающую каплю и специальный прибор, вызывающий вибрацию экспериментального сосуда. Оказалось, что оптимальными условиями для развития привыкания служит сила раздражителя от 400 до 1 600 эрг, частотой 10 и 15 раз/мин. Раздражение с большей силой (20 000 эрг) не приводит к уменьшению числа сокращений, а может вызывать судорожное состояние и летальный исход. Действие раздражителя оптимальной силы с частотой 1 раз в 1 мин не снижало числа сокращений во времени. Полное прекращение реакций в ответ на механическое раздражение в оптимальных условиях опыта наступало через 13 — 20 мин. В опытах Кинастовского животные с выработанным привыканием отвечали на сигнал другой модальности и на сигнал этой же модальности, но большей силы, что отличало полученный феномен от утомления. В аспекте рассматриваемых работ представляют интерес опыты А. Б. Когана (1963, 1964) на сувойках. Была показана способность этих инфузорий сокращаться в ответ на механическое раздражение — падение на предметный столик маленького свинцового шарика с высоты 3 см. Оказалось, что если к телу сувойки подвести стеклянную палочку так, чтобы выпрямление стебелька осуществлялось только на 3/4 его длины, то инфузории могут изменять свое поведение (замедление выпрямления стебелька, сокращение длины при выпрямлении) и, что особенно важно, сохранять эти изменения некоторое время после прекращения раздражения — удаления палочки. Автор опытов называет такую форму поведения своеобразной приспособительной реакцией. Отсутствие условного сигнала делает невозможным предположение об условнорефлекторном механизме формирования следовых реакций в этом случае. Можно предположить, что у сувоек вырабатывалось привыкание к модифицированному сокращению. Влияние различных факторов на выработку привыкания у инфузорий спиростом исследовал Аппельвайт (Applewhite, Morowitz, 1967; Applewhite, 1968; Applewhite, Stuart, 1969). В опытах Аппельвайта изучалось привыкание к вибрационному раздражению на популяциях из 60 — 100 инфузорий. Вибрационный раздражитель подавался каждые 4 с (падение груза с силой в 70 условных единиц). Критерием привыкания служило отсутствие сокращений на три раздражения подряд (использовалась фоторегисграция). Сохранение выработанного привыкания проверялось через 15 и 30 с после окончания опытов. В процессе опытов выявилась зависимость привыкания от температуры: при проверке сохранения привыкания через 15, 30, 60 и 90 с при температурах +15, +25 и +37° С оказалось, что более быстрое забывание (больший процент сократившихся инфузорий) наблюдается при температуре 37 С. Известно, что среда культивирования простейших может оказывать влияние на некоторые показатели функционального состояния животных (биение ресничек, изменение вязкости протоплазмы и т. п.). Поэтому особый интерес представляют эксперименты Аппельвайта и сотрудников (Applewhite et al., 1969) по изучению влияния на привыкание ионов металлов. Оказалось, что предварительное 15-минутное содержание инфузорий в растворе хлоридов калия и натрия (в концентрации к 0,5 до 0,005 моль/л), а также кальция, магния и марганца (в концентрации от 0,25 до 0,002 моль/л) не повлияло на выработку и сохранение привыкания, а воздействие ионов магния оказалось ярко выраженным. В этих опытах о динамике привыкания судили по первой, средней и последней парам раздражений, сохранение проверялось через 30 и 120 с после воздействия последнего раздражения. Двигательная активность "магниевых" инфузорий при этом не изменялась. Одним из предполагаемых механизмов влияния магния на привыкание авторы считают активирование эти ионом некоторых энзимов3. Интересные данные получены Аппельвайтом и соавторами (Applewhite et al., 1969) при изучении клеточной локализации феномена привыкания. После выработки привыкания животных разрезали пополам и вновь отдельно обучали переднюю и задние части. Оказалось, что для принятого критерия выработки (отсутствие реакции до трех нулей подряд) понадобилось примерно одинаковое число механических стимулов. Анализ полученных фактов позволяет согласиться с мнением авторов работы о том, что привыкание не является следствием утомления, повреждения или местной адаптации. Изучение роли макронуклеуса в процессе привыкания продемонстрировало отсутствие различий в динамике выработки этой реакции у половинок инфузорий, содержащих ядро и лишенных его, что позволило автору высказать предположение о локализации памяти клетки в цитоплазме. Оценивая серию работ Аппельвайта и сотрудников по изучению привыкания у ресничных инфузорий, необходимо отметить, что они вырабатывали нестойкие реакции (отсутствие реакции на три последовательных стимула), которые при оценке динамики выработки могут быть определены как первые признаки формирования привыкания, но не как прочно выработанная реакция. Основные закономерности выработки привыкания у Stentor coeruleus были изучены Вудом (Wood, 1970, 1972). У животных вырабатывалось привыкание к механическому раздражению. При постоянной частоте раздражения увеличение силы сигнала замедляло скорость выработки привыкания: в опытах с постоянной силой тока привыкание вырабатывалось тем медленнее, чем реже давался сигнал; при очень сильных раздражениях привыкание не вырабатывалось совсем. Следы привыкания в виде снижения частоты ответов на механические стимулы сохранялись в течение 3 ч. Несомненным достоинством опытов Вуда служит использование им теста на "растормаживание" как одного из существенных показателей наличия или отсутствия обучения в такой элементарной форме, каким является привыкание. Оказалось, что привыкание, выработанное на механический стимул с силой в 0,04 усл. ед., не растормаживается после однократного раздражения большей силы (0,12 усл. ед.) или после подпорогового раздражения электрическим током. По мнению автора, неспособность к растормаживанию служит единственным отличием свойств привыкания одноклеточных организмов от высших животных. Оригинальную форму обучения у стентора наблюдали Беннет и Френсис (Bennet, Francis, 1972). Критерием обучения служило сокращение времени передвижения стентора в капиллярной трубке, соединенной с относительно большим резервуаром, наполненным водой. При повторении проб с частотой один раз в 60 с время сокращалось от 58 с в первой пробе до 31 с в третьей. Реакция сохранялась примерно на одном уровне в течение 30 мин. Было показано также, что обучение происходит лишь при строгом соблюдении определенных условий опыта — вертикальном расположении капиллярных трубок с внутренним диаметром не более 0,5 мм. В трубках с диаметром 1 и 2 мм обучения не происходило. Авторы приходят к выводу о том, что изученный вид обучения есть не что иное, как результат привыкания стентора к механическому стимулу. Знакомство с экспериментами Беннета и Френсиса позволяет нам подобное объяснение считать справедливым. Итак, приведенный материал показывает, что одноклеточные организмы обладают свойством изменять поведение под влиянием различных раздражителей. Эти изменения сохраняются в течение 30 — 60 мин, а иногда до 3 ч. Сохранение следов от раздражений свидетельствует о способности простейших к накоплению индивидуального опыта. Наиболее детально вопрос о характерных особенностях функциональных механизмов, определяющих поведение простейших, был исследован в серии работ Н. А. Тушмаловой с сотрудниками (Тушмалова, 1968, 1974, 1977; Тушмалова, Доронин, 1971; Доронин, 1974; Тушмалова, Зазулина, 1977; Доронин, Тушмалова, 1978; Тушмалова, Кузьмичева, 1979). Приступая к изучению привыкания у донервных животных на примере привыкания Spirostomum ambigaum к вибрационному раздражению, авторы поставили своей целью прежде всего ответить на вопрос: в какой степени привыкание простейших соответствует понятию приобретенное поведение, выработанному в опытах на позвоночных животных и высших беспозвоночных. Были изучены такие свойства привыкания, как тренированность, растормаживание, информационная значимость интервалов между раздражениями, зависимость выработки этого феномена от исходного функционального состояния и возраста животных. По динамике реакции привыкания всех животных можно отнести к двум группам. В первой группе привыкание было относительным и заключалось лишь в уменьшении числа сокращений в единицу времени. Во второй группе животных реакция была абсолютной — в ответ на раздражение сокращения отсутствовали. Первое проявление привыкания отмечено после 1 — 10 мин; отсутствие сокращений до 10 нулей подряд — после 13 — 47 мин. Как показал анализ экспериментальных данных, оптимальными следует считать сигналы, действующие через промежутки времени, сравнимые с рефрактерными периодами4 (около 5 с), а по силе — с порогом сокращения инфузорий. Быстрее всего привыкание развивается при действии сигнала через 7 с. При увеличении интервала между раздражениями увеличивается время, необходимое для достижения 100 %-ного угашения реакции. Однако реакция вырабатывалась и при сигнале, действующем значительно реже — через 1 мин: в этих случаях феномен развивался значительно медленнее. Привыкание вырабатывалось и при раздражении 1—9 раз в минуту. Однако для достижения критерия 10 нулей требовалось значительно больше времени (до 50 мин). Наиболее четко значение временного интервала в динамике выработки привыкания было продемонстрировано в опытах с применением раздражителей в аритмическом режиме, по случайному закону, отмечалось замедление в выработке привыкания при аритмическом режиме раздражения. Как и у более высокоорганизованных животных, у инфузорий были выявлены индивидуальные отличия в динамике привыкания (рис. 1). У животных с выработанным привыканием в 100 % сохранялась реакция на сигнал другой модальности: и более сильный сигнал этой же модальности, что отличает изучаемый феномен от утомления. Условия, при которых наблюдалось утомление инфузорий, резко отличались от условий выработки привыкания. Нереагирование на сигнал любой силы и частоты — утомление — развивалось в результате длительной стимуляции инфузорий до 3 — 4 ч с частотой раздражения 1 раз в 1 с и силой, превышающей пороговую в 4—5 раз. При изучении ультраструктурных изменений инфузорий в процессе привыкания также обнаружены разнонаправленные сдвиги при привыкании и утомлении в ядрышке макронуклеуса. Так, привыкание сопровождалось активацией ядрышка — увеличением синтеза рибосомной РНК. При утомлении наблюдалось резкое снижение синтеза рибосомной РНК. Анализ результатов показал, что привыкание к вибрационному раздражителю может быть отнесено к категории явлений, характеризующих следовые процессы, т.е. к памяти. Неясным оставался вопрос о том, в какой степени следовые явления одноклеточных обладают свойствами, общими с другими, более высокоорганизованными животными. Одним из критериев приобретенности (выработанности) реакции служит способность к растормаживанию5. Использование приема растормаживания привыкания показало, что эта реакция никогда не восстанавливается при действии экстренных раздражителей более слабых, чем агент, на который было выработано привыкание. При действии же раздражителей большей силы, чем индифферентный, растормаживание носило случайный характер. Воздействия стимулами другой модальности (электрический ток, культуральная среда, насыщенная кислородом или углекислотой) не дали положительных результатов. Эффект растормаживания привыкания был отмечен лишь при действии очень сильного раздражителя — пищевого, т. е. при внесении в камеру с инфузориями капли бактериальной суспензии. Для выяснения вопроса, какой же из факторов (рН среды или непосредственно бактерии) является действующим, в контрольных экспериментах в камеру вносили каплю прокипяченной суспензии либо фильтрат суспензии бактерий, либо чистый подщелоченный культуральный раствор. Статистически достоверное растормаживание наблюдалось только при использовании капли культуральной среды повышенной щелочности (рН > 8). Помимо растормаживания, важным критерием способности животных формировать следовые реакции: является ускорение обучения при неоднократном повторении процедуры обучения — тренированность. У популяции инфузорий привыкание вырабатывалось в течение 20 мин, затем инфузории отсаживались в часовое стекло с культуральной жидкостью. Через 60 мин клетки вновь подвергались воздействию вибрационного раздражители Процедура повторялась 5 — 6 раз. Анализ полученных данных показал, что при первом применении раздражителей (первый опыт) уровень ответов снижается до 35—40 %. По мере повторения опытов достоверно уменьшается вероятность сокращения только на первые раздражители. При 5-й и 6-й повторностях инфузории реагируют на механический стимул с вероятностью, которая не отличается (Р > = 0,9) от уровня ответов в конце первого опыта. Таким образом, процесс тренированности привыкания у инфузорий выражается только в понижении вероятности сокращения на первые стимулы по мере применения серий раздражителей (при интервале между опытами 60 мин).
 Рис.
1. Динамика выработки привыкания у
отдельных инфузорий (по: Тушмалова,
1972): по оси ординат — время, мин; по оси
абсцисс — номера инфузорий; светлые
столбцы диаграмм — время, необходимое
для достижения 1 нуля, с сетчатой
штриховкой — 3, с косой штриховкой — 5,
черные — 10 нулей
Рис.
1. Динамика выработки привыкания у
отдельных инфузорий (по: Тушмалова,
1972): по оси ординат — время, мин; по оси
абсцисс — номера инфузорий; светлые
столбцы диаграмм — время, необходимое
для достижения 1 нуля, с сетчатой
штриховкой — 3, с косой штриховкой — 5,
черные — 10 нулей
Не отмечено прямой зависимости динамики привыкания от уровня спонтанной двигательной активности и пищевой возбудимости: скорость выработки не зависела от числа пищеварительных вакуолей. Однако при длительном голодании привыкание не вырабатывалось. Обнаружена зависимость привыкания от возраста инфузорий и состояния их ядерного аппарата. Оптимальным для формирования привыкания оказался возраст 45 — 55 ч после деления (примерно 50 % продолжительности жизни). В возрасте 105 ч, когда макронуклеус инфузорий находится в состоянии конденсации (физиологическая реорганизация, по И. Б. Райкову, 1967), привыкание не вырабатывалось. Говоря о привыкании простейших, важно подчеркнуть, что этот феномен проявляется не только у ресничных инфузорий, но и у более простых одноклеточных — Amoeba proteus. Н. А. Тушмалова и И. Л. Кузьмичева (1978, 1979) изучали закономерности формирования привыкания амеб на периодическое предъявление света 800 — 2 000 лк с интервалом в 1 — 2 мин. Критерием выработки привыкания служило прекращение течения цитоплазмы в ответ на 10 предъявлений света подряд. Как и у ресничных инфузорий, у амеб было выявлено абсолютное привыкание (52,5%) и относительное — уменьшение числа ответных реакций (47,5%). Оказалось, что по зависимости скорости выработки привыкания от силы и частоты раздражения этот феномен у амеб сопоставим с таковым у спиростом. Таким образом, проведенные опыты показали, что приобретенное поведение простейших обладает свойствами, общими с таковыми у животных, имеющих нервную систему, и одновременно несет черты более примитивной организации.
Поведение кишечнополостных
Об условнорефлекторной деятельности кишечнополостных. Вопрос о способности кишечнополостных формировать условные рефлексы в настоящее время остается открытым. Результаты немногочисленных экспериментальных попыток выработки условных реакций у различных видов этого типа до сих пор не дали положительных результатов. Одна из первых работ была предпринята А. А. Зубковым и Г. Г. Поликарповым (1951). Они проводили опыты в часовом стекле и обнаружили, что движения гидры, прикрепленной близко к поверхности воды, совершаются в направлении от поверхности воды, хотя на глубине в условиях равномерного освещения наблюдаются движения во всех направлениях; через 60 мин после начала опыта движения в сторону поверхности воды прекращаются. При вторичном повышении уровня воды "выработанная" реакция сохраняется до 3—4 ч, а затем угасает. На основании таких свойств реакции, как угашение при неподкреплении, торможение при действии экстрараздражителей (сотрясение и изменение освещенности), авторы определили наблюдаемую модификацию поведения гидр как выработку условного рефлекса, однако результаты проведенных опытов могут иметь и иные объяснения. Так, Л. М. Чайлахян (1957) объясняет "приобретенность" такой реакции гидр лишь изменением состояния мышечных элементов. Более детальный анализ условнорефлекторной деятельности кишечнополостных дан в работе Л. М. Чайлахяна (1957) на примере опытов с пресноводным полипом. Сочетание света (условный сигнал длительностью от 5 с до 1 мин) с электрическим током — безусловный раздражитель при интервалах от 2 до 3 мин привело к проявлению реакции сокращения животных только на изолированное действие света. При увеличении интервала между условными и безусловными раздражителями до 5—6 мин "условная" реакция отсутствовала. Выявленные закономерности позволили автору заключить, что реакция полипа на свет не условный, а суммационный рефлекс — результат повышения возбудимости животного вследствие суммации следов от предыдущих раздражений электрическим током, под влиянием чего допороговая интенсивность света становилась пороговой. Нами была проведена серия экспериментов по изучению способности к формированию условнорефлекторной памяти у пресноводной гидры (Hydra attenuate). В первых двух сериях опытов (Тушмалова, Симирский, 1977) были исследованы реакции гидр на действие сочетанных раздражителей при попытке выработать пищевые и электрооборонительные условные рефлексы. Условным сигналом служил свет лампы освещенностью в 1000 лк с изолированным действием в течение 30—60 с, на протяжении 30 с совпадающий с безусловным пищевым подкреплением (циклопы). Интервалы между сочетаниями определялись временем заглатывания циклопа и составляли 6 — 10 мин. Критерием положительной условной реакции было выбрано открывание гипостома в ответ на изолированное действие света. Контролем служили животные, которые ежедневно получали дозированную пищу в отсутствии света, и животные, получающие пишу при непрерывном освещении красным светом. Изменение реакции гидр от сочетания к сочетанию выражалось лишь в том, что свет неспецифически влиял на пищевое заглатывание гидр, изменяя как время от захвата пиши до заглатывания, так и время заглатывания пищи. Ни в одном из опытов условной пищевой реакции в ответ на изолированное действие света не наблюдалось. Во второй серии опытов условным сигналом служила вибрация частотой 60 Гц с длительностью импульса 3—6 с, безусловным — постоянный электрический ток (15 В), подаваемый в виде импульсов длительностью 0,5—3 с. Критерием выработки условного рефлекса была реакция сокращения тела при допороговой вибрации. Для исключения суммации разномодальных раздражителей интервал между сочетаниями был увеличен до 3 мин. Увеличение числа сочетаний до 100 — 200 не привело к снижению порога вибрации. В третьей серии опытов (Тушмалова, Устинова, 1978, 1979) была изучена возможность выработки химического оборонительного условного рефлекса на свет у гидр. Опыты проводились с животными в возрасте 8 — 10 дней, считая его началом отделение гидры от материнской особи. Условным раздражителем служил свет (1000 лк), безусловным — 0,2 %-ный раствор хлорида натрия (эта концентрация вызывает сокращение всего тела гидры). Условный раздражитель действовал изолированно 30 с, а 30 с его действие совпало с безусловным. Раздражители предъявлялись с интервалом 4 мин, что по данным Л. М. Чайлахяна исключало суммационный эффект. Ежедневно каждая гидра получила 20 сочетаний условного и безусловного раздражителей. В процессе опытов у 100 % гидр наблюдалось уменьшение латентного периода реакции сокращения тела гидры в ответ на безусловный раздражитель с достоверностью Р < 0,001. Однако число условных ответов к 6-му дню опытов увеличилось только до 18,6 %. Следовательно, и в этой серии экспериментов выработка условных рефлексов также не наблюдалась, а была лишь отмечена модификация поведения в виде уменьшения латентного периода реакции на безусловный раздражитель. Таким образом, результаты опытов, проведенных с различными методическими приемами, оказались аналогичными — они не выявили свойств приобретенных реакций, которые можно было бы определить как условнорефлекторные. Особенности привыкания кишечнополостных. Исследование закономерностей формирования привыкания кишечнополостных началось относительно недавно. Так, Рушфор (Rushforth, 1963) изучал привыкание гидр к сфокусированному пучку света. Скорость привыкания к свету, подаваемому с частотой 1 — 5 раз в 1 мин, зависела от силы освещенности: в то время как для первого отсутствия ответа при сильном свете понадобилось 40 сокращений, при слабом — всего 3 — 5. В другой группе экспериментов изучалось привыкание к механическим раздражителям: один характеризовался частотой 105 Гц длительностью 2 с и подавался с интервалом в 16 с, другой характеризовался частотой 50 Гц, длительностью 1 с и подавался с интервалом в 19 с. Привыкание к механическому стимулу вырабатывалось тем быстрее, чем чаще подавался сигнал: при частоте 105 Гц — за 8 ч, при частоте 50 Гц — за 20 ч. В этих опытах гидры, "привыкшие" к механическому раздражению, отвечали сокращением на действие света. После удаления щупалец у гидр исчезали сокращения на механические раздражения, но сохранялись реакции на свет. Одновременное же удаление щупалец и гипостома (прикрепительное приспособление в области ротового отверстия) прекращало сокращения на оба названные раздражения. Принципиальная важность этих опытов заключается прежде всего в том, что они доказали существование у гидр различных рецепторов для восприятия света и механического раздражения. Полученные данные согласуются с выводами Лентса (Lentz, 1966) о том, что у гидр рецепторы, воспринимающие свет, локализованы в гипостоме, а воспринимающие механические раздражения — в щупальцах. Привыкание у гидр может вырабатываться и на химические раздражения (Rushforth, 1965, 1967). На основании анализа экспериментальных данных Рушфорт (Rushforth, 1967) дает следующие критерии привыкания гидр: 1) достаточно частое предъявление стимулов приводит к уменьшению ответа вплоть до прекращения; 2) при прекращении раздражения ответные реакции восстанавливаются; 3) более быстрое развитие привыкания наблюдается при меньшем межстимульном интервале; 4) привыкание развивается тем быстрее, чем слабее стимул; 5) при выработке привыкания к одному из раздражителей сохраняется реакция на сигнал другой модальности. Результаты привыкания у гидр показали необходимость дальнейшей работы в этом направлении, так как до сих пор отдельные свойства привыкания остались неизученными. В частности, не проводились тесты на растормаживание, недостаточно изучено такое свойство этой реакции, как тренированность. Изучение способности к тренированности затруднено еще и тем, что она может проявляться не только в ускорении выработки реакции, а например, в изменении соотношения отдельных фаз при выработке реакции. Выяснению этих вопросов и были посвящены эксперименты Н. А. Тушмаловой и сотрудников на гидре (Hydra attenuate) (Тушмалова и др., 1975; Бресткина и др., 1978). Критерием выработки привыкания служило прекращение сокращения щупалец при действии пороговых вибрационных импульсов частотой 60 Гц, следующих через каждые 10, 15 и 30 с на протяжении 30 мин. Как и у инфузорий, угашение реакции сокращения щупалец зависело от частоты раздражения: при более частом оно было быстрее. Приобретенная реакция сохранялась на протяжении 15, 60 мин после прекращения раздражения. "Привыкшая" гидра сохраняла реакцию сокращения щупалец в ответ на действие раздражителей большей силы или иной модальности. Как и в опытах с инфузориями, выработка привыкания у гидр зависела от исходного функционального состояния животных. Так, привыкание наиболее быстро вырабатывалось при уровне пищевой возбудимости, наблюдаемой через 4 — 5 дней после кормления животных. Скорость выработки привыкания зависела от возраста гидр — привыкание вырабатывалось быстрее у однодневных гидр по сравнению с 15-дневными (рис. 2).
 Рис.
2. Динамика выработки привыкания у гидр
разного возраста (по: Бресткина и др.,
1978) по оси ординат — число ответных
реакций %; по оси абсцисс — возраст, сут
Рис.
2. Динамика выработки привыкания у гидр
разного возраста (по: Бресткина и др.,
1978) по оси ординат — число ответных
реакций %; по оси абсцисс — возраст, сут
Воздействие сильными (намного превышающими пороговый) раздражителями на "привыкшую" гидру не изменяет дальнейшего протекания процесса привыкания. Этот факт может свидетельствовать в пользу того, что, по-видимому, для гидры, так же как и для простейших, нехарактерно явление растормаживания в его типичном проявлении. Гидр повторно через различные промежутки времени подвергали действию механических (вибрационных) раздражителей в режиме привыкания на протяжении 30 мин. В результате выяснилось, что при интервалах, сравнимых со временем сохранения следа после однократного привыкания, наблюдается ярко выраженное ускорение привыкания от опыта к опыту. Однако при интервалах между опытами в 24 ч декремента не наблюдалось. Этот результат представляет интерес, поскольку именно по степени выраженности тренированности у животных различного уровня филогенеза (или нейронных систем различной сложности) при различных интервалах между приложениями можно судить о степени консолидации следа и, возможно, определить четкий критерий различия памяти кратковременной и долговременной — основных функциональных механизмов поведения. Таким образом прогресс в развитии функциональных механизмов поведения у кишечнополостных по сравнению с простейшими заключается в появлении нового свойства привыкания — тренированности.
Поведение червей
Характеристика условнорефлекторной деятельности плоских червей (планарий). Прогресс в развитии нервной системы на уровне низших (ресничных) червей, или турбеллярий, по сравнению с ранее описанными группами животных выражен началом процесса цефализации. Интегрирующее значение нервной системы на этой стадии филогенеза выражается в регулировании церебральным ганглием важных функций организма, например, координации различных двигательных реакций (Bullock, Horridge, 1965). Вопрос о выработке истинных классических условных рефлексов у низших (ресничных) червей, представителями которых являются планарии, долгое время считался дискуссионным. Впервые схема периодического сочетания условного раздражителя с безусловным была использована Мак-Коннелом и соавторами (Tompson. McConnel, 1955; McConnel et al., 1959, I960). Эксперименты проводились в специальной камере, снабженной латунными электродами. Действие условного сигнала (свет) на 2 с предшествовало безусловному электрическому раздражению. В течение одного опыта животные получали 150 сочетаний света с электрическим током через каждые 20 с. Критерием выработки условных рефлексов служило сокращение животных в ответ на свет. По окончании серии сочетаний свет — ток частота ответов на свет в виде сокращений и поворотов постоянно повышалась. В контрольных экспериментах, где животные получали избирательно свет или электрический ток (или не подвергались воздействию раздражителей), отмечено лишь небольшое изменение числа ответов. В результате статистической обработки результатов исследования авторы пришли к выводу о возможности выработки классических условных рефлексов у планарий. Наварра (Navarra, 1961) на планарии Dugesia tigrina в течение одного опыта 300 раз сочетал электрическое раздражение со светом и получил аналогичные данные. Ли (Li, 1963), работая с планарией Cura forlmani, в условиях свободного передвижения в качестве подкрепления применял выключение верхнего света при пересечении животными узкого пучка света, направленного параллельно дну экспериментального цилиндра (условный сигнал). У экспериментальных животных по сравнению с контрольными наблюдался большой процент положительных реакций. Вест (Best, 1962) в опытах с планарией Dugesia dorotocephala и Cura forlmani в сложном лабиринте при использовании в качестве условного сигнала света, а безусловного — лишение планарии воды подтвердил способность планарии к дрессировке. В работе Гриффард (Griffard, 1963) с Phagoata gracilis условным раздражителем служил ток воды, безусловным — пропускание электрического тока. На основании проведенных опытов автор также приходит к положительному заключению о выработке условных рефлексов у планарии. Необходимо отметить, что Гриффард впервые проводил эксперименты более длительное время, чем предыдущие авторы, проверяя сохранение условной реакции через 10 — 15 ч. Используя методику Томпсона и Мак-Коннела (Tompson, McConnel, 1955), Джекобсон и соавторы (Jacobson et al., 1967) также пришли к выводу о возможности выработки у планарий классических условных рефлексов. Эксперименты Томпсона точно воспроизвёл Хэлас с сотрудниками (Halas et al., 1962). Однако эти авторы рассматривают реакции планарии на условный сигнал (свет) как рефлекс сенсибилизации. А. Н. Черкашин и соавторы (1966) изучали реакции планарии на сочетание монофазного электрического тока (безусловный раздражитель) и света (условный раздражитель). Авторы делают заключение о том, что в данных методических условиях приобретенные реакции не достигают уровня условных рефлексов. Таким образом, вопрос о физиологических механизмах приобретенных реакций у планарий оставался открытым. Для окончательного ответа на него были проведены дальнейшие эксперименты на 5 видах планарий с различной экологией, включая эндемиков озера Байкал. В первой серии вырабатывался электрооборонительный условный рефлекс на свет (Воронин, Тушмалова, 1965). Оказалось, что для первого проявления реакции, сходной с условнорефлекторной, понадобилось от 51 до 76 сочетаний. Однако упрочения ее не происходило, несмотря на 300 сочетаний раздражителей. Во второй серии опытов в качестве условного раздражителя применялся слабый электрический ток, вызывавший ориентировочный рефлекс в виде поворота головного конца тела животного, и в качестве безусловного — отрицательная реакция планарий на свет (уплывание в сторону неосвещенной части желоба). Последовательного закрепления реакции из опыта в опыт, так характерного для классических условных рефлексов, не наблюдалось и в этом случае. Временная связь у всех планарий была нестойкой, количество положительных реакций из каждых 10 сочетаний условного сигнала с безусловным редко превышало 50%. Была использована и более простая схема экспериментов, в которой условный сигнал был постоянным. Дно экспериментальной камеры было разделено на темную и светлую половины. В опыте сначала определялось время нахождения животных на темном и светлом полях, затем на одном из полей они получали удар электрическим током, вызывающим продольное сокращение тела. Основным критерием выработки условной реакции в этой серии экспериментов служило уменьшение времени нахождения планарий в той части камеры, где возникало электрическое раздражение. Закономерного и стойкого увеличения времени пребывания животных на поле, где животным не наносилось безусловное раздражение, не наблюдалось. В дальнейшем был применен более адекватный безусловный раздражитель — раствор поваренной соли (Тушмалова, Громыко, 1968). В этом случае максимальный уровень положительных реакций не превышал 50 — 70%, и, как при электрическом подкреплении, они характеризовались нестойкостью от опыта к опыту и в течение одного эксперимента. Однако при этом возможно было четко отдифференцировать начало выработки условных рефлексов (после 40 — 60 сочетаний), максимальный уровень положительных реакций (120 — 200 сочетаний) и снижение уровня условнорефлекторной деятельности (сокращение числа положительных реакций от 10 до 0 %) после 200 — 300 сочетаний. У контрольных животных при изолированном применении света в 100% случаев наблюдалось лишь слабое возбуждение, выражающееся в сокращении (не более 8 — 10% по отношению к положительным реакциям опытных животных). Аналогичная картина наблюдалась в опытах с псевдообучением (предъявление условного и безусловного раздражителей в случайном порядке). При остром угашении реакции самопроизвольно не восстанавливались. Из анализа экспериментального материала следует, что условные рефлексы у изученных пресноводных планарий недостаточно стойки, не обладают всеми качествами классических условных рефлексов. Они характеризуются следующими признаками, общими для экологически различных видов животных: непрочностью в течение одного опыта, непрочностью от опыта к опыту (упрочение не наступало даже после 335 сочетаний условного сигнала с безусловным), угасанием реакций после 200 — 300 сочетаний, несмотря на подкрепление. Перечисленные свойства условных рефлексов не являются отражением индивидуальных особенностей отдельных видов, так как они характерны для животных с различной экологией (от живущих в пойменных ручьях Подмосковья до эндемиков Байкала) (Тушмалова, 1968, 1967). Отмеченные закономерности не являются и следствием примененных методических приемов, аналогичные данные были получены во всех 6 сериях опытов, значительно отличающихся методически. Таким образом, подобные реакции можно отнести к категории лишь примитивных нестойких условных рефлексов, свойственных животным определенного уровня филогенетического развития (Воронин и др., 1972; Тушмалова, 1976). Особенности строения головного ганглия планарий. Одна из замечательных филогенетических особенностей ресничных червей, к которым относятся планарии, заключается в том, что они представляют уровень "перехода" диффузной нервной сети в концентрированную систему. У ресничных червей впервые в эволюции нервные элементы концентрируются на переднем конце тела (Hanstrom, 1928; Заварзин, 1941; Беклемишев, 1964), т. е. появляются зачатки цефализации. Детальные сведения о клеточном (цитоархитектоническом) строении этого примитивного мозга в литературе долгое время отсутствовали, и лишь стремление определить степень соответствия примитивного поведения планарий структурной организацией их "мозга" послужило поводом к конкретным исследованиям. Строение мозга планарий было изучено на примере типичного представителя ресничных червей P. nigra (Бочарова, Тушмалова, 1968; Свешников и др., 1968; Тушмалова, Свешников, 1972). Церебральный ганглий P. nigra расположен приблизительно в 500 мкм от головного конца тела и состоит из двух частей. Из-за отсутствия оболочки его границы могут быть определены относительно. Характерной особенностью головного ганглия планарий служит наличие в нем многочисленных специфических образований, состоящих из клеток, которые располагаются вокруг проходящих через мозг дорсовентральных мышц (инсулярные, или островковые включения по: Bohmig. 1906), в количестве от 40 до 70. В состав ганглия входят разнообразные по величине и форме клетки (большинство биполярные, реже мультиполярные). Особенностью большинства клеток является бедность цитоплазмой и отсутствие ядрышек у большинства клеток. Размеры клеток варьируют от 3 — 5, 10 — 15 до 15 — 20 ("гигантские" клетки) мкм. Последние отличаются богатой цитоплазмой, большим ядрышком и высоким содержанием РНК в цитоплазме. Наличие в цитоплазме этих клеток характерной зернистой субстанции дало право считать их наиболее дифференцированными нейронами всего мозга. Таким образом, исследования показали, что церебральный ганглий планарий несет черты примитивной организации, состоит в основном из мелких, малодифференцированных клеток. Основной особенностью гистологического строения ганглия служит наличие в нем большого количества мышечных и паренхиматозных клеток, выполняющих, по-видимому, нейросекреторные функции. Только отдельные ("гигантские") клетки могут быть с большей степенью уверенности классифицированы как нервные. Представление о примитивности нервной системы планарий разделяют многие авторы (Кричинская, 1972; Лурье, 1975). Они считают, что все нервные клетки планарий бифункциональны и обладают способностью к нейросекреции. Таким образом, появившаяся впервые в эволюции у планарий цефализация представлена здесь в своей примитивной форме и к ней приурочена приобретаемая в индивидуальной жизни организма примитивная форма нестойких условных рефлексов. Свойства условнорефлекторной деятельности кольчатых червей. Среди большой группы кольчатых червей, являющихся эволюционными потомками плоских червей, особое место занимают представители класса олигохет — дождевые черви, на которых проводились основные опыты по условным рефлексам. Наиболее подробно способность к формированию условных рефлексов у дождевых червей Lumbricus rubellus изучена Н. М. Хоничевой (1968). Она вырабатывала оборонительные условные реакции в Т-образном лабиринте. Черви обучались поворачивать в правый или левый рукав лабиринта. Безусловным раздражителем служил переменный ток различной интенсивности, а условным — сам лабиринт, элементы которого, вероятно, воспринимались проприоцептивной и тактильной афферентациями. Критерием выработки рефлекса служило увеличение числа по воротов в рукав лабиринта, где животные не подвергались электрической стимуляции. Максимальное число сочетаний доходило до 400. Однако постепенного, закономерного увеличения числа положительных реакций отмечено не было. Характерной особенностью обучения червей явилось колебание процента положительных реакций. Сравнение динамики обучения трех групп червей показало, что повышение силы безусловного раздражителя (электрического тока) увеличивает скорость выработки реакций. Так, при напряжении 5 В максимальный процент положительных реакций был отмечен на 20-й опытный день, при 12 В — на 17-й опытны день, при 18 — уже на 11-й день. Оказалось, что эта тенденция к более быстрому обучению осуществляется за счет сокращения продолжительности его 1-й стадии, которая равнялась 12, 8 и 5 дням соответственно для слабого, среднего и сильного подкрепляющего раздражителя. Наблюдалась зависимость максимального процента положительных реакций от величины подкрепляющего раздражителя: при 5 В — 54, при 12 В — 70, при 18 В — 85%. Существенно, что удаление надглоточного ганглия не изменяло динамику обучения — факт, несомненно свидетельствующий в пользу незначительной интегрирующей роли данного отдела мозга. Совсем иные закономерности были получены в опытах на полихетах. Так, Копеланд (Gopeland, 1930) у Nereis virens выработал устойчивый условный рефлекс на тактильное раздражение. У Nereis pelagica (беломорская форма) была четко продемонстрирована возможность выработки пищевых условных рефлексов на свет и вибрацию (Воронин и др., 1982). Критерием выработки условных рефлексов служило изменение "знака" фотонегативной реакции. До начала опытов черви постоянно находились внутри стеклянной трубки, а после 30 — 50 сочетаний условного раздражителя с пищей животные проявляли четкую поисковую пищевую реакцию, выходя из трубки. Анализ результатов показал, что у полихет вырабатываются реакции, обладающие всеми основными свойствами истинных условных рефлексов: возрастание числа положительных ответов от опыта к опыту, высокий максимальный процент положительных реакций (до 80 — 100) и длительность их сохранения (до 6 — 15 дней). Весьма существенно, что выработанная реакция угасала при отсутствии подкрепления и самопроизвольно восстанавливалась. В контрольных опытах — при псевдообуславливании — увеличения числа положительных ответов не наблюдалось. Выявленные закономерности условнорефлекторной деятельности полихет коррелируют с относительно дифференцированным мозгом животных. Хорошо известно, что одной из особенностей их мозга является возникновение специального ассоциативного центра — грибовидных тел. Удаление этих отделов мозга приводит к нарушению условных рефлексов, как показано в опытах на пчелах (Воскресенская, 1957). Таким образом, истинные условные рефлексы как один из достаточных совершенных механизмов, определяющих приобретенное поведение, впервые в эволюции, по-видимому, появляются у полихет (Тушмалова, 1980).
К проблеме условнорефлекторной деятельности иглокожих
По сравнению с другими группами беспозвоночных животных поведение иглокожих менее изучено. К отдельным попыткам выработать условные рефлексы у этой группы беспозвоночных относится работа Дибшлага на трех видах морских звезд (Asterias rubens, Astropecten bispinonus, A. spinolosus) (Diebschlag, 1938). Условные рефлексы, выработанные при сочетании света или тактильного условного сигнала с оборонительным подкреплением, сохранялись всего 15 мин. Такую реакцию нельзя отнести к категории истинного условного рефлекса. Позже В. И. Сафьянц (1958), работая на морских ежах, показал, что сочетание понижения уровня воды с последующим осушением приводит к опусканию ежей ниже уровня снижения воды. Такую модификацию поведения, свидетельствующую о накоплении индивидуального опыта, можно объяснить на основе механизма привыкания — неассоциативного обучения. Наиболее детально вопрос об условнорефлекторной деятельности иглокожих был изучен В. А. Соколовым (1960, 1966) в опытах на морской звезде (Asterias rubens). Ему удалось показать, что у морских звезд вырабатываются двигательные условные рефлексы на свет и тактильный раздражитель. Важно подчеркнуть, что в этих опытах были продемонстрированы такие важные критерии условных рефлексов, как выработка угашения и дифференцировки.
Поведение членистоногих
Характерной эволюционной особенностью представителей данной систематической группы служит высокая степень цефализации. Другой отличительной чертой является достаточно четко выраженная структурно-функциональная специализация мозга. Названные особенности наложили отпечаток на свойства функциональных механизмов, определяющих приобретенное поведение, закономерности которого изучаются на чисто поведенческом и нейрофизиологическом уровнях. Сказанное относится прежде всего к насекомым. Высшая нервная деятельность наиболее детально изучалась у ракообразных и насекомых. Условные рефлексы ракообразных. Беспозвоночные этой группы резко отличаются по морфологической организации нервной системы: у низших раков она имеет признаки кольчатых червей, что отражается и в функциональных механизмах поведения. Положительные результаты выработки условного рефлекса у дафний были получены лишь в своеобразных опытах А. Б. Когана и А. П. Семенова (1955). У дафний был выработан пищевой условный рефлекс по схеме: кормление в темноте (ночь), голодание на свету (день). На высших ракообразных были получены результаты при использовании обычной схемы выработки условных реакций. В опытах на раках-отшельниках Сполдинг (Spaulding, 1904) выработал относительно стойкий (сохранялся 7 дней) пищевой условный рефлекс на затемнение. Возможность выработки угасательного и дифференцировочного торможения у рака-отшельника (Pagurus striatus) была продемонстрирована Михайловым (Mikhailoff, 1922) еще в 1922 году. Положительные результаты были получены в опытах А. Я. Карась (1964) на ресничных раках Astacus leptodactilis по выработке двигательного пищевого условного рефлекса на свет. Наиболее правдоподобно закономерности высшей нервной деятельности у высших раков были исследованы в серии работ Л. Я. Карась (1962, 1963, 1964) на крабах Carcinus means. Была показана возможность выработки пищевых, зрительных, тактильных условных рефлексов и условных рефлексов на раздражение статорецепторов и выработки основных видов внутреннего торможения (угасательного, дифференцировочного и условного тормоза). Особенностью положительных и отрицательных условных связей была их нестойкость. Весьма существенно, что у животных этого уровня филогенеза формировались условные рефлексы на цепные и комплексные раздражители. Однако в отличие от насекомых (пчел) эти реакции также отличались нестойкостью. Автор предполагает, что сигнальным признаком цепных раздражителей была сила отдельного компонента, а не их определенная последовательность. Такое представление свидетельствует о низком уровне аналитико-синтетических свойств нервной системы ракообразных. Морфологической основой сформулированного представления может быть существование связей в ЦНС этих животных (Коштоянц, 1957).
Заключение
Анализ работ, посвященных изучению приобретенных форм поведения у беспозвоночных, позволяет сделать заключение о том, что способность к накоплению индивидуального опыта появляется в эволюции на самых ранних ступенях развития — у донервных простейших организмов (Тушмалова, 1976, 1981). Однако функциональные механизмы, определяющие поведение беспозвоночных животных разного уровня филогенеза, различны (Тушмалова, 1980). Так, из двух форм эволюции ВНД (Воронин, 1977) сигнальная форма индивидуального приспособления появляется лишь у полихет, а низшим червям, кишечнополостным и простейшим свойственна несигнальная форма приспособления. Для рассмотрения основных закономерностей эволюции поведения беспозвоночных принципиальное значение имеет факт формирования приобретенного поведения у донервных организмов за счет макромолекулярных и ультраструктурных перестроек (Тушмалова и др., 1974; Зазулина, 1979). В эволюционном плане весьма существенно, что характер этих перестроек однозначен у животных разного уровня филогенеза, обладающих различными функциональными механизмами приобретенного поведения (Тушмалова, 1980, 1983). Исходя из представлений Л. А. Орбели (1961) о том, что в процессе эволюционного развития старые функциональные отношения не уничтожаются, а лишь затормаживаются новыми, можно высказать предположение о сохранении протоплазмой клеток организмов, обладающих нервной системой, свойства простейших — способности формировать следовые реакции за счет внутриклеточных перестроек. Развиваемое нами представление согласуется с мыслью П. К. Анохина (1968), считающего, что эволюция изменила лишь "средства доставки информации", но ее конечная обработка осуществляется одним и тем же способом — молекулярным. Следовательно, можно предположить, что усложнение (совершенствование) функциональных механизмов поведения по мере возрастания эволюционного уровня определяется структурными изменениями нервной системы. Этот вывод на примере беспозвоночных иллюстрирует одно из существенных методологических положений нейрофизиологии о зависимости "качественно нового ранга рефлекса — условного рефлекса" от уровня развития нервной систем (Карамян, 1969). Действительно, при сравнении функциональных механизмов поведения беспозвоночных обращает на себя внимание параллель в степени сложности структурной функциональной организации. Это есть не что иное, как выражение морфофизиологического прогресса — ароморфоза по А.С. Северцову (1967) (см. табл. 1).
Таблица 1 Соотношение морфологических и функциональных ароморфозов на примере эволюции поведения беспозвоночных
Группы животных |
Ароморфозы |
|
|
Морфологический уровень (организация нервной системы) |
Функциональный уровень (высшая форма физиологических механизмов поведения) |
Простейшие |
Донервный уровень |
Привыкание |
Кишечнополостные |
Примитивная нервная система |
Привыкание + тренированность |
Плоские черви |
Зачатки цефализации |
Нестойкие условные рефлексы |
Кольчатые черви |
Ассоциативный центр мозга — грибовидные тела |
Истинные условные рефлексы |
Насекомые |
Структурно-функциональльная дифференцировка мозга |
Продолжительные и отрицательные условные рефлексы, инсnрументальные условные рефлексы, цепные условные рефлексы. |
Таким образом, основные закономерности функциональных механизмов поведения беспозвоночных являются по существу филогенетическим отражением принципа единства структуры и функции.
Фриш к. Как пчелы разговариваривают друг с другом
Печатается по изд.: Фриш К. Из жизни пчел / Пер. с нем. Т.И. Губиной. М.: Мир, 1966. С. 122-170.
В предыдущих главах говорилось о дрессировке, которая позволяет получить некоторые данные о чувствах пчел. Предпосылкой для таких опытов служит то, что пчелы, которых мы хотим дрессировать, прилетают к нашему опытному столику. Чтобы привлечь пчел к опытному столику, положим на него несколько листов бумаги, обильно смазанных медом. В большинстве случаев проходит несколько часов, а нередко и несколько дней, пока одна из снующих повсюду в поисках корма разведчиц случайно окажется поблизости и, привлеченная запахом меда, начнет его набирать. Теперь можно считать игру выигранной и начать подготовку к опытам с уверенностью в том, что через несколько минут, кроме этой пчелы, у нашего столика окажутся десятки и даже сотни пчел. Если проследить, откуда прилетают эти пчелы, окажется, что все они, почти без исключения, принадлежат к той семье, что и первая обнаружившая источник взятка пчела. Итак, создается впечатление, что пчела каким-то образом сообщила дома, в улье, о богатой находке и привела к ней других пчел. Нам бы очень хотелось узнать, каким образом она это сделала. Чтобы выяснить это, есть только один способ: посмотреть, что делает возвратившаяся домой пчела и как ведут себя по отношению к ней другие пчелы. В обычном улье этого не увидишь, придется воспользоваться наблюдательным ульем. Выставим рядом с ульем кормушку. Пометим первую прилетевшую на нее пчелу, чтобы потом ее можно было узнать среди других пчел в улье и не потерять из виду. Вот она входит в леток, бежит вверх по сотам и затем оказывается где-нибудь среди своих ульевых подруг. Она отрыгивает из зобика собранный ею мед, появляющийся из ее ротового отверстия в виде блестящей капли, которая тотчас всасывается двумя или тремя ее подругами, протягивающими к ней хоботки. Они берут на себя заботу о его дальнейшем использовании и, смотря по обстоятельствам, либо кормят им своих голодных сограждан, либо заполняют медом ячейки, одним словом — занимаются внутренними делами, в которые сама сборщица не вникает. А между тем на соте разыгрывается представление, достойное пера прославлявших пчел великих поэтов. Но им оно еще не было известно, и поэтому читателю придется удовольствоваться его прозаическим описанием.
Круговой танец как средство взаимопонимания
Освободившись от груза, сборщица начинает своего рода круговой танец. Она бежит быстрыми, семенящими шагами по тому месту сота, где только что находилась, стремительно поворачиваясь то вправо, то влево, и, постоянно меняя таким образом направление, описывает один или два круга. Этот танец происходит в самой гуще пчел и особенно примечателен и интересен тем, что в него вовлекаются и окружающие танцовщицу пчелы. Те из них, которые находятся ближе других к танцовщице, семенят за ней и, вытянув усики, стараются коснуться ими ее брюшка. Они повторяют все повороты танцовщицы, так что она в своих стремительных движениях как бы тащит за собой целый хвост других пчел (рис. 1).
 Рис.
1. Круговой танец сборщицы нектара на
соте.
За танцовщицей следуют три
семенящие за ней пчелы, которые принимают
информацию
Рис.
1. Круговой танец сборщицы нектара на
соте.
За танцовщицей следуют три
семенящие за ней пчелы, которые принимают
информацию
Это кружение может продолжаться несколько секунд, полминуты или целую минуту. Затем танцовщица внезапно прекращает его, освобождается от свиты, часто еще на другом или третьем месте сота отрыгивает капельку меда — и снова начинает такой же танец. Закончив его, она спешит к летку и снова летит к кормушке, чтобы принести новую ношу. При каждом возвращении домой представление на сотах повторяется. В обычных условиях пчела танцует в темноте пчелиного улья. Следовательно, видеть танцовщицу ее товарищи по улью не могут. И если они могут заметить ее движения и повторять за ней все повороты, то руководствуются при этом исключительно своими осязательными и обонятельными восприятиями. Что должен означать этот круговой танец? Очевидно, он сильно возбуждает ближайших к танцовщице ульевых пчел. Пристально наблюдая за той или иной пчелой из свиты танцовщицы, можно увидеть, как она начинает готовиться к полету, слегка чистится, а затем пробирается к летку и покидает улей. Спустя некоторое время вслед за первой пчелой на кормушке появляются новые пчелы. Вернувшись домой нагруженными, они тоже танцуют, и чем больше танцовщиц, тем больше новичков прилетает на место кормления. Не приходится сомневаться в том, что существует взаимосвязь между этими явлениями. Танец в улье оповещает пчел о том, что обнаружен богатый взяток. Но как находят пчелы то место, куда следует лететь за ним? Прежде всего напрашивается предположение, что пчелы после окончания танца устремляются к летку вместе с танцовщицей и летят вслед за ней, когда она снова направляется к источнику корма. Однако тщательные исследования показывают, что это не совсем так. Новички, по-видимому, не знают, где находится цель их поисков. Символические жесты кругового танца указывают им только, что источник корма следует искать вокруг улья, и именно так они и поступают. Убедиться в этом можно при помощи простого опыта. Покормим небольшую группу меченых пчел на столике, стоящем примерно в 10 метрах к югу от улья. Затем немного подальше от улья в южном, западном, северном и восточном направлениях расставим в траве кормушки. Через несколько минут после того, как сборщицы, возвратившиеся с южной кормушки, начали танцевать, новички из нашего улья появляются на всех кормушках. Если спрятать корм от собирающих его пчел, они будут вести себя точно так же, как если бы вследствие неблагоприятной погоды прервался естественный взяток и самые обычные цветы перестали выделять нектар: пчелы остаются дома, танцы прекращаются. Теперь расставленные вокруг улья кормушки с медом могут часами и даже днями стоять в траве, не обнаруживаемые ни одной пчелой. Это может показаться странным, потому что небольшое число пчел, помеченных на кормушке, — это не единственные сборщицы семьи. В то время как они летали к чашечке с сахарным сиропом, сотни и тысячи их подруг посещали различные цветы, собирая с них пыльцу и нектар. Когда мы прекращаем кормление пчел на искусственной кормушке, остальные пчелы продолжают сбор нектара. Почему же, вернувшись с цветочного взятка, они при помощи танца не посылают своих подруг на его поиски во всех направлениях, а также и к кормушке? На это можно дать такой ответ: конечно, они посылали других пчел, если находили богатый источник взятка, но не к чашечке с сахарным сиропом, а к тем цветкам, которые они сами успешно использовали.
Биологическое значение цветочного запаха, рассмотренное с новой стороны
Не стеклянная чашка, а цветы — естественные сосуды с кормом для пчел. Мы действуем сообразно с природой, если на выбранном нами месте кормления вместо наполненной сиропом стеклянной чашки выставим маленький букет цветов, например альпийских фиалок. Чтобы иметь возможность использовать любые цветы и не зависеть от количества выделяемого ими в данный момент нектара, нанесем на каждый цветок по капле сиропа, который будем пополнять по мере выбирания его пчелами. Чтобы пчелы находили пищу в цветках и не могли собирать со стола случайно упавшие на него капли, поставим вазу с цветами в большое блюдо, наполненное водой. Помеченные пчелы, нашедшие богатый взяток в альпийских фиалках, танцуют на сотах. Поставим где-нибудь в стороне в траву чашку с альпийскими фиалками, которые не обрызгивали сахарным сиропом, а рядом — чашку с другими цветами, например с флоксами. Сигнал танцовщиц действует безотказно, и вскоре повсюду появляются пчелы, мечущиеся в поисках по всему лугу. Они подлетают также и к чашкам с цветами, опускаются на альпийские фиалки и упорно копошатся в них, как будто не сомневаясь в том, что здесь-то они и найдут что-нибудь подходящее. Но мимо чашки с флоксами они пролетают, не проявляя к ним никакого интереса. Теперь удалим с места кормления альпийские фиалки и заменим их флоксами, которые таким же образом обильно обрызгаем сахарным сиропом. Сборщицами оказываются те же самые пчелы, что и прежде, но они берут взяток уже не с альпийских фиалок, а с цветков флокса. На лугу все остается по-прежнему. Но уже через несколько минут картина меняется. Интерес к альпийским фиалкам ослабевает, вновь прибывающие пчелы летят только на флоксы, более того, везде в соседних садах, где есть только флоксы, мы видим пчел, усердно посещающих их цветки, — любопытное зрелище для каждого, кто знает, что глубокие трубочки венчика этого растения доступны лишь длинному хоботку бабочек и что пчелы совершенно не в состоянии достать спрятанный в их глубине нектар. Именно поэтому обычно их никогда не видно на флоксах. Совершенно очевидно, что разыскивающим взяток пчелам танцовщицы сообщили, что им нужно искать и какие именно виды цветов дают богатый взяток! Успех этого опыта не зависит от того, используем ли мы в качестве источника корма цветки альпийской фиалки или флокса, горечавки или вики, чертополоха или лютика, бобов или иммортелей. Целесообразность такого поведения пчел становится очевидной, как только мы представим себе естественные условия. Если пчелы-разведчицы находят какие-нибудь цветущие растения, они сообщают о своей находке, танцуя в улье. Мобилизованные ими пчелы устремляются именно к тому виду цветков, которые обильным выделением нектара вызвали танцы, и не тратят время на бесполезное отыскивание цветков без нектара. Объяснимо ли все это? Ведь невозможно допустить, чтобы в языке пчел имелись названия всех видов цветов. И все же это так. «Язык цветов» раскрывается здесь удивительно просто, целесообразно и восхитительно. В то время, когда сборщица высасывает сладкий сок из цветка, ее тело пропитывается его ароматом. Возвратившись домой, она все еще благоухает ароматом этих цветов, когда кружится в танце. Ее ульевые подруги, следующие за ней и так оживленно обследующие ее усиками (органами обоняния), во время танца воспринимают этот запах, запечатлевают его в памяти и руководствуются им, когда разыскивают взяток, обследуя окружающую местность. Если вместо цветов использовать эфирные масла или искусственные ароматические вещества, связь, существующая между пчелами и запахом, станет еще более очевидной. Накормим меченых пчел сиропом из стеклянной чашки, которая стоит на подставке, имеющей запах мяты. Возбужденные танцами, вылетевшие из улья новички вьются вокруг всех попадающихся им на глаза предметов, получивших этот запах после нанесения на них небольших количеств мятного масла. На другие запахи они не обращают внимания. Но стоит только переменить ароматическое вещество, нанесенное на подставку, как с переменой предлагаемого запаха изменится и цель пчелиных поисков. Однако первоначальным вариантом опыта, который мы принимаем за исходный, предусматривается кормление пчел на чашечке без запаха. В этом случае свита танцовщицы не может обнаружить какого-либо исходящего от нее специфического запаха. Но даже и теперь они вылетают из улья, зная, что все душистые цветы, встречающиеся на их пути, не заслуживают внимания и нет необходимости терять время на их посещение. Ботаники прошлых столетий видели в запахе цветов только средство для привлечения насекомых, разыскивающих пищу. Но для пчел запах, кроме того, служит опознавательным знаком, позволяющим им с уверенностью отличать цветки, на которых они уже побывали, от других, имеющих сходную окраску. Эта способность пчел — необходимая предпосылка их цветочного постоянства. Однако значение запаха заключается не только в этом. Подобно точным выражениям словесного языка, принесенный домой специфический запах просто и ясно сообщает пчелам в улье цель их поисковых полетов, к которым их побуждает танец.
Как пчелы приносят домой запах цветов
Не очень внимательный наблюдатель склонен считать, что многие цветы не имеют запаха. Еще издали бросается в глаза яркая окраска желтого лютика, голубого ластовника, красных бобов, но букеты из этих цветов не наполняют комнату ароматом. И все же тот, чье обоняние не притупилось от чрезмерного курения, может обнаружить нежный, свойственный каждому виду запах. Для этого достаточно собрать десяток таких цветков вместе и поднести их к носу. Среди насекомоопыляемых растений редким исключением являются такие, цветки которых полностью лишены запаха. К ним относятся брусника и дикий виноград. И действительно, при мобилизации на эти цветки у пчел в улье, как и следовало ожидать, не возникает никакого представления о цели поискового полета. Удивительно только, что даже самого слабого, едва ощутимого для нас цветочного запаха достаточно, чтобы пчелы в улье могли узнать, откуда прилетела танцовщица. Каким образом удается танцовщице доносить до дома тонкий запах цветов, на которых она побывала? Отчасти это можно объяснить тем, что ароматические вещества удерживаются на теле пчелы лучше, чем, например, на стекле, металле, бумаге, вате или даже на теле других насекомых. Человек может это проверить при помощи собственного обоняния. Еще нагляднее это можно показать, если надрессировать пчел на какой-нибудь цветочный запах, а затем предложить им на выбор положенные рядом тела пчел и другие предметы, которые пропитывались цветочным запахом в закрытых сосудах и после этого некоторое время проветривались. Ни на одном другом объекте невозможно так долго распознавать запах и ни один из них так упорно не посещают дрессированные пчелы, как тело пчелы. Ее внешний покров, по-видимому, приспособлен природой к тому, чтобы впитывать ароматы цветов. Но к сказанному необходимо добавить, что выделяющийся у основания цветка нектар, хранится в его душистой чашечке и поэтому приобретает специфический запах цветка. Взявшая его сборщица несет домой в своем медовом зобике вместе с нектаром пробу аромата, с которым она знакомит других пчел, скармливая им принесенную каплю нектара. Среди этих пчел есть и такие, которые при танце следуют за ней и, получив из ее рта душистый пароль, вылетают на поиски. Было бы очень интересно узнать, какой запах действеннее — тот, которым «надушена» пчела, или принесенный в медовом зобике? Это можно узнать, если поставить оба запаха в условия соревнования. Нанесем на цветки флокса капли сахарного сиропа и оставим их примерно на час, чтобы они пропитались запахом цветка. Затем дадим возможность нескольким пчелам, находящимся на цветках цикламена, через узкую щель брать из склянки сахарный сироп с запахом флокса (рис. 2). Во время танцев дома от их внешнего покрова будет исходить запах цикламена, а от раздаваемого ими сиропа — запах флокса.
 Рис.
2. Через узкую щель пчела набирает в
медовый зобик сироп с запахом флокса,
в то время как ее тело пропитывается
запахом цикламена. Мв — миска с водой,
закрытая сеткой
Рис.
2. Через узкую щель пчела набирает в
медовый зобик сироп с запахом флокса,
в то время как ее тело пропитывается
запахом цикламена. Мв — миска с водой,
закрытая сеткой
Чтобы увидеть результат, пронаблюдаем за чашечками с цветками флокса и цикламена, поставленными в траве поблизости от кормушки. Обе чашки посещаются новичками. Но запах, принесенный в зобике, выигрывает соревнование в том случае, когда источник корма находится на значительном расстоянии от улья. Опыт был повторен в условиях, когда расстояние между местом кормления и ульем составляло 600 метров. При дальних полетах тело пчелы проветривается сильнее и удерживаемый внешним покровом запах заметно теряет свою интенсивность. Вот почему мобилизованные новички руководствуются в своих поисках почти исключительно цветочным запахом нектара, доставленного в медовом зобике. Таким образом мы узнаем, какое биологическое значение имеет воспринявший запах цветков нектар, доставленный пчелами домой в медовых зобиках, как в хорошо закупоренных флакончиках.
Регулирование спроса и предложения
Танцы пчел приобретают свой полный биологический смысл только при таких обстоятельствах, когда они возникают под влиянием обильного взятка. При слабом взятке большая мобилизация невыгодна для семьи и танцев не происходит. Если срезать несколько цветущих веток, например акации, поставить их в сосуд с водой и защитить от насекомых, то в течение нескольких часов в цветках скопится много нектара. Теперь предложим этот букет группе пчел, которые до этого летали за сахарным сиропом на кормушку. Потребуется небольшая уловка, чтобы заставить их, не теряя времени, переключиться на посещение нового для них источника корма. Как только это произойдет, они начнут использовать естественный богатый источник взятка и быстро получат подкрепление в результате мобилизационных танцев. Но вскоре пчел станет так много, что они быстрее собирают и уносят нектар, чем он снова накапливается в цветочных чашечках. Вследствие излишка пчел взяток становится более скудным. И, хотя сбор продолжается с неослабевающим упорством, танцы прекращаются и группа пчел-сборщиц не получает нового пополнения из родного улья. Наряду с количеством степень «сладости» выделяемого нектара имеет также решающее значение для продуктивности взятка. Если в стакан с водой добавлять один кусок сахара за другим, то в конце концов сахар перестанет растворяться даже при продолжительном помешивании и в виде осадка опустится на дно стакана. Такой густой, «насыщенный» раствор сахара содержит столько сладкого вещества, сколько вода вообще может принять. Нектар некоторых цветков как раз и является таким насыщенным раствором. В этом случае его, конечно, стоит набирать как можно больше — сколько может войти в зобик — и мобилизовать на эту работу все силы семьи. В цветках других растений в это же самое время образуется жидкий, содержащий мало сахара нектар. При равном количестве жидкости в зобиках пчелы доставляют домой гораздо меньше сахара. Мобилизовывать сборщиц для использования этой находки так же энергично, как и в первом случае, нецелесообразно, и в действительности этого не происходит. Для того чтобы танцы пчел были оживленными и продолжительными, сахарный раствор должен не только выделяться в изобилии, но и быть очень сладким. Чем менее он сладок, тем более вялыми будут танцы, а чем слабее танец, тем незначительнее его вербующая сила. Если содержание сахара в нектаре снизится до определенного уровня, танцы прекратятся даже в том случае, если нектар выделяется в изобилии. Таким несложным способом регулируется мобилизация пчел-сборщиц в зависимости от продуктивности источника взятка. При одновременном цветении многих видов растений сильнее всего посещаются те из них, цветки которых производят больше, чем другие, более сладкого нектара. Пчелы, разыскавшие такие цветки, танцуют оживленнее, чем те, которые в это же время обнаружили менее богатые источники взятка. Специфический запах, приносимый домой пчелами-танцовщицами, определяет правильный выбор степени мобилизации сил семьи. С предельной ясностью, например, дается понять, что сегодня, судя по запаху, больше всего нектара удастся добыть в цветках сливы. Таким образом, в медовые кладовые пчел вливается нектарный поток в основном из источника, больше всего заслуживающего внимания в данное время. Одновременно и цветки, выделяющие наибольшее количество более сладкого нектара, лучше других посещаются пчелами и благодаря этому обеспечивают себя лучшим опылением и наиболее полным завязыванием семян.
«Флакончики с духами» на теле пчелы
Каждая рабочая пчела несет при себе готовый к употреблению «флакончик с духами», иначе говоря, маленькую парфюмерную фабрику. Она находится недалеко от кончика брюшка, в кожной складке, которая обычно загнута внутрь и поэтому не видна, но может произвольно выпячиваться в виде влажно блестящего валика. Тогда в воздухе распространяется пахучее вещество, выделяемое крошечными железами, расположенными в этом кожном кармане. Его аромат, напоминающий запах мелиссы, ощутим и для нас; для пчел он более интенсивен и воспринимается за несколько метров как привлекающий запах. Выше уже шла речь о том, как при «вилянии хвостиком» с помощью этого запаха одни пчелы указывают другим путь к летку. Сборщицы используют пахучий орган также и при посещении цветков, если взяток достаточно хорош и к нему желательно привлечь новые вспомогательные силы. Выделяя привлекающий запах, пчелы помогают отыскивать цель своим подругам, которых они подняли по тревоге танцами и заставили вылететь из улья. В этом нетрудно убедиться с помощью соответствующего опыта. Поставим вблизи наблюдательного улья две чашки с сахарным сиропом и на каждой из них позволим собраться по десятку пчел. Они прилетают из одного и того же улья, но каждая группа «знает» только свою чашку. Предложим теперь на одной чашке «хороший взяток» (сахарный сироп в изобилии), на другой — «скудный взяток» (промокательная бумага, смоченная сахарным сиропом таким образом, чтобы его можно было собрать лишь с трудом). Сборщицы, использующие богатый взяток, танцуют, другие — нет. К первой группе за одно и то же время присоединяется в десять раз больше новичков, чем ко второй. И это очень целесообразно. Как это происходит? Пчелы, находящиеся на соте, не могут знать, откуда прилетели танцовщицы, потому что цветочный запах не был придан ни одной из этих двух кормушек. Они обыскивают местность, не имея определенной цели. Но если они приблизятся к богатому источнику корма, то их притянет к нему привлекающий запах ранее прилетевших сюда сборщиц; в то же время они часто пролетают мимо кормушки, скудно снабженной сиропом, и не замечают ее. Что это действительно так, показывает контрольный опыт: можно как бы закупорить пчелиный флакончик с духами, заклеив кожный мешочек, в котором он находится, тонким слоем шеллачной пленки. Теперь пчелы уже не смогут раскрыть пахучую складку. Это не мешает пчелам-сборщицам в их работе. Они даже не замечают этого и при богатом взятке танцуют так же оживленно, как и прежде. Выставим на этот раз две чашки, наполненные сахарным сиропом. Обе группы пчел восторженно танцуют. Но та группа пчел, которые не могут издавать привлекающий запах, получают пополнение, составляющее лишь десятую часть от пополнения второй группы. Такую же роль, как на кормушках, пахучий орган играет и при посещении пчелами цветков.
Виляющий танец сообщает расстояние до источника корма
В течение многих лет опыты с кормушкой мы проводили только в непосредственной близости от улья. В районе улья новички быстро ориентировались и были многочисленны. Если контрольные чашки выставлялись на большем расстоянии, в то время как кормушка оставалась вблизи от улья, то новички прилетали на чашки тем позже и в тем меньшем числе, чем больше было расстояние. Это не вызывало удивления. Совершенно ясно, что мобилизованные пчелы прежде всего ищут корм поблизости от улья и, только если они ничего здесь не находят, радиус их полетов все более увеличивается. Но однажды, когда кормушка была установлена на расстоянии многих сот метров от улья, рядом с ульем ее искали только немногие новички, тогда как район далеко отставленной кормушки облетывался значительной группой пчел. Это возбудило подозрение, что танец указывает также, насколько далеко следует лететь. Если организовать опыт так, чтобы пронумерованные пчелы из наблюдательного улья собирали корм вблизи от него и одновременно другие меченые пчелы из той же семьи — на удаленной кормушке, то на сотах мы увидим поразительную картину: все пчелы, посещающие кормушки, расположенные вблизи улья, танцуют круговой танец (см. рис. 1), а пчелы, прилетающие с дальних кормушек, танцуют виляющий танец. При этом пчела пробегает некоторое расстояние по прямой, возвращается, делает полукруг к исходной точке, снова бежит по прямой и описывает полукруг в другую сторону. Такой танец может продолжаться несколько минут на одном и том же месте.
 Рис.
3. Виляющий танец
Рис.
3. Виляющий танец
Этот танец заметно отличается от кругового быстрыми виляющими движениями брюшка, которые производятся как раз во время прямолинейного пробега (виляющий пробег). Одновременно танцовщица издает шорох, воспринимаемый даже человеком, если вставить в ухо один конец пластмассового стетоскопа, а другой конец поднести к танцующей пчеле. Производимые при этом звуки можно зарегистрировать при помощи микрофона. Колебания будут воспроизводиться не как длинный продолжительный тон, а как повторяющиеся очень короткие вибрационные толчки. Каждый отдельный вибрационный толчок длится ничтожную долю секунды (15/1 000 секунды), такая же короткая пауза отделяет его от следующего. Частота колебаний отдельного тона составляет примерно 250 герц и соответствует частоте колебаний крыльев. Следовательно, эти свистящие шорохи производятся грудной мускулатурой крыльев, не сопровождаясь при этом настоящими ударами крыльев. Приближенно в течение секунды следуют один за другим 30 таких вибрационных толчков. Эта частота воспринимается нашим слухом как скрипучий шум. При помощи приклеенного к спинной части брюшка танцовщицы миниатюрного магнита можно записать эти колебания на электромагнитную пленку, причем виляющие движения брюшка записываются вместе с ними (рис. 4). Видно, что короткие вибрационные толчки независимо от виляющих движений перекрывают их, то есть они не связаны с определенной фазой виляющего движения.
 Рис.
4. Электромагнитная запись виляющих
движений с наложенными на них вибрационными
движениями. Изображенные над буквой а
виляющие движения на рис. б воспроизведены
в увеличенном виде. (По Г. Эшу)
Рис.
4. Электромагнитная запись виляющих
движений с наложенными на них вибрационными
движениями. Изображенные над буквой а
виляющие движения на рис. б воспроизведены
в увеличенном виде. (По Г. Эшу)
Однако продолжительность звучания точно соответствует продолжительности виляющего пробега, следовательно, этот пробег «подчеркивается» не только виляющим движением, но и издаваемым одновременно звуком. Уже упоминалось, что пчелы хотя и не «слышат» колебания, передаваемые по воздуху, но очень чувствительны к вибрации вещества, на котором они находятся. Поэтому они могут воспринимать скрипучий шум танцовщицы, следуя за ней по соту. Виляющий танец, и особенно фаза прямолинейного пробега, с большим вниманием прослеживается следующими за танцовщицей пчелами. Если стоящую близко от улья кормушку переставлять постепенно все дальше и дальше, то, когда расстояние достигнет 50 — 100 метров, круговые танцы сборщиц переходят в виляющие. Когда кормушка оказывается между 100 и 50 метрами, виляющие танцы сменяются круговыми. Круговой и виляющий танцы — различные выражения пчелиного языка, сообщающие, близко или далеко расположен источник взятка. Именно в этом смысле их и понимают пчелы в улье. Одним только указанием «ближе 100 метров» или «дальше 100 метров» пчелам, которые должны вылететь и найти источник корма, была бы оказана слабая помощь, так как зона их лета простирается на многие километры во все стороны от родного улья. При постепенном перемещении кормушки до границы зоны их полета обнаруживается закономерное изменение виляющего танца, позволяющее пчелам и наблюдающему за ними исследователю получить более точное представление о расстоянии до источника взятка. При расстоянии 100 метров танцы становятся стремительными и повороты (см. рис. 3) быстро следуют один за другим. Чем значительнее расстояние, тем умереннее становится темп танца, тем медленнее следуют один за другим повороты, тем устойчивее и продолжительнее прямолинейный виляющий пробег. При помощи часов можно установить, что при удалении кормушки на 100 метров пчела делает за четверть минуты примерно от девяти до десяти прямолинейных пробегов во время танца, при расстоянии 500 метров — около шести пробегов, при расстояниях 1 000 метров — от четырех до пяти пробегов, 5 000 метров — два пробега и 10000 метров — в среднем чуть больше одного пробега (рис. 5).1
 Рис.
5. Кривая наглядно показывает, как
замедляется темп танца с увеличением
расстояния. Слева: число виляющих
пробегов за 1/4 минуты; внизу: расстояние
от улья до кормушки в километрах
Рис.
5. Кривая наглядно показывает, как
замедляется темп танца с увеличением
расстояния. Слева: число виляющих
пробегов за 1/4 минуты; внизу: расстояние
от улья до кормушки в километрах
Многочисленные наблюдения показывают, что при этом сигнал, имеющий отношение к расстоянию, связан с продолжительностью виляющего пробега, «временем виляния», которое так выразительно подчеркивается виляющими движениями и производимым звуком. Пчелы должны обладать тонким чувством времени, благодаря которому танцовщица, двигаясь в соответствующем ритме, способна информировать своих подруг так, чтобы они могли правильно понять и оценить эту информацию. Действительно ли она способна к этому? Насколько точно придерживаются новички, вылетающие из улья, расстояния, указанного им при помощи виляющего танца? Чтобы узнать это, на определенном расстоянии от улья покормим несколько пронумерованных пчел сахарным сиропом, установив кормушку на подставке, имеющей слабый запах лаванды. Разложим такие же душистые приманки, но только без корма, на разных расстояниях от улья. Пчелы-сборщицы танцуют на сотах и посылают своих подруг на поиски «ресторана», издающего запах лаванды. В одном таком опыте кормушка была установлена за 750 метров от улья, а ароматизированные пластины разложены в том же направлении на расстоянии 75; 200; 400; 700; 800; 1000; 1 500; 2 000 и 2 500 метров от улья. Возле каждой из них находился наблюдатель, регистрировавший в течение 1,5 часа каждую прилетающую пчелу. На (рис. 6, а) приведено число новичков, которые появились на разных пластинах, а вычерченная кривая делает результат опыта более наглядным. В другом опыте кормушку ставили на расстоянии 10; 100; 400; 800; 1 200; 1 600; 1 950; 2 050; 2 400; 3 000, 4 000 и 5 000 метров от улья (рис. 6, б). Сверх ожидания мобилизованные пчелы строго следовали указаниям танца, часами упорно обыскивая район расположения кормушки.
 Рис.
6. Результат двух «ступенчатых опытов».
В первом опыте (а) кормушка с несколькими
пронумерованными пчелами находилась
в 750 метрах от улья, во втором опыте (б)
— в 2 километрах. Кривые показывают
число новичков, появившихся на
соответствующих наблюдательных пунктах
Рис.
6. Результат двух «ступенчатых опытов».
В первом опыте (а) кормушка с несколькими
пронумерованными пчелами находилась
в 750 метрах от улья, во втором опыте (б)
— в 2 километрах. Кривые показывают
число новичков, появившихся на
соответствующих наблюдательных пунктах
Но откуда пчелы могут знать, на какое расстояние им нужно лететь? Некоторое представление о их способе измерения расстояния дают наблюдения, проведенные в ветреную погоду. Если при полете к кормушке пчелам приходится лететь против ветра, то по возвращении домой они показывают в танцах большее расстояние, чем в безветренную погоду, а при попутном ветре — меньшее. Если в безветренную погоду им приходится лететь к месту сбора взятка круто в гору, то это сказывается на танцах так же, как удлинение расстояния, а полет вниз, — как его сокращение. Вероятно, мерилом определения ими расстояния служит энергия, затрачиваемая на полет к источнику взятка.
Виляющий танец указывает пчелам также и направление к источнику взятка
Пчелиная семья получила бы мало пользы, узнав, что за 2 километра от улья цветет липа, если бы одновременно не давалось указаний о направлении, в котором ее нужно искать. Виляющий танец содержит и такое сообщение. Оно кроется в фигуре танца, а именно в направлении прямолинейного виляющего пробега. Сообщая направление, пчелы пользуются двумя различными способами в зависимости от того, танцуют ли они, как обычно, на вертикальной поверхности сота в улье или на какой-либо горизонтальной поверхности, например на прилетной доске. Указание направления на горизонтальной поверхности следует рассматривать как исторически более старую форму. А так как она к тому же более понятна, с нее мы и начнем. Вспомним о том, что солнцем пользуются как компасом. Если при полете от улья к кормушке солнце находилось от сборщицы, например, впереди слева под углом 40°, то, вернувшись в улей, она в прямолинейном виляющем пробеге придерживается того же угла по отношению к солнцу и указывает таким образом прямо на кормушку (рис. 7). Семенящие за ней пчелы обращают внимание на то положение, которое она занимает по отношению к солнцу во время виляющего пробега. Благодаря этому при вылете из улья они принимают то же положение, что и сборщица, и избирают правильное направление к источнику корма.
 Рис.
7. Указание направления по солнцу при
танце на горизонтальной поверхности.
Слева:
Ну — наблюдательный улей; К — кормушка;
— — — — направление полета к месту
взятка. Справа: виляющий танец на
горизонтальной поверхности
Рис.
7. Указание направления по солнцу при
танце на горизонтальной поверхности.
Слева:
Ну — наблюдательный улей; К — кормушка;
— — — — направление полета к месту
взятка. Справа: виляющий танец на
горизонтальной поверхности
Но происходит это только тогда, когда танцовщица видит солнце или по-крайней мере голубое небо, танцуя, например, на прилетной доске, что нередко бывает в теплую погоду, когда приемщицы встречают возвращающихся сборщиц у входа в улей. Можно также вынуть один сот из улья и подержать его в горизонтальном положении под открытым небом. Танцующих пчел не так легко привести в замешательство. Они указывают направление в ту сторону света, где брали взяток, и если вращать лежащий сот, как поворотный круг на железной дороге, то они, позволяя танцевальной площадке вращаться под их ногами, подобно стрелке компаса, продолжают указывать нужное направление. Но, если только небо будет закрыто от их глаз, наступит полная дезориентация и в танцах начнется путаница. Внутри улья темно, неба не видно, а соты расположены вертикально. Все это не позволяет указывать направление тем путем, с которым мы только что познакомились. При этих обстоятельствах пчелы пользуются вторым, еще более замечательным способом. Принимая за основу угол между направлением, в котором они летели к кормушке, и прямой линией к солнцу, они сохраняют его в танце, используя при этом в качестве одной из составляющих направление силы тяжести. При этом виляющий пробег вверх означает, что кормушка расположена от улья прямо по направлению к солнцу; виляющий пробег вниз говорит о противоположном направлении; пробег, например, под углом 60° влево от направления вверх указывает на то, что кормушка находится влево под углом 60° от прямого направления от улья к солнцу (рис. 8), и так далее. То, что новички благодаря тонкому восприятию направления силы тяжести узнают таким образом в темном улье, они используют затем при вылете применительно к направлению на солнце.
 Рис.
8. Указание направления по солнцу при
танце на вертикальной поверхности сота.
Слева показана ориентировка виляющего
танца на вертикальном соте при данном
положении кормушки
Рис.
8. Указание направления по солнцу при
танце на вертикальной поверхности сота.
Слева показана ориентировка виляющего
танца на вертикальном соте при данном
положении кормушки
С помощью того же метода «ступенчатого опыта», который применялся при исследовании сообщения о расстоянии, можно выяснить, следуют ли мобилизованные пчелы полученным указаниям относительно полета в определенном направлении. В качестве примера на рис. 9 приведен результат одного такого опыта. На кормушке, находящейся за 250 метров от улья, брали корм несколько пронумерованных пчел. Подставка кормушки имела запах. В 200 метрах от улья на равных расстояниях одна от другой, соответствующих углу 15°, были веерообразно размещены пластины с запахом. Приведенные данные показывают, сколько новичков было обнаружено во время полуторачасового опыта на наблюдательных пунктах. Лишь немногие из них отклонились от правильного пути.
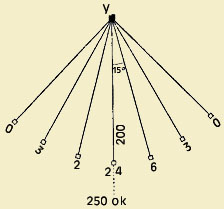 Рис.
9. Результаты «веерного» опыта. У — улей;
К — кормушка.
Маленькие квадратики
обозначают выставленные душистые
приманки без корма. Цифры показывают
количество посетивших их новичков
Рис.
9. Результаты «веерного» опыта. У — улей;
К — кормушка.
Маленькие квадратики
обозначают выставленные душистые
приманки без корма. Цифры показывают
количество посетивших их новичков
В тропиках дважды в году солнце в полдень стоит в зените, то есть не находится ни в какой определенной стране света, и определить направление к цели по местонахождению солнца в отношении стран света невозможно. Как же поступают в этом случае пчелы? Они решили эту проблему поразительно просто: остаются дома. Как только солнце приблизится к зениту, пчелы устраивают себе обеденный перерыв даже в те дни, когда тропическая жара в полдень не настолько велика, чтобы нельзя было использовать имеющийся в природе хороший взяток. Только путем особого вмешательства их можно заставать посещать кормушку и в эти часы, но в таком случае, возвратившись домой, сборщицы танцуют беспорядочно, делая прямой пробег во всех направлениях. Этого и следовало ожидать, и это служит подтверждением их способности ориентироваться по солнцу. Неожиданно оказалось, что угол 2 — 3° от зенита уже вполне достаточен для того, чтобы пчелы могли определить направление по положению солнца и правильно передавать его в танце. Фасеточные глаза, укрепленные неподвижно на головной капсуле и состоящие из тысяч слегка наклоненных отдельных глазков, представляют собой удивительные приспособления для измерения даже таких небольших углов. В горах и крылатые существа не всегда прямым путем могут достигнуть цели. Какими жестами пользуются пчелы, чтобы указать своим товарищам окольный путь к источнику корма? В скалистой местности, в районе озера Вольфгангзее, имеется богатый выбор возможностей для изучения этого вопроса. Однажды наш наблюдательный улей был помещен на горе Шафберг за гребнем скалы, а кормушку, которую быстро накрыли вместе с находящимися на ней мечеными пчелами, перенесли вокруг края обрыва к месту. Рис. 10 воспроизводит план местности, в которой проводился опыт, и расстояние от улья до кормушки. Сборщицы летали туда и обратно кружным путем, изображенным на чертеже в виде острого угла, но в танцах они не указывали направления, в котором фактически летели от улья; они также не принимали в расчет вторую сторону острого угла, описываемого ими при полете,— оба эти указания могли бы ввести в заблуждение их подруг.
 Рис.
10. План местности, где проводился опыт.
У
— улей с пчелами; К — кормушка: — — —
кружной путь, которым летали пчелы;
....... прямой воздушный путь к кормушке
Рис.
10. План местности, где проводился опыт.
У
— улей с пчелами; К — кормушка: — — —
кружной путь, которым летали пчелы;
....... прямой воздушный путь к кормушке
Виляющий пробег танцовщиц показывал направление от улья к кормушке по воздушной прямой линии, по которой они никогда не летали. Только так могли они правильно направить своих подруг по улью к месту взятка. Мобилизованные пчелы искали в указанном направлении и, перелетев через препятствие, достигали цели. Познакомившись с источником взятка, они тоже нашли более легкий путь вокруг хребта. Поведение указывающих направление пчел было вполне целесообразным. Но то, что, проделав окольный путь, они оказались в состоянии построить действительное направление без транспортира, линейки и чертежной доски, — это относится, вероятно, к наиболее удивительному чуду из чудес, которыми так богата жизнь пчел. Это невольно наводило на мысль, что для каждой все более трудной задачи пчелы обязательно находят решение. И все же однажды они не знали, как помочь себе. Улей находился за редкой металлической решеткой конструкций, внутри радиобашни. Кормушка с помощью лебедки и привязанного к тросу кормового столика была помещена на верхушку башни, точно над летком родного улья. Выражение для понятия «вверх» пчелиным лексиконом не предусмотрено, так как в облаках цветы не растут. Сборщицы, побывавшие на верхушке башни, не знали, как сообщить направление, и совершали круговые танцы, которые мобилизовали их подруг на поиски взятка на земле во все стороны луга, и ни одна из них не нашла пути к источнику взятка, находящемуся наверху. Когда кормушка была перенесена на луг на расстояние, соответствующее высоте башни, система, указывающая направление, стала функционировать безупречно. Виляющий танец с его прямолинейным поступательным виляющим пробегом, а также круговой танец с его круговыми пробегами, по-видимому, с изумительной символической выразительностью призывают к действию: один — направляя вдаль, а другой — к поискам вокруг родного улья. Благодаря хорошо отрегулированной системе те пчелы, которые должны лететь на далекое расстояние, получают точные указания о цели путешествия. Но, когда мобилизуются и следуют указаниям танца сотни новичков, среди них часто находятся отдельные пчелы, которые действуют по-другому. Некоторые пчелы, присутствовавшие при круговых танцах, ищут взяток вдали, а наблюдавшие за виляющими танцами — вблизи улья (см. рис. 6) или в неправильном направлении от него. Разве они не поняли языка пчел? Или это упрямцы, которые предпочитают сами выбирать дорогу? Однако, рассмотрев явления, послужившие поводом для такого «неправильного» образа действий, в целом следует сделать вывод, что такие «оригиналы» могут быть весьма полезны. Если где-то цветет рапс, то хотя и хорошо было бы быстро послать туда множество пчел из улья, но не мешает также разузнать, не раскрылись ли в это же время бутоны цветков рапса и на другом поле. Этим оригиналам, которые не следуют схеме, мы обязаны тем, что все источники взятка в зоне лета пчелиной семьи быстро разыскиваются и используются.
Танцы сборщиц пыльцы
Наряду с медом в качестве второго незаменимого продукта питания пчелиная семья собирает цветочную пыльцу. Сборщицы пыльцы также сообщают друг другу об обнаруженном обильном источнике взятка, и делают они это тем же способом, что и сборщицы нектара. Они используют круговые танцы при близком и виляющие танцы при удаленном источнике взятка, чтобы передать сообщение о расстоянии и направлении. Однако небольшое отличие все же есть: у сборщиц нектара представление о виде цветов передается при помощи цветочного запаха, задержавшегося на теле и принесенного в медовом зобике. Сборщицы пыльцы не приносят домой душистый нектар, но зато они доставляют небольшую частицу посещенных ими цветов — цветочную пыльцу. Она имеет специфический запах, заметно отличающийся от запаха лепестков и различный у каждого сорта цветов. Таким образом, пыльцевые штанишки можно назвать душистыми посланиями. Это доказывается следующим опытом. Подготовим для сборщиц пыльцы два места кормления. На одном из них (рис. 11, Ш) группа меченых пчел собирает пыльцу с шиповника, на другом (К) — вторая группа ведет сбор этого корма с крупных цветков колокольчика. Удалим цветы с обоих пунктов и устроим перерыв в кормлении. Спустя некоторое время, не найдя пыльцы, сборщицы останутся дома, и только одна из них время от времени вылетает на разведку, чтобы посмотреть, не появилось ли здесь опять что-нибудь пригодное для сбора. Выставим на том месте, где стояли колокольчики, свежий букет этих же цветов. Разведчица тотчас соберет обножку, полетит домой и начнет танцевать.
 Рис.
11. У — улей с пчелами, Ш - чашка с цветами
шиповника; К — чашка с цветками
колокольчика (Объяснения см. в тексте)
Рис.
11. У — улей с пчелами, Ш - чашка с цветами
шиповника; К — чашка с цветками
колокольчика (Объяснения см. в тексте)
Первыми после перерыва обратят внимание на ее танец пчелы, уже раньше собиравшие пыльцу с колокольчиков, так как знакомый запах как бы говорит им, что цветы снова выделяют пыльцу. Немедленно поспешат они к цветкам колокольчика, на которых вскоре благодаря продолжительным танцам появляются новички. Но сборщицы пыльцы с шиповника остаются в улье, так как запах цветков колокольчика им незнаком. Этого еще недостаточно, чтобы выяснить, что же имеет решающее значение — запах лепестков или запах пыльцы. Проведем опыт несколько иначе. Устроим еще раз на обоих местах кормления перерыв, после которого на том же месте снова предложим пчелам колокольчики, предварительно заменив их собственные пыльники пыльниками шиповника (рис. 12).
 Рис.
12 а — цветок колокольчика; часть
цветочного венчика удалена и видна
внутренность цветка; пыльца с загнутых
вниз пыльников большей частью прилипла
к пестику, б — цветок колокольчика, из
которого удалены все части, содержащие
пыльцу, и заменены пыльниками шиповника;
в — цветок шиповника; г — цветок шиповника
после удаления его пыльников. Внутрь
цветка помещены пыльники вместе с двумя
пестиками из цветков колокольчика. Пл
— пыльники; Пс — пестик
Рис.
12 а — цветок колокольчика; часть
цветочного венчика удалена и видна
внутренность цветка; пыльца с загнутых
вниз пыльников большей частью прилипла
к пестику, б — цветок колокольчика, из
которого удалены все части, содержащие
пыльцу, и заменены пыльниками шиповника;
в — цветок шиповника; г — цветок шиповника
после удаления его пыльников. Внутрь
цветка помещены пыльники вместе с двумя
пестиками из цветков колокольчика. Пл
— пыльники; Пс — пестик
Прилетевшая пчела-разведчица находит на привычном месте знакомые ей цветки колокольчика и собирает в них обножку. Таким образом, одна из пчел, посещавших раньше цветки колокольчика, на прежнем месте и в тех же цветках колокольчика собирает обножку из пыльцы шиповника. Прилетев домой, она танцует, но все ее подруги, которые целыми часами и днями собирали вместе с ней пыльцу на цветках колокольчика, не обращают ни малейшего внимания на ее оживленный виляющий танец. Напротив, совершенно «чужие» для нее сборщицы пыльцы с шиповника спешат к ней, обнюхивают ее обножку и летят туда, где они привыкли собирать пыльцу с шиповника и где теперь напрасно ищут его цветки. Хотя пчелы дали себя одурачить, но благодаря этому стало известно, что не запах цветков колокольчика, в которых копошилась пчела, а запах принесенной ею цветочной пыльцы шиповника играет решающую роль. Обратный опыт дает аналогичный результат. Разведчица, которая собирает в цветках шиповника пыльцу колокольчика, мобилизует при помощи танцев сборщиц пыльцы с колокольчика.
Об опрокинутом улье и о доказательствах восприятия пчелами поляризованного света
Чтобы точнее изучить поведение пчел на горизонтальной танцевальной площадке, лучше всего использовать наблюдательный улей, который можно наклонять и при помощи винта и гайки закреплять в любом положении. Если поверхность сота наклонена хотя бы на 15°, танцовщицы уже могут указать направление к солнцу посредством виляющего пробега вверх и правильно показать любой угол между направлением к кормушке и направлением к солнцу. Но если сот лежит горизонтально, то бежать вверх по нему невозможно. Тогда ключ, о котором шла речь выше (см. рис. 8), уже не подходит для указания направления. Забавно наблюдать, как пчелы при таких обстоятельствах с неизменным усердием продолжают танцевать, но без какой-либо ориентировки виляющего пробега (так как небо для них невидимо). Направление пробега в этом случае совершенно беспорядочно и постоянно меняется. Как только танцовщица увидит солнце или кусочек голубого неба, танцы становятся ориентированными и указывают направление на кормушку. Уже говорилось о том, что эта поразительная настройка на голубое небо объясняется поляризацией небесного света. Несколько слов о том, как это можно доказать. Наблюдательный улей в горизонтальном положении. Стеклянные стенки его над сотами закрыты доской, в которой прорезано четырехугольное окошечко. Над ним в круглой вращающейся раме размещен большой поляризатор. При проведении опыта улей закрывают с трех сторон и танцующие на соте пчелы видят через окошечко в доске ограниченную, например северную, часть голубого неба. Небесный свет частично поляризован и, как мы уже знаем, на каждом участке неба характеризуется определенным направлением колебаний. Поляризатор преобразует его еще значительнее, пропуская лучи только с какой-либо одной плоскостью колебаний. Установим теперь вращающийся поляризатор над танцующей пчелой так, чтобы проходящие через него световые лучи сохраняли плоскость колебаний, которую они имели на северной стороне неба. При этом пчелы продолжают танцевать правильно и указывают действительное направление к кормушке. Но стоит только повернуть поляризатор, изменив тем самым направление колебаний поляризованного света, как танцующие пчелы отклонятся в сторону поворота и покажут неверное направление. Поправка к ошибочному указанию при этом не всегда точно совпадает с углом, на который был повернут поляризатор. От чего зависит величина неверного указания и действительно ли она определяется направлением колебаний поляризованного света, можно проверить, если воспользоваться поляризатором. По отношению к поляризованному свету его можно рассматривать как модель отдельного пчелиного глазка в сложном составном органе зрения (фасеточном глазу) пчелы. Глядя через поляризатор на небо, попробуем уяснить себе, как оно может воздействовать на пчелиный глаз. В качестве примера рассмотрим такой опыт. Группа пронумерованных пчел из лежащего горизонтально наблюдательного улья берет сахарный сироп на кормушке, расположенной точно на западе. Танцовщицы могут видеть часть голубого неба на западе, то есть в направлении к кормушке. Их виляющие пробеги указывают на запад. Расположим большой круглый поляризатор так, чтобы направление колебаний света от западной части неба осталось неизменным. Пчелы продолжают танцевать правильно. Их танец, очевидно, соответствует образцу освещенности, характерному для западной части неба, который определяется направлением колебаний поляризованного света. Этот образец можно увидеть, если посмотреть через поляризатор на западную часть неба. При полете к кормушке пчелы придерживаются направления, при котором они все время видят этот образец прямо перед собой. В танце они становятся так, чтобы при виляющем пробеге он также был всегда впереди. Поэтому они и указывают верное направление на запад (рис. 13, а; образец М2 сначала не будем принимать во внимание).
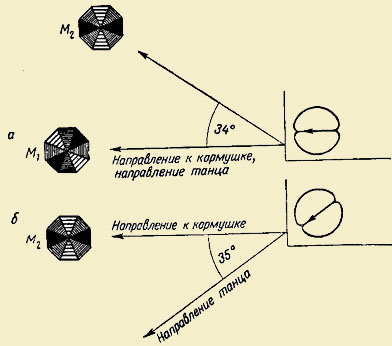 Рис.
13. Вверху: пчелы берут сироп из кормушки,
расположенной к западу от наблюдательного
улья. Если они при танце на горизонтальной
поверхности сота видят западную часть
неба, то указывают правильно на запад.
Они ориентируются при виляющем пробеге
по образцу освещенности М1, который
они видят прямо перед собой при полете
к кормушке. Нижний рисунок: посредством
поляризатора образец М1 заменяется
для танцовщиц образцом М2 Они
указывают ошибочное направление, потому
что при полете к кормушке видели образец
М2 под углом 34° вправо от направления
их полета
Рис.
13. Вверху: пчелы берут сироп из кормушки,
расположенной к западу от наблюдательного
улья. Если они при танце на горизонтальной
поверхности сота видят западную часть
неба, то указывают правильно на запад.
Они ориентируются при виляющем пробеге
по образцу освещенности М1, который
они видят прямо перед собой при полете
к кормушке. Нижний рисунок: посредством
поляризатора образец М1 заменяется
для танцовщиц образцом М2 Они
указывают ошибочное направление, потому
что при полете к кормушке видели образец
М2 под углом 34° вправо от направления
их полета
Если повернуть поляризатор на 30° против часовой стрелки, направление танца сразу изменится и пчелы укажут направление на 35° южнее. На западной части неба через шестигранный звездчатый поляризатор, служащий для нас моделью сложного пчелиного глаза, виден образец М1. Поместим обычный поляризатор перед шестигранным поляризатором и придадим ему такое же положение, какое занимает поляризатор, находящийся над ульем; затем переменим образец М1 на образец М2. Можно предполагать, что подобное действие происходит в пчелином глазу. Если снова удалить находящийся перед искусственным глазом поляризатор и направить этот глаз на небо, то образец М2 появится только в одном месте, а именно на 34° севернее западного направления. Следовательно, пчелы при полете от улья к кормушке видели точно перед собой образец М1 и на 34° правее от себя — образец М2 и должны были держаться таким образом, чтобы на 34° правее их возникал образец освещенности М2. В опыте мы показали им образец освещенности М2 на западе, то есть перемещенным влево на 34°. Они не видели ничего другого, кроме этого образца, и по нему ориентировали свой танец, указывая направление на 35° влево от этого образца, как при полете к кормушке, с ошибкой только на 1°. Конечно, лишь один такой опыт нельзя считать достоверным. Однако, после того как примерно в сотне всесторонне измененных экспериментов такого рода результаты всегда в основном оказывались одинаковыми, появилась возможность с уверенностью считать, что пчелы ориентируются по поляризованному свету.
Танцы на рое
С помощью танцев пчелы могут сообщать сведения не только об источниках нектара и пыльцы, но также и о других целях своих полетов, например о том, где находятся лужи с пригодной для сбора водой, которая нужна не только для утоления жажды, но также иногда в больших количествах для охлаждения улья, или о том, где можно собрать с почек деревьев прополис, чтобы промазать улей и заделать в нем щели. Особый интерес представляют танцы разведчиц, подыскивающих жилище и сообщающих пчелиному рою положение пригодного для гнезда места. Непосредственно после роения пчелы собираются вокруг матки роевой гроздью, повисающей в большинстве случаев на каком-нибудь растущем поблизости дереве. Обязанность пчел-разведчиц — разыскать приют для новой пчелиной семьи: дупло дерева, расщелину скалы, полость в стене, пустой улей пли что-нибудь подобное. Десятки пчел-разведчиц отправляются в разные стороны. Проходит немного времени, и одна здесь, другая там находит какое-нибудь место, которое принимается во внимание, даже если оно расположено на расстоянии нескольких километров. Удачливые разведчицы, возвратившись, танцуют на поверхности роевой грозди и показывают направление и расстояние до найденного ими приюта, точно так же как пчелы-сборщицы указывают направление и расстояние к месту источника корма. На пчелином рое можно наблюдать танцующих разведчиц, число которых непрерывно возрастает. Одни из них указывают небольшие, другие, напротив, — довольно значительные расстояния; одна пчела показывает одно, другая — иное направление в соответствии с расположением обнаруженного жилища. При этом бросается в глаза, что мобилизация пчел разными разведчицами происходит с неодинаковой энергией. Как при очень богатом взятке или концентрированном нектаре, мобилизационные танцы пчел очень оживленны, а при убывающем поступлении нектара они становятся все слабее и слабее, так и пчелы-разведчицы танцуют тем оживленнее, чем больше обнаруженное ими жилище соответствует требованиям пчелиной семьи. Характер их танцев зависит от многих причин: размеров дупла, расположения летка и его защищенности от ветра (в жилом помещении не должно быть сквозняка) и даже от запаха внутри дупла, который должен быть приятен для пчел, и кто знает, от скольких еще подобных вещей! На протяжении нескольких часов, а иногда даже нескольких дней происходит нечто удивительное. Наиболее энергичные танцовщицы приобретают все больше и больше сторонников среди своих товарищей, которые, исследовав облюбованное пчелами-танцовщицами место и убедившись в его достоинствах, сами принимаются усиленно пропагандировать его при помощи танцев. Танцовщицы, старавшиеся до этого направить рой в менее пригодное жилище, тоже заражаются вихревыми движениями более удачливых разведчиц и позволяют подчас «переубедить» себя. Сначала они устремляются за своими «конкурентами», танцующими на роевой грозди, а потом, следуя их указаниям, осматривают облюбованное ими жилище и наконец сами направляют пчел туда же. Многие из тех пчел, которые не могут отстаивать выбранное ими жилище с такой же энергией, как другие, более счастливые разведчицы, при подобном ходе событий просто-напросто перестают танцевать. Таким образом, все приходят к общему согласию, дружно танцуя в одном направлении, и, как только это произойдет, роевая гроздь распадается и под предводительством сотен своих подруг, которые уже знают дорогу, пчелы летят к жилью, признанному лучшим из всех обнаруженных. Человеку действительно есть чему поучиться у пчел. Но, вместо того чтобы посмотреть, как они, к всеобщему удовольствию, согласовывают различные мнения, он собирает рой в роевню, грубо разрушая при этом в своих интересах естественный ход событий.
Значение танцев пчел для пчеловодства и сельского хозяйства
Тот, кто едет за границу, владея соответствующим языком, получит больше удовольствия и пользы от поездки, чем его спутник, не знающий иностранного языка. Примерно так же обстоит дело и с пчеловодом в его отношениях с пчелами. Зная их «язык», он получает возможность заставить пчел лучше служить его целям. Когда время главного взятка миновало, кое-какие растения продолжают еще цвести. Однако их нектарные фонтанчики уже не так обильны, как несколько недель назад. Опытный пчеловод знает, что цветки осота, простирающие теперь повсюду к небу сотни тысяч своих высоких, окруженных зелеными чашелистиками цветочных головок, могли бы дать ему еще несколько килограммов меда. Но у пчел уже нет больше настоящей тяги к собирательной деятельности. На головках осота можно обнаружить главным образом шмелей. Они имеют преимущество перед пчелами благодаря своим более длинным хоботкам, а нектара в цветках этих растений не так уж много, чтобы танцы могли заметно усилить их посещение пчелами. Пчеловоду хочется сказать своим пчелам, что им не следует сидеть дома без дела и стоило бы попытаться достать из головок осота то, что там еще есть. Он сможет сделать это, если будет знать их язык. Ему достаточно привлечь несколько пчел из семей своей пасеки при помощи небольшого количества меда или сахарного сиропа на букет осота и покормить их на его цветках, обрызганных сахарным сиропом. Вернувшись домой, эти пчелы танцуют, сообщая при помощи принесенного ими запаха об открытом источнике взятка и о цели их мобилизационного призыва. Вскоре их подруги начинают вылетать из улья и искать запах осота. Посещение головок осота повышается во много раз. На практике этот прием всячески изменялся и упрощался. Вместо того чтобы кормить пчел на цветках осота, можно давать им сахарный сироп с ароматом цветков осота в улье. Такой сироп получают, настаивая чистый сахарный сироп в течение нескольких часов на цветках осота. Цветки других видов, запах которых после погружения в сахарный сироп изменяется, вследствие чего они становятся непригодными для этой цели, кладут в кормушку сухими и, налив в нее немного сахарного сиропа, ставят перед летком пчелиной семьи. В результате, затрачивая незначительные усилия, можно получать более высокие сборы меда с осота и с других медоносов в такие периоды, когда на соседних пасеках пчелы ничего не собирают. Агроному также нередко требуется направить пчел на определенные растения для улучшения их опыления и завязывания семян. Так, например, получение столь необходимых семян одного из наших важнейших кормовых растений, красного клевера, — дело трудное и ненадежное. Пчелам нелегко дотянуться своими короткими хоботками до основания цветочной трубочки, чтобы добыть нектар из этих шмелиных цветков. Там, где на полях систематически возделывается красный клевер, шмелей оказывается слишком мало, чтобы опылить миллионы отдельных цветков, а пчелы не проявляют никакого стремления посещать поля красного клевера, не отличающиеся обилием нектара, и охотнее обращаются к более надежным источникам взятка. Лишь в отдельные, редкие годы, когда красный клевер выделяет много нектара, пчелы охотнее посещают его. Обычно урожаи семян клевера низки. Этому горю, однако, легко помочь. На краю клеверного поля ставят ульи с пчелами. Затем описанным выше способом, при помощи сиропа с запахом цветков красного клевера, мобилизуют пчел, добиваясь таким путем усиления его посещаемости. Этот прием повышает урожай семян в среднем на 40%. Благодаря надежным урожаям прием «управления летной деятельностью пчел при помощи запаха» (дрессировка) быстро привлек внимание опытных семеноводов.2 Применяемый пока лишь в некоторых местностях, он, безусловно, скоро получит широкое распространение в районах интенсивного земледелия. Ведь для того, чтобы направить пчел на работу, пользуясь их же «языком», требуется немного усилий, но этот прием помогает пчеловоду наполнить бочки медом и дает большие доходы сельскому хозяйству.
Память на время у пчел
Каждый из нас по собственному опыту знает, что такое чувство времени. На него можно полагаться в большей или меньшей степени, но нет человека, у которого оно полностью отсутствовало бы. Собака или попугай хорошо запоминают час радостных для них событий, если эти события регулярно повторяются. Трудно строить предположения, может ли быть нечто подобное у насекомых, так как их организм устроен совсем иначе, чем наш. Решить это может только опыт. Устроим искусственный взяток на столике, установленном на воле, приманив туда пчел на чашку с сахарным сиропом. Насекомых, посещающих эту кормушку, пронумеруем, чтобы можно было узнавать «в лицо» каждую отдельную пчелу. Будем кормить пчел не в течение всего дня, а только определенное время, например с 4 до 6 часов после полудня. Так же поступим и в следующие дни. Разведчицы пронумерованной группы, которые посещают кормушку до 4 и после 6 часов пополудни, находят ее пустой и возвращаются домой ни с чем. Через несколько дней, в течение которых кормление производилось только с 4 до 6 часов дня, проводится контрольный опыт. На этот раз в кормушку совсем не наливается корма, чашка остается пустой также в период с 4 до 6 часов дня. С 6 часов утра до 8 часов вечера около нее неотлучно дежурит наблюдатель, который метит каждую пчелу, прилетевшую к чашке. Это очень скучная работа, потому что в нашем опыте из шести пчел, которые в предшествующие опыту дни посещали чашечку, за весь период с 6 часов утра до половины четвертого дня появилась только одна пчела № 11, чтобы проверить кормушку. Она прилетела между семью и половиной восьмого утра и вскоре появилась еще раз. В остальное время на кормушке царило абсолютное спокойствие. Однако, как только наступило время обычного кормления, дело пошло живее. За два часа между 4 и 6 часами дня у чашечки было зарегистрировано 38 посещений, сделанных пятью из шести помеченных пчел. Хотя их прилеты были напрасными, они через короткие промежутки времени снова возвращались и до десяти раз в течение получаса обследовали пустую кормушку, делая это так настойчиво, как будто они непременно должны были там что-то отыскать. К исходу обычных часов кормления, около шести часов вечера, посещаемость кормушки быстро снизилась, и вскоре на месте кормления снова стало спокойно. Вопреки ожиданиям опыт удался. Его результаты наглядно представлены на диаграмме (рис. 14). На нижней горизонтальной линии откладывается время с 6 часов утра до 8 часов вечера. Время дрессировки (с 4 до 6 часов в дни, предшествовавшие опыту) обведено рамкой. Весь отрезок времени разделен тонкими черточками на получасовые промежутки, над каждым из которых можно обнаружить обведенные квадратами опознавательные номера всех пчел, посещавших кормушку в данные полчаса, и подсчитать их число.
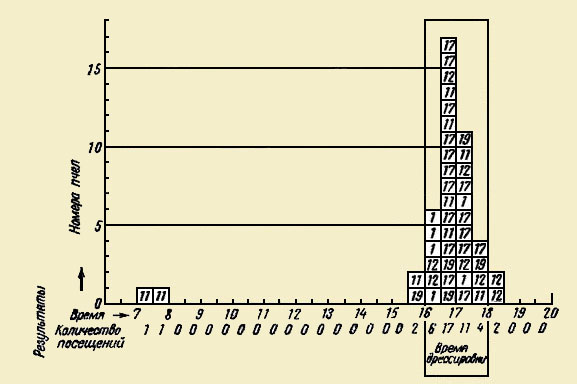 Рис.
14. Проверка способности пчел запоминать
время. Несколько пронумерованных пчел
ежедневно получали сахарный сироп на
кормушке с 16 до 18 часов. В день контрольного
опыта (20 июля 1927 года) кормушка целый
день — и в обычное для дрессировки время
— была пуста. На нижней горизонтальной
линии отложены часы дня. Над каждым
получасовым отрезком нанесены номера
пчел, которые прилетали в этот период
к кормушке. Помещенные друг над другом
квадраты с одинаковыми номерами означают,
что за этот период одна и та же пчела по
несколько раз прилетала из улья к
кормушке, чтобы произвести разведку.
(По Ингеборг Белинг.)
Рис.
14. Проверка способности пчел запоминать
время. Несколько пронумерованных пчел
ежедневно получали сахарный сироп на
кормушке с 16 до 18 часов. В день контрольного
опыта (20 июля 1927 года) кормушка целый
день — и в обычное для дрессировки время
— была пуста. На нижней горизонтальной
линии отложены часы дня. Над каждым
получасовым отрезком нанесены номера
пчел, которые прилетали в этот период
к кормушке. Помещенные друг над другом
квадраты с одинаковыми номерами означают,
что за этот период одна и та же пчела по
несколько раз прилетала из улья к
кормушке, чтобы произвести разведку.
(По Ингеборг Белинг.)
При помощи этого метода стаю возможным повышать только за счет дополнительного опыления пчелами урожай семян красного клевера и других сельскохозяйственных культур в 1,5 — 2 раза. Так как в результате дрессировки число пчел-сборщиц на цветках возрастает в 10 — 15 и более раз, оказалось возможным использовать на опылении значительно меньше пчелиных семей, чем без дрессировки. А. Ф. Губин, Н. П. Смарагдова и другие советские исследователи показали, что, применяя дрессировку, можно с равным успехом использовать для опыления красного клевера как более длиннохоботных кавказских, так и обычных среднерусских пчел. Об использовании пчел для опыления красного клевера подробно говорится в книге А. Ф. Губина «Медоносные пчелы и опыление красного клевера», Москва, 1947. — Прим. перев. Этот опыт неоднократно повторяли с другими пчелами и в разное время дня. Исход каждого из них не оставлял сомнения в том, что пчелы уже через несколько дней поразительно точно запоминают любой час кормления. Успешные результаты этих опытов побудили нас подвергнуть более трудному испытанию способность пчел чувствовать время. Результат нового опыта превзошел все ожидания. Удалась также дрессировка на два, три и даже на пять различных периодов кормления в течение дня. На рис. 15 приведен пример одной такой трехвременной дрессировки. Пунктуальность пчел в этом случае несколько пострадала: они прилетали к кормушке слишком рано. Такое явление часто наблюдается также и при дрессировке на одно время суток. И это не совсем нецелесообразно. Лучше прийти слишком рано, чем слишком поздно, так как в природе немало голодных ртов, готовых стянуть пищу из-под носа у других. Таким образом, пчел не так-то легко провести. И если вспомнить, что 13 августа с утра до вечера они не находили на опытном столике ни капли сахарного сиропа, то один взгляд на рис. 15 ясно покажет, что трехкратное кормление в определенные сроки на протяжении шести дней, предшествовавших опыту, не могло не оказать на них влияния.
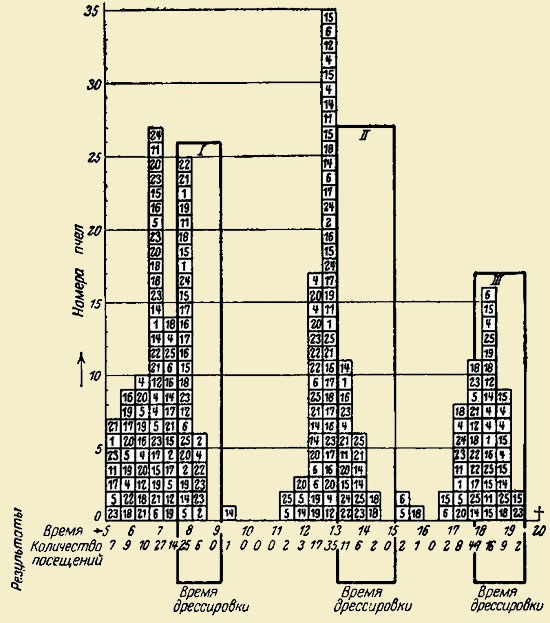 Рис.
15. Результат трехвременной дрессировки.
Продолжительность дрессировки — шесть
дней. В день контрольного опыта (13 августа
1928 года), несмотря на то что кормушки
весь день оставались пустыми, из 19
пронумерованных пчел прилетели все, и
именно в то время, которое отмечено на
рисунке. (По Ингеборг Белинг.)
Рис.
15. Результат трехвременной дрессировки.
Продолжительность дрессировки — шесть
дней. В день контрольного опыта (13 августа
1928 года), несмотря на то что кормушки
весь день оставались пустыми, из 19
пронумерованных пчел прилетели все, и
именно в то время, которое отмечено на
рисунке. (По Ингеборг Белинг.)
Возникает вполне естественный вопрос: где находятся у пчелы часы? Не в желудке ли, который в привычное для кормления время начинает пульсировать и понуждает ее вылетать из улья к кормушке? Этого не может быть уже потому, что время появления корма в кормушке — не время приема пищи в обычном смысле слова. Пчела летит к кормушке не для того, чтобы насытиться, а для того, чтобы собрать запас корма и отнести его в улей. Кроме того, она целый день находится на заполненных медом сотах и ей достаточно вытянуть хоботок, чтобы в любое время утолить голод. Это предположение полностью опровергается и следующим опытом: предложим группе пчел в течение многих дней с утра до вечера брать сахарный сироп, который в определенные часы дня предоставляется им в изобилии или который в эти часы более сладок, чем в остальное время. Пчелы собирают его без перерыва, их желудок никогда не остается пустым, и все же в дни наблюдений в привычное «наилучшее время сбора» они с огромным усердием прилетают к пустым теперь кормушкам. Но, может быть, пчела, подобно путешественнику, наблюдает за положением солнца? Этому, пожалуй, можно поверить, так как стало известно, что при определенных обстоятельствах пчелы очень внимательно следят за положением солнца на небе. Имеет ли положение солнца решающее значение для ориентировки пчел во времени, сами мы выяснить не можем и должны проверить это при помощи опыта. Можно перенести семью в темную комнату, чтобы район полета ее пчел был ограничен четырьмя стенами этого помещения. Если в этой «тюрьме» постоянно поддерживать температуру от 25 до 28° Цельсия, освещать ее при помощи электроламп и в изобилии снабжать пчел кормом на искусственной кормушке, то маленькая семейка даже в таких противоестественных условиях может здравствовать долгие годы. Она не ощущает смены времен года и выращивает в своих сотах расплод и летом и зимой. При равномерном освещении у пчел нет никакой возможности определять время по положению солнца или по степени освещенности. Несмотря на это, дрессировка на время удается и в таких условиях, если пчел в этом помещении кормить в определенные часы. Используя искусственное освещение, вероятно, можно распространить опыты также и на ночное время и в ночные часы кормления получать такие же результаты, как и днем. Опыт дрессировки пчел на больший, чем 24 часа, отрезок времени не удается. Можно неделями предлагать корм с промежутками в 19 часов (это не представляет никаких трудностей в темной комнате с равномерным освещением) — промежуток времени в 19 часов пчелы не воспринимают. В другом опыте пчел продолжительное время кормили каждые 48 часов. После этого в течение двух суток они прилетали за сиропом каждые 24 часа. Создается впечатление, что пчелы запоминают не отрезок времени между кормлениями, а определенное время дня, в течение которого им дают корм. Очевидно, есть две возможности: или пчелы руководствуются дневными периодическими воздействиями, недоступными нашему восприятию, или они имеют «внутренние часы», связанные с обменом веществ в их организме. Если это так, то все неудачи дрессировки на 19 или 48-часовой ритм можно объяснить только тем, что пчелы по своему образу жизни тесно связаны с суточным циклом и вследствие этого но переключаются на другую периодичность.
 Рис.
16. Трансокеанский опыт для проверки
способности пчел определять время
суток. Пчелы, дрессированные в Париже
на посещение кормушки в определенные
часы, были доставлены на самолете в
Нью-Йорк и там подвергнуты испытанию.
(По М. Реннеру.)
Рис.
16. Трансокеанский опыт для проверки
способности пчел определять время
суток. Пчелы, дрессированные в Париже
на посещение кормушки в определенные
часы, были доставлены на самолете в
Нью-Йорк и там подвергнуты испытанию.
(По М. Реннеру.)
Четкое разъяснение дал трансокеанский опыт. В Мюнхене сделали две совершенно одинаковые разборные темные камеры и одну из них переправили в Париж, а другую — в Нью-Йорк. Когда в Париже полдень (12 часов дня) и солнце стоит в зените, то у жителей Нью-Йорка еще только 7 часов утра и они видят утреннее солнце (рис. 16). Если пчелы руководствуются местным положением солнца, которое они воспринимают даже в темной камере благодаря проникающим туда лучам или другим неведомым нам образом, то после дрессировки на какое-то определенное время в Париже и быстрое перемещение через океан в Нью-Йорк они должны появиться на кормушке по нью-йоркскому времени. Но этого не происходит. Дрессированные в парижской темной камере на посещение кормушки в определенное время и перевезенные на самолете в Нью-Йорк, маленькие путешественницы и там прилетали к кормушке по парижскому времени. Итак, пчелы действительно в состоянии определять время суток по каким-то «внутренним часам». Очень выразительно демонстрируют они нам эту способность и другим образом. Пчелы-разведчицы семьи, находящейся в роевом состоянии, разыскав подходящее жилище, сообщают другим пчелам о его местонахождении при помощи танцев, причем направление к цели полета они указывают по положению солнца. Иногда они в течение многих часов остаются дома и, повторяя танцы, снова и снова обращают внимание пчел на возможность организовать новое гнездо. Находясь в улье, они не могут заметить, что солнце тем временем переместилось на небе. Несмотря на это, в своих продолжительных танцах они меняют угол по отношению к направлению силы тяжести в точном соответствии с тем, как изменяется за это время угол между направлением к объекту их полета и направлением к солнцу. Они делали это даже тогда, когда их в закрытом улье унесли в подвал, откуда они не могли видеть ни солнца, ни неба. Так пчелы дают нам новые доказательства наличия у них «внутренних часов» и одновременно согласованности их действий с суточным ритмом солнца. Способность чувствовать время необходима пчелам — в этом убеждает только что приведенный пример. Ведь и для общей ориентировки в пространстве солнце может быть использовано в качестве компаса только в том случае, если есть возможность принимать в расчет время суток. Так что и с этой точки зрения дрессировка пчел на кормушку, наполняемую в определенные часы сиропом, оказывается совсем не таким уж противоестественным опытом, как это может показаться некоторым. Многие цветы приглашают своих посетителей «к столу» также по часам. Они выделяют цветочную пыльцу и нектар лишь в определенное время суток. Часто выделение нектара хотя и продолжается целый день, но в некоторые часы происходит обильнее. У многих видов растений «лучшее время» приходится на утро, у других — на середину дня, а у некоторых — на послеполуденное время, и каждый вид растений обычно проявляет в этом отношении постоянство. А так как пчелы также в высшей степени постоянны в отношении посещения цветов, то есть каждая из них в течение дня посещает цветки только одного вида, это означает, что все группы сборщиц в известные часы очень напряженно работают, а остальное время бездействуют. Незанятые пчелы вынуждены оставаться дома, потому что внешний мир полон опасностей. Пчелы-сборщицы и в самом доле остаются в улье, когда источники, с которых они берут взяток, временно перестают его давать. Только отдельные разведчицы вылетают, чтобы проверить, не открылся ли взяток. Если такая разведчица обнаружит, что цветки снова наполнились нектаром, она танцует на сотах точно так же, как при впервые открытом новом источнике взятка, и снова призывает всю свою группу сборщиц на луг. При нерегулярных перерывах на взятке разведчицы летают более продолжительное время, чтобы не пропустить момента, когда их группа пчел сможет использовать заслуживающий внимания вновь открывшийся взяток. Если время взятка строго ограничено, пчелы-разведчицы быстро приспосабливают свои вылеты именно к этим часам дня. В безвзяточное время вся группа сборщиц обычно выбирается из беспокойной сутолоки «танцевальной площадки» и дремлет в каком-нибудь укромном уголке на краю сота, а их внутренние часы продолжают идти своим ходом. Когда наступает время взятка, все это общество, казалось бы заснувшее, пробуждается к жизни, как от звонка будильника. Пчелы медленно сползаются со всех сторон на участок сота, где происходят танцы и где они имеют возможность встретиться с первой разведчицей, возвратившейся домой с удачного полета. Иногда они сразу принимаются за работу по собственному инстинктивному побуждению.
Психические способности пчел
Этот раздел будет кратким, так как о том, что мало известно, не стоит много говорить. Но кое-что сказать все же необходимо. Узнав об остроумном устройстве сотов или о способности пчел поддаваться дрессировке, о их танцах и использовании ими небесного компаса, а также об энергичном избавлении от трутней в то время, когда они становятся ненужными семье, читатель, возможно, будет склонен приписывать пчелам незаурядные умственные способности. Однако это не так. Целесообразность какого-либо действия еще не доказывает, что это действие совершается обдуманно. Обычно мы считаем сознательным такое действие, когда какое-либо существо, используя уже накопленный жизненный опыт, правильно действует в новом, необычном для себя положении. А для этого необходима способность запоминать то, что происходило раньше, осмысливать создавшееся положение, связывая его с образами воспоминаний. Одна предпосылка для этого у пчел есть — хорошая память. Опыты с дрессировкой, о которых шла речь в предыдущих главах, вполне убеждают в этом. Если надрессировать пчел на какой-нибудь цвет, они даже после перерыва в несколько дней из-за плохой погоды ищут корм на бумаге того же цвета. Неделями, а иногда и до конца жизни пчелы помнят запах, на который их дрессировали короткое время, даже если потом им не случалось с ним встречаться. Не приходится сомневаться также и в том, что пчелы способны улавливать известную связь между явлениями. Если мы, например, давая пчелам корм в синей коробке, успешно дрессируем их на синий цвет, это служит доказательством того, что пчелы связывают синий цвет, который они видели, проникая в коробку, с найденным внутри кормом, что они «поняли» известную взаимосвязь между этими явлениями. Но нельзя переоценивать эту психическую способность пчел, так как она ограничена очень узкими рамками. Мне вспоминается здесь одна история об одиночной пчеле халикодоме. Это близкая родственница нашей медоносной пчелы, но она не относится к общественным насекомым. Халикодома строит для каждого яйца отдельную круглую ячейку из глины и наполняет ее медом до тех пор, пока запас не окажется достаточным для того, чтобы личинка, которая выведется из яйца, могла прокормиться им до полной зрелости. Собрав такой запас, пчела откладывает на него яйцо, запечатывает отверстие ячейки, чтобы уберечь свое потомство от врагов, и, не заботясь больше о его судьбе, принимается за постройку следующей ячейки. Один исследователь рассказывает о следующем опыте. Воспользовавшись временем, когда пчела-мать собирает взяток на цветах, он проделывает отверстие в только что построенной, но еще не заполненной кормом ячейке. Вернувшись, пчела замечает происшедшую перемену. Эго видно по тому, как она исследует усиками зияющее отверстие. Однако пчеле не приходит в голову заделать дыру, хотя это не составило бы для нее большого труда. Вместо этого она, как обычно, освобождается от кормовой ноши, которая вываливается через отверстие. Ношу за ношей вытряхивает она таким образом на землю после каждого полета за взятком. Следовало ожидать, что, заметив безуспешность своей работы, пчела будет приносить корм с еще большим усердием или бросит эту ячейку. Но ни того ни другого не происходит. Пчела приносит столько пищи, сколько необходимо для развития личинки при нормальных условиях. Затем она откладывает яйцо, которое сейчас же падает через проломанное дно на землю, и сверху заботливо запечатывает ячейку с зияющей снизу дырой. Я не знаю, верно ли это описание во всех подробностях. Если нет, то, во всяком случае, оно превосходно придумано и хорошо согласуется с многократными подобными же наблюдениями над другими насекомыми и медоносными пчелами. Я не смог бы привести из их жизни ни одного примера разумного действия. Даже упоминавшиеся выше опыты дрессировки не удавались, как только поставленная задача хоть немного отклонялась от привычных для пчел действий, выработавшихся в течение сотен тысячелетий. Сама природа, так сказать, вооружила их психической способностью связывать, например, цветочный запах с поиском корма. Но достаточно применить для дрессировки не цветочный, а, например, гнилостный запах, чтобы она совершенно не удалась. Гнилостный запах не кажется пчелам неприятным, так как они без малейшего колебания посещают кормушки в коробках с этим запахом. Соответствующим опытом можно доказать, что «пчелиный нос» воспринимает этот запах так же хорошо, как и цветочный. Однако предки наших подопытных пчел никогда не находили мед в какой бы то ни было связи с гнилостным запахом. А психические возможности одной пчелиной особи не позволяют ей уловить связь между этими двумя явлениями собственными силами. Таким образом, способность к обучению у пчелиных семей ограничена узкими рамками того, что для них имеет значение в естественных условиях и к чему они привыкли с древних времен. Пчела привыкает к синему цвету или запаху шиповника, если она находит возле них корм, точно так же, как бесчисленные поколения ее предков. Она унаследовала от них способность строить шестигранные восковые ячейки, формировать обножку, указывать при помощи танцев место хорошего взятка и зажаливать трутней, когда прошло их время. Со строгой правильностью привычные условия вызывают привычные действия. Сознают ли пчелы что-либо из того, что они делают? На этот вопрос никто не может ответить с уверенностью. Еще ни одному человеку не удалось также разгадать загадку, каким образом предки пчел приобрели способности, которые получили от них в наследство теперешние поколения.
Крушинская н.Л., Лисицына т.Ю. Характерные черты условнорефлекторной деятельности дельфинов
Печатается по изд.: Крушинская Н.Л., Лисицына Т.Ю. Поведение морских млекопитающих. М.: Наука, 1983. С. 127-166.
Экспериментальные данные, характеризующие условнорефлекторную деятельность дельфинов немногочисленны и в ряде случаев противоречивы. Первые публикации по этому вопросу принадлежат; Л. Г. Воронину и его ученикам (Воронин, Козаровицкий, 1969; Стародубцев, 1969; Воронин, 1970; Воронин и др., 1971, 1974). В работе Л. Г. Воронина, Л. Б. Козаровицкого (1969) приводится подробный анализ особенностей высшей нервной деятельности афалины при выработке системы пищедобывательных условных рефлексов на звуковой раздражитель. Проведенные исследования показали, что скорость образования положительных пищедобывательных условных рефлексов у дельфина не отличалась от таковой у других позвоночных животных. При выработке дифференцировок, переделки сигнального значения раздражителей, угашении положительных условных рефлексов выявились некоторые особенности их высшей нервной деятельности. Во-первых, авторами отмечается сильно выраженная ориентировочно-исследовательская деятельность у афалины, проявляющаяся в период становления условных реакций и при введении новых агентов в эксперимент. Во-вторых, дельфину была свойственна неуравновешенность нервных процессов с преобладанием возбуждения. Этим было обусловлено большое число межсигнальных реакций, часто носивших характер нападения на рычаг или агрессивных атак сигнализатора при неподкреплении условного ответа. В-третьих, у дельфина наряду с неуравновешенностью проявилась инертность нервных процессов, которая могла принимать черты патологической застойности (дельфин нажимал на педаль до 400 раз, не получая за это подкрепление). Инертностью нервных процессов авторы объясняют тот факт, что дельфины с трудом привыкают ко всему новому и отвыкают от старых привычек. В-четвертых, наряду с явно выраженной инертностью у дельфина была обнаружена хорошая концентрация их нервных процессов. Об этом свидетельствует устойчивость выработанных условных рефлексов в ситуациях, которые могли повлечь срыв высшей нервной деятельности (при применении сшибки раздражительного и тормозного процессов). Л.Г. Воронин и Л.Б. Козаровицкий отмечают склонность дельфина к развитию неврозоподобных состояний в период выработки торможения условнорефлекторных реакций. Так, период стабилизации во время выработки дифференцировки затянулся в связи с тем, что у афалины произошло нарушение условнорефлекторной деятельности: она отказывалась от корма, не подходила к установке, «зависая» в углу бассейна. Эти периоды заторможенности у нее сменялись бурными вспышками двигательной активности: она на большой скорости плавала по бассейну, била хвостом по воде, атаковала сигнализатор, рвала зубами паралоновую обшивку на рычаге и т. д. Такое невротическое состояние у дельфина могло быть достаточно устойчивым. К этому следует добавить, что многие исследователи (McBride, Hebb, 1948; Вуд, 1979; Прайор, 1980) отмечают у дельфинов проявление бурных эмоциональных вспышек во время проведения экспериментов. Так, афалина совершив ошибку во время опыта сломала установку и впала в затяжное агрессивное возбуждение (Morgan, цит. по: Воронин, 1970). Ф. Вуд (1979) сообщает, что однажды во время работы связным в открытом море известный дельфин по кличке Таффи, не получив положенного вознаграждения за работу, ударил экспериментатора. В наших экспериментах афалина Дик после каждого неправильного выбора цели била хвостом плавающего рядом ската. Приведенные примеры показывают, что, работая с дельфинами, всегда следует учитывать их эмоциональное состояние, проявление которого может неблагоприятно отражаться на ходе эксперимента. Исследование динамики образования различных видов пищедобывательных условных рефлексов на звуковые раздражители было проведено В. А. Протасовым и др. (1976) на большой группе афалин (15 животных). Образование условнорефлекторных подходов по сигналу к месту подкрепления у дельфинов в среднем происходило после 3 — 10 предъявлений, стабилизация условного рефлекса — после 20 — 60 применений условного сигнала. Образование инструментального условного рефлекса (нажатие рострумом на рычаг) потребовало более длительной тренировки для укрепления рефлекса (137 — 168 сочетаний). Угашение выработанных условных рефлексов наступало после 10 — 30 применений сигнала без подкрепления. Угашение было нестойким и восстанавливалось в том же опыте. Проведенные на двух афалинах эксперименты показали, что у них легко и быстро, как и у большинства высших наземных животных образуются тормозные дифференцировки, условный тормоз первого и второго порядка, рефлексы второго порядка (Протасов и др., 1975). В экспериментах (Протасов и др., 1976) была показана возможность образования у дельфинов динамического стереотипа при частичном подкреплении последовательных применений условных раздражителей. У афалины был сформирован стереотип подхода на каждое второе и каждое третье применение условного сигнала. Подход на каждое четвертое применение сформировать не удалось, так как дельфин перестал покидать стартовую площадку при применении сигнала. По данным В.А. Протасова и др. (1975), медленно и трудно у афалин образуются рефлексы с дифференцированием нескольких положительных условных раздражителей и рефлексы с одновременным предъявлением положительного и тормозного условного раздражителей, но легко и быстро, как и у большинства высших наземных животных, у дельфинов образуются дифференцировки и переделки условнорефлекторных ответов при последовательном предъявлении тормозных и положительных раздражителей. На основании анализа результатов экспериментов, проведенных на большой группе афалин, авторы (Протасов, Сергеев, 1975; Протасов и др., 1976) заключают, что по скорости формирования простых пищедобывательных рефлексов дельфины не отличаются от других позвоночных животных, что подтверждает выводы, сделанные Л.Г. Ворониным, Л.Б. Козаровицким (1969). Обсуждая полученные данные, В.А. Протасов и др. (1976) выделяют некоторые особенности поведения дельфинов при образовании у них условных рефлексов. Они заключают, что для поведения дельфинов в неволе характерны слабая выраженность ориентировочно-исследовательской деятельности и склонность к развитию автоматизмов. Наличие автоматизированных реакций отмечалось авторами в начальный период обучения, когда животные, слабо реагируя на предъявленные сигналы, монотонно подходили к манипулятору через определенные интервалы времени или реагировали только на один из манипуляторов, независимо от сигнального значения предъявляемого раздражителя; в случае альтернативного выбора у них развивался «односторонний автоматизм». Предрасположенность дельфинов к автоматизированным реакциям, как полагают авторы, закреплена экологически и связана со спецификой охоты на косяки рыбы, при которой обеспечивается «вероятностное подкрепление». Как известно (Купалов и др., 1964; Фирсов, 1972; Крушинский, 1977), односторонний автоматизм реакций возникает не только у дельфинов, но и у других животных в период выработки рефлексов, когда животное еще не выделило из комплекса признаков условного сигнала существенный, по которому происходит обучение. Односторонний автоматизм также отмечается у разных животных при перенапряжении нервной системы во время решения трудной задачи с альтернативным выбором (Крушинский и др., 1972; Крушинский, 1977). На основании собственных исследований с дельфинами и данных других авторов мы полагаем, что автоматизм реакций у дельфинов не следует рассматривать, как отличительный, признак их поведения, поскольку он возникает функционально как реакция на трудные ситуации. Выводы В. А. Протасова и др. (1976) о слабовыраженном у дельфинов ориентировочно-исследовательском рефлексе явно вступают в противоречие с мнением большинства исследователей, изучавших поведение афалин (McBride, Hebb, 1948; Tavolga, 1966; Воронин, Козаровицкий, 1969; Caldwell, Caldwell, 1972a, b; Вуд, 1979; и многие др.). Мнение о слабовыраженном у дельфинов ориентировочно-исследовательском рефлексе могло сложиться у экспериментаторов (Протасов и др., 1976) при проведении опытов в период адаптации животных к условиям неволи. В этот период, по нашим наблюдениям, дельфины обычно плавают тесной группой, не реагируют на внешние раздражители и избегают контактов с незнакомыми объектами. Период адаптации у дельфинов продолжается разное время (Caldwell, Caldwell, 1972b) и зависит от того, какие взаимоотношения у них складываются с другими обитателями бассейна. Внутригрупповые отношения могут существенно влиять на их работу при проведении экспериментов и дрессировке (Pryor, 1975). Р. Дефран, К. Прайор (Defran, Pryor, 1981) приводят данные, свидетельствующие, что афалинам в равной степени свойственны как осторожность (пассивно-оборонительный рефлекс), так и исследовательская реакция на новые объекты и ситуации. В связи с этим можно предположить, что отсутствие реакции у дельфинов на новые объекты в бассейне обусловлено временным подавлением ориентировочно-исследовательского рефлекса на новую обстановку пассивно-оборонительной реакцией. Таким образом, при анализе результатов исследований, касающихся возможности обучения дельфинов, следует учитывать индивидуальные особенности психического склада подопытного животного. К ним относятся уровень эмоциональности, склонность к установлению контактов с человеком, степень выраженности пассивно-оборонительной реакции и т. д. В связи с этим следует привести один интересный пример из практики обучения афалины по кличке Василиса. Этот дельфин отказывался участвовать в опытах, цель которых состояла в обучении животных различать звуковые сигналы заданных параметров по стандартной пищедобывательной методике. Любые попытки в течение длительного времени вовлечь ее в работу оказались тщетными. Во время опытов с другими дельфинами самка «зависала» в углу бассейна. Вместе с тем в другой форме эксперимента, построенного на игровой активности, Василиса проявила большую заинтересованность в работе и продемонстрировала способность к решению простых логических задач (Крушинский и др., 1972). Следует отметить, что в дальнейшем афалина Василиса на протяжении многих лет была одной из лучших цирковых артисток Батумского океанариума (Томилин, 1980). Приведенный пример наглядно демонстрирует, что дельфины могут проявлять индивидуальную склонность, определяемую свойствами их психического склада, к тем или иным экспериментам, которые им предлагает человек в условиях неволи. Остановимся несколько подробнее на разборе экспериментальных данных, полученных при исследовании формирования цепных условных рефлексов и выработке сложных видов дифференцировок у дельфинов. В своей повседневной жизни дельфинам приходится реагировать не только на одиночные раздражители окружающей их среды, но чаще на целые их комплексы. Дельфины живут в мире звуков, многие из которых являются коммуникационными сигналами, связанными в определенной последовательности. Исследованию становления условнорефлекторных связей дельфинов на цепной акустический раздражитель посвящена работа Л. Б. Козаровицкого и др. (1971). Эксперименты были проведены на самке афалине по пищедобывательной условно-рефлекторной методике. В качестве условного сигнала применялся акустический цепной раздражитель, состоящий из трех тонов: А — 12, Б — 6 и В — 3,2 кГц. Каждый член цепи действовал изолированно 2 с, а весь цепной раздражитель — 6 с. Реакция дельфина на рычаг при действии условного сигнала поощрялась пищей. Полученные данные показали, что у афалины легко вырабатывается условный рефлекс на цепной раздражитель. Этот рефлекс укрепляется так же быстро, как и рефлексы на одиночные раздражители. В процессе выработки условного рефлекса на акустический цепной раздражитель у дельфина наблюдалось развитие запаздывательного торможения. Такого рода закономерность наблюдалась другими авторами при выработке условного рефлекса на цепи раздражителей у обезьян, собак, кроликов, голубей и рыб (Воронин, 1947; Ширкова, 1949; Овчинникова, 1955; Бару, 1954; и др.). Вследствие развития запаздывательного торможения по мере укрепления рефлекса ответные двигательные реакции у животных отодвигались к концу цепи, т. е. к моменту пищевого подкрепления. Проведенные Л.Б. Козаровицким и др. (1971) эксперименты показали, что становление условного рефлекса на цепной раздражитель у дельфинов происходит так же, как и у других позвоночных животных. Особенность образования цепных пищедобывательных условных рефлексов у афалины исследовалась Ю.Д. Стародубцевым (1969). Дельфин был обучен последовательно нажимать на 8 педалей, за что получал пищевое вознаграждение. Затем ему был предложен более сложный вариант задачи: нажимать не на каждую педаль, а только в определенной последовательности (на 2-ю, 5-ю и на 7-ю педали). При таком режиме подкрепления у дельфина не удалось сформировать последовательную цепь движений. Дельфин, не получая пищи при нажатии на любую педаль, продолжал многократно на нее воздействовать разными способами (сверху, снизу, сбоку), разными частями тела (рострумом, лбом, грудными и хвостовым плавниками). Сходная задача, предложенная обезьянам, не вызывала у них затруднений. При формировании цепи движений на определенную последовательность воздействия на рычаги обезьяны, не получив подкрепления при воздействии на одну педаль, сразу переходили к другой, а не продолжали упорно манипулировать с одной, как это делал дельфин. Такая застойность реакции у дельфинов объясняется Л.Г. Ворониным (1970) инертностью их нервных процессов. Сложный вид навыка, образованного по принципу цепного рефлекса, удалось выработать у афалины Дж. Лилли (Lilly, 1967). Цепь этих навыков была следующей: дельфин нажимал на кнопку — вызов лифта, затем заплывал в расположенную там ванну и, нажимая рострумом на соответствующую педаль, поднимался к поверхности воды, либо в здание, откуда его переводили в бассейн. Вместе с тем следует отметить, что не менее сложные цепи условнорефлекторных движений удается сформировать и у крыс. Р. Пиррел и Дж. Шерман (Pierrel, Sherman, 1963) обучали крыс производить ряд последовательных действий: взбираться по спиральной лестнице, опускать разводной мостик и перебираться по нему; влезать по веревке и, опустившись на подъемнике на площадку, нажимать на рычаг, после чего они получали пищу. Такие сложные инструментальные цепи формируются в результате подкрепления отдельных движений, последовательное выполнение которых приводит к образованию столь сложного навыка. Таким образом, приведенные выше данные показывают, что формирование цепных двигательных условных реакций у дельфинов происходит так же, как и у других видов млекопитающих. Обнаруживающаяся некоторая специфика поведенческих реакций дельфинов в процессе обучения может определяться особенностью протекания их нервных процессов. Общение дельфинов со своими сородичами осуществляется посредством многообразных и сложных коммуникационных звуков: зрительных, тактильных и химических сигналов. С помощью этих сигналов дельфины узнают животных, принадлежащих к своему сообществу, отдельных индивидуумов в группе, определяют их положение на иерархической лестнице, выбирают полового партнера, узнают о том, что сородич нуждается в помощи, предупреждают о своих агрессивных намерениях и т. д. Из сказанного становится ясно, что дельфины в своей повседневной жизни должны уметь дифференцировать огромное число сигналов, адресующихся к разным анализаторам. М. Колдуэлл и Д. Колдуэлл (Caldwell, Caldwell, 1968a, 1972а, с) в исследованиях по изучению коммуникационных сигналов у дельфинов на большом материале показали, что каждый дельфин обладает индивидуальным, только ему присущим характером свиста. Для выяснения вопроса, способны ли дельфины различать индивидуальные свисты других животных, был проведен специальный эксперимент (Caldwell, Caldwell, 1972а). Самец афалина должен был нажимать рострумом на рычаг только при сигналах, издаваемых определенными особями. На магнитофонную ленту были записаны свисты нескольких дельфинов, издаваемых в разных ситуациях. Эту запись переписали на другую ленту, включив туда свисты еще нескольких дельфинов. Подопытный дельфин мог безошибочно выделять свист одного индивидуума из восьми свистов других животных, записанных на магнитофонную ленту. Причем из множества сигналов, подаваемых восемью дельфинами, он мог отличать разные типы свистов, принадлежащие только четырем определенным дельфинам. Этот эксперимент показал, что дельфины могут узнавать своих сородичей по их индивидуальным свистам, издаваемым в различных ситуациях. Экспериментальное исследование по выработке дифференцирования акустических сигналов было проведено (Herman, Arbeit, 1973; Herman, 1980) на самке афалине. Акустические сигналы предъявлялись последовательно из двух динамиков. Положительный сигнал подавался в случайном порядке из левого или правого динамика; реакцию дельфина на педаль подкрепляли только в ответ на действие положительного стимула. Новую пару сигналов вводили после достижения дельфином определенного критерия при распознавании предыдущей пары звуков. Из 25 пар звуковых сигналов дельфин правильно дифференцировал 23 пары. Переделка дифференцирования звуковых сигналов (изменение положительного стимула) также успешно осуществлялась дельфином. При этом обнаруживалась явная тенденция к улучшению решения этих задач при их повторении. Это выражалось в уменьшении числа проб, необходимых для достижения критерия в каждой новой задаче. По мнению авторов, дельфины усваивают «концепцию» проводимого с ними эксперимента и могут справляться с задачей с первых ее применений. Ускорение процесса обучения по мере увеличения последовательных переделок при различении зрительных стимулов наблюдается и у других животных: горилл (Rumbaugh, Steinmetz, 1971), крыс и голубей, но не достигается лишь у рыб (Bitterman, 1965). Переделка дифференцировок рассматривается М. Биттерманом как один из показателей качественного различия способности к обучению у разных видов позвоночных. Однако вопрос остается спорным, поскольку по этому признаку не удается найти качественные различия в познавательных способностях крыс и таких высокоорганизованных животных, как обезьяны и дельфины. Изучение способности дельфинов различать геометрические фигуры с помощью сонара проводились главным образом путем выработки у дельфинов инструментальных пищедобывательных условных рефлексов. Исследования показали, что дельфины с помощью отраженных эхосигналов могут различать лоцируемые объекты по их размерам (Morris et al., 1966; Дубровский, Краснов, 1971), форме (Белькович и Борисов, 1971) и материалу (Evans, Powell, 1967; Дубровский и др., 1970). Мы не ставим своей задачей изложение данных, характеризующих разрешающие возможности сонара дельфина. Этому вопросу посвящены специальные обобщающие работы (Яблоков и др., 1972; Айрапетянц, Константинов, 1970; Белькович, Дубровский, 1976). Данные по эхолокационному различению дельфинами объектов под водой мы будем приводить только в целях выяснения их познавательных возможностей. Одно из первых и наиболее обстоятельных исследований способности дельфинов к дифференцированию зрительных стимулов было проведено В. Келлогом, Ш. Райсом (Kellogg, Rice, 1966) на самке афалине. Дельфину было предъявлено 31 изображение разных геометрических фигур, из которых составлялись пары для зрительного их различения. Дифференцируемые фигуры предъявлялись одновременно в двух соседних прорезях окон специально сконструированного аппарата. При выборе положительного стимула дельфин дотрагивался носом до изображения соответствующей фигуры и получал подкрепление. Зрительные стимулы предъявлялись как под водой, так и над водой. Перед началом опыта, который проводился под водой, дельфин занимал стартовую позицию перед пластиковым подвижным экраном, раздвижение которого служило сигналом начала опыта. При одновременном предъявлении дифференцируемых раздражителей более сильный фактор — пространственное расположение фигур — обычно маскирует более слабый признак — форму предметов. Для того чтобы упростить задачу различения предъявляемых зрительных стимулов, В. Келлог и Ш. Раис провели предварительную серию опытов. Отрицательный стимул, предъявляемый в паре с положительной фигурой, имел дополнительный контрастирующий признак — большую освещенность и светлую латунную рамку вокруг изображения фигуры (положительная фигура была изображена на сплошном черном фоне). По мере тренировки освещенность целей выравнивалась и рамку у отрицательной фигуры убирали. Результаты экспериментов показали, что дельфин быстро усвоил задачу, и из 25 пар стимулов, которые предъявлялись под водой, правильно дифференцировал 21 пару (84%). В целях выяснения возможностей дельфинов к переносу опыта было выбрано 12 пар фигур, которые дельфин различал в результате обучения; 10 пар других комбинаций тех же фигур предъявлялись дельфину в качестве новых тестовых задач для различения. В тестовых парах обе фигуры, только положительную или отрицательную, поворачивали на 90 или на 180°, а в ряде опытов заменяли на другие (рис. 1). Как следует из рис. 28, дельфин выбирал положительную фигуру независимо от угла ее поворота и сочетания в паре с другой отрицательной фигурой, а также при замене положительного треугольника на круг. Результаты проведенных экспериментов демонстрируют способность дельфинов к некоторым формам обобщений, сопоставимым, по мнению авторов, с таковыми у обезьян.
 Рис.
1. Примеры изображения фигур, предъявлявшихся
дельфину для исследования его способности
к зрительному распознаванию
а — в
тестовых парах изменялась отрицательная
фигура; б — изменялась положительная
фигура; в — изменялись обе фигуры; г —
задача, не решенная дельфином (по:
Kellogg, Rice, 1966)
Рис.
1. Примеры изображения фигур, предъявлявшихся
дельфину для исследования его способности
к зрительному распознаванию
а — в
тестовых парах изменялась отрицательная
фигура; б — изменялась положительная
фигура; в — изменялись обе фигуры; г —
задача, не решенная дельфином (по:
Kellogg, Rice, 1966)
В той серии опытов, где дельфину фигуры для различения были предложены на воздухе, он не смог обучиться различать их из-под воды даже после 100 применений. Авторы считают, что это обусловлено искажением изображений фигур вследствие рефракции. Однако после того как животное было обучено различать фигуры в воде, оно стало их дифференцировать и на воздухе. Можно думать, что сложности, возникшие с различением дельфином фигур, предъявляемых на воздухе, были обусловлены их относительно небольшими размерами (4—5 см2), близкими к порогу различения. По наблюдениям самих авторов и опытам других исследователей (Adler, Adler, 1978; Близнюк, 1978; Прайор, 1981), дельфины могут различать геометрические фигуры больших размеров (30X50 см), предъявленные им на воздухе, и дифференцировать большое число жестов тренера, которые служат им сигналами для выполнения определенных действий. Исследования способности дельфинов различать зрительные стимулы под водой были проведены также Л. Германом и др. (Herman et al., 1969) на одной самке афалине. Различные зрительные стимулы дву- и трехмерные геометрические фигуры, различающиеся по форме и размерам; контрастные фигуры — белый круг на черном фоне и черный круг на белом фоне) одновременно предъявлялись дельфину для различения. Все фигуры демонстрировались за стеклянным окном в целях исключения возможности использования дельфином сонара. Как заключают авторы, образование навыка дифференцирования данных стимулов происходило с трудом и число правильных ответов не превышало 60%. Обсуждая исследования В. Келлога и Ш. Раиса, Л. Герман (Herman, 1980) высказывает сомнение в том, что в их опытах была надежно исключена возможность эхолокационного различения предъявляемых стимулов. Так, по его мнению, особенности рельефа самих латунных пластин, на которые наносился трафаретный рисунок фигур, могли служить для дельфина источником отраженных эхосигналов. Приводимые Л. Германом аргументы не кажутся нам достаточно убедительными для того, чтобы опровергнуть трактовку опытов В. Келлога и Ш. Раиса как обучение дельфина зрительному различению фигур. Несмотря на некоторые методические погрешности, работа В. Келлога и Ш. Раиса остается одним из наиболее интересно задуманных и тщательно выполненных исследований способности дельфинов к сложным видам обучения дифференцированию геометрических фигур и переносу навыка для выявления отношений между стимулами. К группе сложных дифференцировок относится так называемая методика «выбор по образцу». В этой задаче животное должно выбрать из нескольких объектов один, соответствующий сигналу-образцу, на который у него выработан условнорефлекторный ответ. В опытах на обезьянах и дельфинах применяют два варианта этой задачи: выбор по образцу наличного раздражителя и оставленного во времени раздражителя, т.е. в последнем случае животное производит выбор по памяти. Впервые эти эксперименты были проведены на шимпанзе Н. Н. Ладыгиной-Котс (1923) в целях выяснения функций обобщения у обезьян. В этих опытах было выяснено, что для шимпанзе большее значение имеет признак цвета и меньшее — величины и формы. В дальнейшем различные варианты «выбора по образцу» применялись многими исследователями на разных видах обезьян (Маркова, 1967; Фирсов, 1972; и др.) и продемонстрировали большие возможности этих животных к обобщению. Исследование наличного выбора по тождеству сигналов было проведено в нескольких работах на афалинах (Антоненко, Близнюк, 1978; Протасов и др., 1976; Adler, Adler, 1982). В. В. Антоненко, Я. И. Близнюк (1978) провели подобные исследования на двух афалинах. Одной предлагалось выбрать из разных по форме, величине и материалу объемных фигур, находящихся под водой, образец, сходный только по форме. Второму дельфину предлагалась похожая задача, но фигуры были плоскими и предъявлялись над водой. При возможности одновременного использования зрительного и эхолокационного анализаторов у первого дельфина был относительно быстро выработан правильный ответ при дифференцировке формы предметов. В тех случаях, когда дельфинам приходилось решать задачу выбора по образцу, пользуясь только зрительным анализатором (выбор над водой) или только эхолокационным (при очень мутной воде), выработка такой дифференцировки происходила с большим трудом. Авторы заключают, что обучение дельфинов сложным дифференцировкам типа «выбора по образцу» является для них сложной задачей. К такому же выводу пришли В.А. Протасов и др. (1976), исследуя у двух афалин возможность решения задачи «выбора по образцу» объемных фигур, предъявляемых дельфинам для локационного различения под водой. Отмечается невысокий уровень адекватных ответов у дельфинов (69%) при решении этой задачи. Как следует из этих опытов, существенным облегчением решения задачи выбора по тождеству предметов для дельфинов была возможность одновременно использовать зрительный и слуховой анализаторы. О взаимодействии зрительного и слухового анализаторов у дельфинов при различении целей свидетельствуют специально проведенные (Багдонас и др., 1970) эксперименты на обыкновенном дельфине при выработке дифференцирования двух геометрических фигур, различающихся по оптическим и акустическим свойствам и предъявляемых в разных условиях видимости. Было выяснено, что в ухудшенных условиях для функционирования эхолокационного анализатора (применение акустически прозрачных плексигласовых целей) дельфин преимущественно пользуется зрением при различении фигур. В условиях, затрудняющих зрительное различение (повышенная мутность воды до 0,5 м), дельфин включал эхолокационный анализатор. Приведенные данные дают основание считать, что при распознавании объектов под водой зрительный и локационный анализаторы у дельфинов тесно взаимосвязаны и дополняют друг друга. Другим вариантом задачи выбора по образцу можно рассматривать «смену принципа выбора». Эта задача была предложена двум афалинам (Adler, Adler, 1982). У дельфинов вырабатывался условный положительный рефлекс на белый квадрат определенного размера, предъявляемый над водой, а затем им предлагалось выбрать этот положительный стимул из четырех других квадратов. В первом варианте опыта они выбирали его среди четырех белых квадратов разного размера, во втором — из четырех квадратов такого же размера, но разных оттенков. Оба дельфина с высокой степенью надежности справились с обоими вариантами этой задачи. Авторы полагают, что этот эксперимент может демонстрировать достаточно высокий уровень познавательных способностей дельфинов афалин. Исследование возможности дельфинов решать задачи отсроченного выбора по образцу акустических сигналов было проведено в большой серии опытов (Herman, Gordon, 1975; Herman, 1975, 1980). Авторами было показано, что дельфины легко справляются с этой задачей и могут производить выбор по тождеству сигналов с паузой до 8 — 10 с. Подробное изложение этих экспериментов приводится в разделе «Характеристика памяти дельфинов». Приведенные выше данные свидетельствуют о том, что дельфины могут проявлять высокий уровень способности к выработке тонкого дифференцирования зрительных и слуховых сигналов и при опознавании объектов под водой используют как зрительный, так и эхолокационный анализаторы, функционирование которых дополняют друг друга при ориентации животного. Решение дельфинами сложных задач по дифференцированию типа «выбора по образцу» и вычленение положительной фигуры при пробах на инвариантность (изменение ее положения, Kellogg, Rice, 1966) показывают, что афалины, как и обезьяны, способны к обобщениям. Такие формы обобщений Л.А. Фирсов (1972) склонен рассматривать как «модель элементарной абстракции», близкую к интеллектуальной операции. При обучении дельфинов в лаборатории и при дрессировке для цирковых представлений используют так называемое положительное подкрепление (пищевое, игровое, тактильное — прикосновение, ласка и т.д.). Методы наказания или отрицательное подкрепление (удары электрическим током, погружение в холодную воду, действие сильных звуковых раздражителей и т.д.) обычно при работе с дельфинами не применяются. Любое неприятное воздействие со стороны человека, в том числе наказание, надолго нарушает контакт с подопытным животным, что происходит, по-видимому, в силу их повышенной эмоциональности. Вместе с тем в жизни дельфинам приходится, как и всякому животному, сталкиваться с воздействием неприятных, вредящих факторов, на основе которых дельфины с детства обучаются избегать подобные ситуации. Так, для поддержания иерархического порядка дельфины используют угрожающие звуковые сигналы, позы и прямое нападение, укусы, удары хвостом, а провинившихся детенышей родители прижимают на некоторое время ко дну. Трансляция в воду записей голоса косатки оказывала неизменно паническое действие на стада белух. Этим приемом, как отмечает Ф. Вуд (1979), пользуются рыбаки для отпугивания стад белух от устья р. Квичак во время нереста семги. А. Марчисон, Р. Пеппер (Murchison, Pepper, 1972) провели экспериментальное исследование эффективности отпугивающих стимулов на выработку двигательного условного рефлекса у афалин. В качестве «отрицательного подкрепления» за неправильное действие применялось сильное звуковое воздействие (акустический сигнал 50 кГц, создававший давление 63 дБ). По поведению дельфина можно было установить, что применявшийся звук неприятен для животного. Дельфина помещали в специальную подводную камеру (4,8X10,8 м), разделенную на две половины невысоким барьером. При включении акустического сигнала дельфин должен был из стартовой камеры переплыть в «безопасную зону», которая находилась ближе к неприятному источнику звука, после чего звук выключался. У дельфина был выработан условный рефлекс приближаться к подводному источнику неприятного звукового воздействия для выключения этого звука. В течение первых 100 предъявлений раздражителя, действовавшего каждый раз в течение 60 с., дельфин переплывал в безопасную зону в 44% случаев, а при последующих 100 предъявлениях он избегал неприятный раздражитель в 100% случаев. Этот эксперимент подтверждает возможность обучения дельфинов в лабораторных условиях по методике выработки оборонительного рефлекса. Итак, исследования формирования условнорефлекторных связей у дельфинов показали, что по скорости образования положительных пищедобывательных условных рефлексов они не отличаются от других позвоночных животных. У них вырабатываются положительные пищедобывательные рефлексы, все виды внутреннего торможения, наблюдается самовосстановление угашенных реакций, образуются условный рефлекс и условный тормоз второго порядка. Легко и быстро образуются тормозные дифференцировки, переделка условных рефлексов. Трудно и медленно вырабатываются, по мнению одних авторов (Протасов и др., 1976), рефлексы с дифференцированием нескольких положительных условных раздражителей и рефлексы с одновременным предъявлением положительного и тормозного условных раздражителей. По данным других авторов (Kellogg, Rice, 1966), выработка тонких дифференцировок зрительных сигналов с одновременным предъявлением положительных и тормозных сигналов не вызывает серьезных затруднений у дельфинов при соответствующей постановке опытов. По основным видам условнорефлекторных связей — положительным условным рефлексам, некоторым видам дифференцировок, условным рефлексам на цепи раздражителей, формированию цепных условнорефлекторных движений, переделке сигнального значения раздражителей выработке активного избегания — дельфины существенно не отличаются от других высших наземных животных. Предъявление более сложных задач по дифференцированию сигналов типа «выбора по образцу» при наличном и следовом сопоставлении раздражителей выявило у дельфинов способность к сложным формам общения. По этим показателям поведения дельфины могут быть сопоставлены с обезьянами. В период формирования условнорефлекторных связей были выявлены некоторые особенности их высшей нервной деятельности. Исследователи обращают внимание на значительную инертность нервных процессов дельфинов, сочетающуюся с их хорошей концентрацией. Эти особенности обусловливают большую устойчивость временных связей у дельфинов к действию различного рода дополнительных раздражителей, но, с другой стороны, затрудняют выработку некоторых временных связей, требующих быстрого переключения нервных процессов. Одним из отличительных признаков у афалин многими авторами отмечается значительная неустойчивость нервных процессов с преобладанием возбуждения, выражающаяся в легком возникновении неврозоподобных состояний. Возникающие у некоторых авторов разногласия при сопоставлении результатов опытов по обучению дельфинов в большей мере обусловлены тем, что эксперимент обычно проводится на одной или двух афалинах, обладающих, как известно, индивидуальными особенностями поведения, которые определяются уровнем эмоциональности, степенью выраженности ориентировочно-исследовательского рефлекса, пассивно-оборонительной реакции и т. д. Эти особенности индивидуального проявления высшей нервной деятельности дельфинов должны учитываться при анализе результатов исследований их поведения. В заключение необходимо отметить, что изучение высшей нервной деятельности дельфинов требует долгих планомерных исследований с учетом индивидуального подхода к этим высокоорганизованным животным.
Имитационное поведение
Одним из наиболее интересных и биологически важных видов группового поведения является подражание, или имитация. Имитационное поведение широко распространено в животном мире и играет существенную роль в приспособлении отдельных особей и вида в целом к меняющимся условиям среды. У позвоночных животных подражание является мощным цементирующим фактором в стадной жизни животных и имеет большое биологическое значение, поскольку служит основой накопления индивидуального опыта. Л.А. Орбели (1949) указывал на большое эволюционное значение имитационного поведения, считая его «главным охранителем вида». Исследователями выделяются две основные категории подражания у животных: стимулирование видотипичных действий и научение путем подражания или истинное имитационное поведение (Andrew, 1962; Фабри, 1974). Взаимное стимулирование видотипичных, врожденных действий широко распространено в животном мире как элемент группового поведения. В этологической литературе взаимное стимулирование животных обозначается как аллеломиметрическое поведение. Аллеломиметрическое поведение проявляется в наиболее простой форме — рефлексе следования. Оно обнаруживается у стайных рыб, птиц, копытных, хищных млекопитающих, приматов. Одновременный отдых, собирание пищи, бегство или комфортное поведение животных также являются примерами аллеломиметрического поведения. Результатом этих действий является согласованность, унификация и синхронизация поведения членов стада, что обеспечивает безотлагательное бегство всех животных стада в опасных ситуациях. Подлинное подражание, или «имитационное научение» (Штодин, 1947; Кряжев, 1955), отличается от аллеломиметрического поведения тем, что животное обучается перенимать те формы поведения, которые наблюдает у другой особи своего или другого вида. Имитационное научение проявляется в виде подражания тем формам поведения, которые не входят в репертуар врожденной видовой деятельности наблюдателя. Случаи истинной имитации известны для птиц, когда они подражают песне другого вида или имитируют человеческий голос. Примеры обучения путем наблюдения известны у рыб (Козаровицкий, 1961), у крыс (del Russo, 1975), кошек (John et al., 1968), но наивысшими способностями в этом отношении обладают обезьяны (Воронин, 1947; Кряжев, 1955; Фабри, 1965; Riopelle, Hill, 1973; и др.). Формирование и проявление различных видов подражательного поведения в сообществах животных находятся в непосредственной связи с иерархической организацией взаимоотношений членов данной группы. Влияние доминирующих особей в большой степени отражается на проявлениях подражания в сообществах у обезьян (Menzel, 1971; Воронин, 1947; Фабри, 1965). Так, опыты Е. Мензеля (Menzel, 1971) с шимпанзе показали, что группа обезьян, как правило, следует за лидером, которому известно, где находится тайник с лакомствами, и обычно игнорирует попытки низкорангового животного увлечь за собой сородичей к известному ему источнику пищи. Таким образом, подражание широко распространено в животном мире и играет существенную роль в групповых отношениях разных видов. Вместе с тем наивысшего развития имитационное поведение достигает у приматов и китообразных. Стадная жизнь большинства видов зубатых китообразных связана с развитием у них разных форм подражательного поведения. Многие примеры проявления стадного инстинкта или реакции следования наблюдаются у китообразных в природе. Одно из самых загадочных явлений в жизни китообразных — групповое самовыбрасывание животных на берег — рассматривается (Norris, 1967; Томилин, 1969) как проявление реакции следования за гибнущим по той или иной причине членом своего стада. В отношении причин самовыбрасывания китообразных высказываются различные предположения: ошибки в работе эхолокационного аппарата, плохо срабатывающего в условиях мелководья и мутной воды (Dudok van Heel, 1962), неизвестные массовые смертельные заболевания, психические расстройства (Geraci, 1978; и др.). Однако ни одна из этих гипотез не получила окончательного подтверждения. Подробное описание наблюдений за выбрасыванием стада гринд у берегов Флориды приводится в работе В. Феринга, Р. Велса (Fehring, Welb, 1976). Группа дельфинов, состоящая из 49 животных, на протяжении четырех дней несколько раз оказывалась на отмели. Операция по спасению китов крайне затруднялась тем, что отбуксированные в море животные вновь выбрасывались на берег. Все стадо удалось вернуть только после того, как далеко в море были отбуксированы два крупных животных, видимо, лидеров стада. После этого остальные дельфины прекратили попытки, выбрасываться на берег и стадо объединилось. Однако когда одного из молодых дельфинов спасенного стада решили взять для океанариума и за веревку, привязанную к его хвосту, начали подтаскивать к берегу, вновь все стадо последовало за ним. Отдельные животные стали выбрасываться на берег, разбиваясь о каменные гряды. Даже после того как молодую гринду оставили, некоторое время дельфины продолжали выбрасываться на берег. Но и в этот раз удалось спасти оставшихся в живых гринд, отбуксировав их вместе с лидером далеко в море. Спустя 4 дня часть этого стада (13 животных) были найдены обсохшими в 275 км от последнего места их обсыхания. Как сообщают авторы, море в эти дни было спокойным и вода прозрачной, что, видимо, исключает ошибки в работе эхолокационного аппарата. Среди выбросившихся животных не наблюдалось паники — они довольно спокойно двигались к берегу и не производили впечатления больных. Таким образом, во время спасательных операций реакция следования проявилась у гринд дважды, когда все стадо ушло в море за лидирующими животными и когда оно последовало за буксируемым к берегу молодым животным. Более сложный вид подражания — обучение путем наблюдений, или истинную имитацию, исследовали у многих видов зубатых китообразных в неволе: у атлантических афалин (McBride, Kritzler, 1951; Caldwell et al., 1965), у индийских афалин (Тауler, Saayman, 1973), у гринды (Kritzler, 1952), у малой косатки (Brown et al., 1966), у амазонского дельфина (Pilleri et al., 1980). Наиболее обстоятельное изучение различных форм моторного имитационного поведения у афалин проведено С. Тайлером, Г. Сайманом (Tayler, Saayman, 1973). Две взрослые самки афалины содержались в одном бассейне со взрослым самцом морского котика (Arctocephalus pusillus); в этом же бассейне находилось несколько пингвинов, черепахи и скаты. В первые месяцы их совместной жизни обе самки избегали котика, но постепенно степень их контактов стала возрастать, в результате чего в поведении афалин появились многие элементы поведения, присущие котику во время сна, груминга и активного плаванья. Так, дельфины стали плавать, подгребая плавниками как ластами, а хвостовой плавник в это время был совершенно неподвижен. Как известно, дельфины те занимаются грумингом. Однако, наблюдая эту форму поведения у котика, они переняли многие позы и действия, связанные с грумингом. Котики с помощью передних и задних ласт разглаживают и очищают свой мех, при этом они принимают разные позы: вращаются вокруг собственной оси у поверхности воды или принимают вертикальную позу выставив из воды только задние ласты. В это время к поверхности поднимаются пузырьки воздуха, выдыхаемого котиком. Дельфины, подражая котику, с помощью грудных плавников научились потирать живот, лежа на спине у поверхности воды, принимать вертикальную позу, выставив хвостовой плавник и выпуская при этом пузырьки воздуха из дыхала. Выныривая на поверхность, они широко раскрывали рот, подражая зевоте котиков, хотя известно, что зевота не свойственна дельфинам. В первое время своего пребывания в бассейне самки афалин плавали даже во время ночного отдыха. Котики же спали неподвижно у поверхности воды, лежа на одном боку, при этом задние ласты выставлены из воды; они имели дугообразную форму. Иногда они лежали на поверхности на спине, скрестив ласты на брюхе. Спустя некоторое время дельфины переняли позы котика, которые он принимал во время сна. Стали замечать, что время от времени афалины неподвижно лежали у поверхности на боку или на спине, прижав плавники к брюху и выставив хвостовой плавник из воды. Хотя эти позы были неудобны афалинам, они часто их принимали, находясь рядом со спящим котиком. Следует отметить, что котик не перенимал поз и движений дельфинов. Дельфины разных видов, содержащихся в одном бассейне, часто перенимают друг у друга некоторые формы поведения. Так, Д. Браун и др. (Brown et al., 1966) описывают имитацию прыжков вертящегося дельфина (Stenella lingirostris) афалинами, которые содержались в том же бассейне. Гринда пыталась балансировать на носу надувную игрушку, подобно тому как это делали афалины. Но поскольку у гринд отсутствует вытянутый рострум, она манипулировала игрушкой с помощью головы. Некоторые виды деятельности дельфины, главным образом афалины, могут перенимать у человека. Так, С. Тайлер, Г. Сайман (Tayler, Saayman, 1973) описывают некоторые виды орудийной деятельности афалин, которую они наблюдали у людей. Во время уборки бассейна афалины проявляли большой интерес к этой процедуре. Прибор для уборки бассейна состоял из полого скребка со шлангом, куда втягивался мусор, который собирали со дна бассейна с помощью скребка. Однажды аппарат был оставлен на ночь в бассейне. Им завладела одна самка и долго с ним манипулировала, таская во рту по бассейну. Затем, подражая действиям человека, она, придерживая шланг грудными плавниками, взяла в рот совок и стала водить им по дну бассейна. Когда аппарат убрали, обе самки стали использовать для «уборки» бассейна куски кафельной плитки (6x8 см), зажимая их зубами. Действуя сообща, они научились с помощью кафельных плиток собирать в кучу куски морских водорослей (Enteromorpha), которые они употребляли в пищу. Эти примеры использования орудий как в форме игры (чистка бассейна), так и для сбора пищевых объектов свидетельствуют о том, что они могут не только копировать определенные виды деятельности человека, но и модифицировать их сообразно своим потребностям. Имитация человеческой деятельности свойственна не только афалинам, но и речным дельфинам. В гл. 2 мы приводили примеры того, как амазонские дельфины, подражая человеку, «чистили» стенки бассейна и окна с помощью небольших щеточек, которые они держали во рту (Pilleri et al., 1980). Было проведено (Adler, Adler, 1978) экспериментальное исследование способности афалин к обучению путем наблюдений на основе использования инструментальной условнорефлекторной методики. Опыты были проведены на трех афалинах, которые находились в круглом бассейне, разделенном на четыре отсека крупноячеистой сеткой. Дельфинам предложили четыре задачи: втягивание свисающего конца веревки в бассейн; дифференцирование двух квадратов с горизонтальной и вертикальной исчерченностью; дифференцировку двух квадратов красного и зеленого цвета; различение цели по ее местоположению (на левой или правой стороне). Все цели предъявлялись животным над водой. С одним из дельфинов — «демонстратором» проводили эксперимент, а два других находились в это время в соседних отсеках и могли наблюдать все, что происходит в экспериментальном отсеке. После закрепления навыка у «демонстратора» одного из «наблюдателей» переводили в экспериментальный отсек и проводили с ним процедуру обучения. В опытах те же дельфины были и в роли «демонстраторов» и в роли «наблюдателей». В последней задаче «наблюдателя» помещали в экспериментальную камеру спустя четыре дня после проведения опытов с «демонстратором». Оценка результатов опытов показывает, что дельфины могут обучаться сложным задачам дифференцировать цели путем наблюдений даже с отсрочкой на несколько дней. У дельфинов-наблюдателей успешное решение задачи происходило уже с первых предъявлений условных раздражителей. В то же время для обучения дельфина-демонстратора требовалось большее число тренировок. У «наблюдателей» по сравнению с «демонстраторами» заметно сокращался латентный период ответов (от 64 до 25 с). Авторы полагают, что наблюдения за действиями «демонстратора» уже сами по себе являются для дельфинов положительным подкреплением всей ситуации и снижают у них пассивно-оборонительную реакцию перед незнакомыми предметами, которая в сильной степени развита у этих животных. Л. Г. Воронин (1977) также считает, что подражательный рефлекс способствует приучению обезьян к экспериментальной обстановке, сокращая начальный период формирования условнорефлекторной деятельности. Приведенные примеры демонстрируют высокоразвитую способность дельфинов к воспроизведению сложных моторных действий, но вместе с тем у них в равной степени развита и «вокальная» имитация. Исследования показали, что дельфины способны имитировать широкий диапазон самых разнообразных звуков — от булькающих звуков поднимающихся пузырьков воздуха из аквалангов до отдельных слов и простых фраз, произносимых человеком (Lilly, Miller, 1962; Caldwell, Caldwell, 1972a; Beach, Papper, 1973; Tayler, Saayman, 1973; Тарчевская и др., 1978; Richard et al., 1981). Изучение имитации тональных сигналов с частотной модуляцией у афалин было проведено (Тарчевская и др., 1978) на двух взрослых афалинах. Эталонные сигналы с частотной модуляцией от 5 до 10 кГц, близкие к естественным свистам дельфинов, транслировались под водой. Оценка сходства эталонных сигналов с сигналами дельфинов осуществлялась на экране осциллографа. Было выяснено, что дельфины имитировали эталонные сигналы свистами, превышающими последние по частоте в 1,2 — 1,8 раз. Иногда дельфины могли повторять эталонный сигнал два-три раза в пределах одного имитационного. Наблюдалось как непосредственное повторение только что проигранного сигнала, так и постепенное преобразование собственных свистов в имитационные. Недавно проведенные исследования способности афалин к имитации искусственных свистовых сигналов (Richard et al., 1981) показали, что они легко справляются с этой задачей. Самец афалина, имевший до опытов определенный набор естественных свистовых сигналов, которые часто и стереотипно повторялись, был обучен имитировать различные вариации звуковых «моделей», излучаемых генератором. Все предложенные искусственные звуки дельфин научился точно воспроизводить, используя свои «вокальные» возможности. Эти звуковые сигналы отличались от его естественных свистов. При проведении опытов авторы отметили большие возможности дельфина к обобщению, что позволило ему воспроизводить новые сигналы после 1 — 3 предъявлений. Скорость обучения воспроизведению искусственных звуковых сигналов свидетельствует, по мнению авторов, о высокоразвитой способности дельфинов оперировать большим объемом слуховой информации, а способность к «вокальной» имитации играет существенную роль при установлении коммуникативных связей в их природных сообществах. Широко известные эксперименты Дж. Лилли (Lilly, 1962), целью которых было изучение способов установления контактов между человеком и дельфинами, показали, что дельфины обладают чрезвычайно развитыми «вокальными» подражательными способностями. Опыты Дж. Лилли подтвердили, что афалина может в какой-то степени подражать человеческому голосу и после обучения способна воспроизводить слоги, отдельные слова, простые фразы, восклицания, смех. В своих экспериментах в качестве подкрепления он применял электрическое раздражение подкорковых структур мозга, так называемых «центров удовольствия». Сравнение виброграмм с записью голосов дельфинов и человека подтвердило звукоподражание афалины. Человек произносил слова в пределах колебаний от 200 до 3 000 Гц, а дельфин воспроизводил их в частотах от 1 000 до 8 000 Гц. Если магнитофонную ленту с голосом дельфина, имитировавшим слова человека, пропускать с замедленной в 2 — 4 раза скоростью, то воспроизводится известное сходство с голосом человека. Дальнейшие эксперименты Дж. Лилли по обучению дельфинов подражанию человеческой речи, как известно, не подтвердили ожиданий автора о возможности установления сознательных контактов между дельфином и человеком. Тем не менее эти опыты продемонстрировали большие возможности дельфинов в сфере имитации «гомоноидных» звуков, что само по себе можно рассматривать как большую удачу исследователя. Интересные исследования взаимоотношений дельфинов в группе при выработке у них пищедобывательных условных рефлексов и некоторых форм имитационного поведения проведены Л. Г. Ворониным и др. (1971, 1974). Группа состояла из трех черноморских афалин: двух самцов и одной самки. Один самец занимал доминирующее положение в группе. Было проведено несколько серий экспериментов, где выяснилось распределение между дельфинами манипуляторов, при воздействии на которые они сразу получали рыбу. В первой серии опытов дельфины могли нажимать на три рычага, во второй серии им оставляли только один рычаг, а в третьей — получение рыбы и манипуляторы были пространственно разделены, т. е. нажав на рычаг, дельфин должен был плыть за рыбой в другой конец бассейна. В результате проведенных экспериментов было выяснено, что при наличии трех рычагов внимание дельфинов привлекал в основном тот, который выбирал доминантный самец. Второй самец предпочитал подбирать рыбу, которая оставалась от работы доминанта. Насытившись, доминант охотно работал для второго самца, усердно нажимая на рычаг, и отдавал ему всю рыбу. Самка сначала без всякого предпочтения нажимала на разные рычаги, но затем чаще стала пристраиваться к самцам и работала обычно в паре с доминантой. При ситуации, когда оставляли только один рычаг, все животные поочередно на него нажимали, но чаще других доминантный самец. Несколько раз доминант отгонял второго самца от манипулятора и преследовал его с агрессивными намерениями. В том случае, когда работа с манипулятором вознаграждалась в другом месте, чаще проявлялось деспотическое поведение доминирующего самца. Он отгонял не только второго самца, но и самку; установилась очередь в работе: сначала к манипулятору подходил только доминант, затем к добыванию пищи присоединялся второй самец, потом к работе приступала самка. Из приведенной работы следует, что конкурентные отношения между доминантой и подчиненным дельфином возникали не по поводу получения рыбы, а из-за доступа к рычагу, т. е. способу добывания пищи. Таким образом, активный вид деятельности, приводящий к добыванию пищи, является, по-видимому, прерогативой доминирующего животного в сообществе дельфинов. Рефлекс подражания у дельфинов данной группы проявился в том, что наибольшее внимание животных привлекает тот манипулятор, который выбирал доминирующий самец. Два «свободных» рычага мало интересовали дельфинов, хотя с их помощью они также могли добывать пищу. Обсуждая результаты проведенных экспериментов, авторы отмечают миролюбие и отсутствие жестких конкурентных отношений дельфинов в условиях группового эксперимента. Редко возникающие агрессивные стычки очень быстро сменялись миролюбивыми взаимоотношениями. Способность доминирующего самца нажимать на рычаг после собственного насыщения и таким образом «кормить» другого дельфина рассматривается авторами как проявление сотрудничества в сообществе дельфинов. На основании приведенных в этом разделе данных можно заключить, что у дельфинов в сильной степени развиты различные формы подражательной деятельности, которые проявляются в виде реакции следования за лидерами, синхронного плавания, а также в более сложных формах научения путем подражания. Дельфины имитируют движения, позы и звуки не только у особей своего вида, но и у животных, далеко отстоящих от них в таксономическом отношении. Некоторые формы орудийной деятельности, которые они могут перенимать у человека, можно рассматривать одним из показателей их развитого интеллекта. Среди млекопитающих по развитию имитационных способностей дельфины находятся на том же уровне, что и человекообразные обезьяны. Несомненно, развитая способность к сложным формам подражания играет весьма существенную роль в социальной жизни дельфинов. Этот вопрос вызывает большой интерес у исследователей. Л. Герман (Herman, 1980) полагает, что способность дельфинов к «вокальной» и моторной имитации является той основой, на которой развиваются различные формы координированной групповой активности: синхронное плавание, коллективная охота, защитное поведение. Интересную мысль по поводу функциональной роли имитационного поведения дельфинов в их общественной жизни высказывают С. Тайлер, Г. Сайман (Tayler, Saayman, 1973). Авторы предполагают, что имитация половых форм поведения в период ухаживания может играть существенную роль при выборе полового партнера. Синхронность выполнения ритуальных поз ухаживания и координация движений во время совместного плавания, по мнению авторов, может служить стимулом для селективного спаривания и для упрочения социальных связей в сообществе. Другими словами, в каждом сообществе могут существовать собственные «традиции» половых взаимоотношений, которые сохраняются и усиливаются в результате подражания друг другу. Развитая способность к «вокальной» имитации у дельфинов, по мнению Е. Вильсона (Wilson, 1975), может быть причиной большого сходства опознавательных сигналов у животных, принадлежащих к одному сообществу, что дает им возможность быстро узнавать своих сородичей. Известно, что у афалин обнаружены (Caldwell, Caldwell, 1972а) устойчивые индивидуальные «свисты-автографы», представляющие собой определенные контуры частотных сигналов. Они служат опознавательными сигналами для каждого члена группы. Вместе с тем можно предположить, что физические характеристики этих «свистов-автографов» у членов одного сообщества в результате подражания могут приобрести общие черты, или свойства, отличающие их от подобных индивидуальных сигналов другого сообщества. По этим сигналам дельфины могут узнавать своих соплеменников и отличать особей, принадлежащих к другим сообществам. Приведенные выше предположения исследователей о роли имитационных рефлексов в социальной жизни дельфинов весьма интересны, но требуют экспериментальных подтверждений.
Память
Изучение памяти является одной из актуальных проблем поведения животных, поскольку процессы записи, хранения и воспроизведения информации входят непременным компонентом в любую форму их деятельности — от элементарного условного рефлекса до образования абстрактных понятий. Исследование особенности процессов запоминания у животных с высокоорганизованным мозгом представляет наибольший интерес, поскольку их мозг может хранить и перерабатывать большой объем информации (Lilly, 1978). Экспериментальное изучение памяти у дельфинов стало интенсивно развиваться только в последние годы (Herman, Bailey, 1970; Herman, Gordon, 1974; Протасов, Сергеев, 1974; Шурепова и др., 1975; Thompson, 1976; Herman, 1980). Известны две фазы памяти: лабильная, или оперативная, которая соответствует удержанию следа в форме реверберации нервных импульсов (по последним данным, при этом происходят ультраструктурные изменения нейронов), и стабильная (долговременная), которой соответствует сохранение следа за счет биохимических изменений в синапсах (Соколов, 1969, 1981). Функция кратковременной памяти состоит в отборе и фиксации новой нужной информации и оперирование ею в соответствии с требованиями данной ситуации (Baddeley, Hitch, 1974), Оперативная, кратковременная, память играет существенную роль в формировании адекватных, пластичных и быстрых реакций животных на изменение окружающей обстановки. Основное внимание исследователей в экспериментах на дельфинах было сосредоточено на изучении их кратковременной, или оперативной, памяти — той основе, на которой может развиваться способность к «отвлеченной символизации» предметов и явлений окружающей их среды (Herman, 1980). Изучение кратковременной, оперативной, памяти у афалин проводилось с помощью метода отсроченной идентификации стандартного стимула — образца при восприятии ими слуховых и зрительных сигналов (Herman, 1980). Л. Герман, Дж. Гордон (Herman, Gordon, 1974) исследовали способность афалин помнить в течение определенного отрезка времени, какой из двух или нескольких звуковых сигналов предъявлялся в начале опыта. На один из двух звуковых сигналов у дельфинов был выработан рефлекс — нажимать на педаль, расположенную у соответствующего излучателя звука (импульсного или непрерывного). Во время опыта подавался один из двух сигналов, продолжавшийся 4 с (образец), с некоторой временной задержкой (от 15 до 120 с) транслировались оба сигнала подряд с интервалом 0,5 с в случайной последовательности (тестовый сигнал). Нажатие дельфином педали около излучателя, который передал звук, соответствующий образцу, считался правильным ответом и поощрялся рыбой (рис. 2). Дельфину предлагали разные варианты этой задачи. Так, после предъявления сигнала-образца через определенный интервал передавали еще один сигнал, который либо соответствовал «образцу», либо не соответствовал. По тому, какую из педалей нажимал дельфин, можно было судить, правильно или неправильно он сопоставил разделенные во времени звуковые сигналы. В наиболее сложном варианте задачи дельфину предлагалось выбрать стимул-образец из набора разных звуков (Thompson, Herman, 1977). При проведении этого эксперимента дельфину вначале давали прослушать серию из восьми новых для него случайно выбранных звуков. Спустя 4 с транслировали через один из двух боковых излучателей звуковой сигнал. Дельфину предлагалось решить, совпадает ли этот звук с одним из ранее передаваемой серии сигналов. Реакция дельфина на педаль, расположенную у излучателя звука, являлась положительным ответом. Это означало, что «образец» совпадал с одним из сигналов серии. В том случае, если «образец» не совпадал ни с одним из восьми ранее передаваемых звуков, дельфин должен был нажать педаль около «молчащего» излучателя. Дельфин справился с этим сложным экспериментом с высокой степенью достоверности правильных ответов. Зрительная память у афалин исследовалась (Thompson, 1976) методом сравнения зрительного образа со стимулами, которые демонстрировались ему спустя определенный промежуток времени. Все визуальные стимулы предъявлялись дельфину под водой через застекленное окно размером 46x92 см. Один стимул представлял собой белый круг на черном фоне (8,3 см в диаметре), второй — черный круг на белом фоне того же размера. До начала основного эксперимента дельфин был обучен дифференцировать эти фигуры. В начале опыта ему показывали одну из фигур, а затем сразу или через определенный интервал времени демонстрировали в окне два стимула. Дельфину следовало нажать педаль, расположенную у той фигуры, которая демонстрировалась вначале. Дельфин не мог обучиться сравнивать зрительные стимулы, даже если «образец» демонстрировался одновременно с тестовыми фигурами, после 100-го предъявления этой задачи. Процедура обучения выбора по образцу из двух зрительных стимулов значительно облегчилась, после того как предъявление каждого из зрительных стимулов стали сопровождать звуками. На первом этапе обучения эти звуки были пространственно разнесены и проигрывались рядом с изображениями предметов, затем, постепенно сближая источники звуков, их стали подавать из одного центрального излучателя. После серии успешных опытов демонстрацию зрительных изображений перестали сопровождать звуками. Тем не менее дельфин продолжал правильно решать предъявленную задачу отсроченного выбора зрительных стимулов по образцу. Максимальный интервал во времени между «образцом» и демонстрацией тестовых изображений не превышал 30 — 35 с. Л. Герман (Herman, 1980) объясняет облегчение зрительного распознавания дельфином стимулов после временного присоединения к ним звуковых сигналов тем, что ассоциативное обучение осуществлялось в центральном представительстве более высокоразвитой у них слуховой системы, где зрительные образцы были представлены слуховыми кодами.
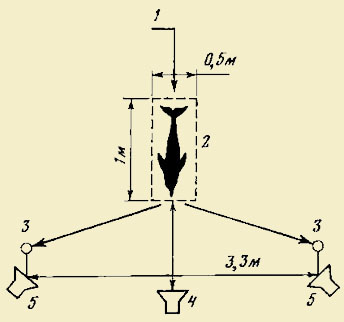 Рис.
2. Схема эксперимента с афалинами
«отставленного выбора по образцу
акустических сигналов» 1 — вход в
экспериментальный бассейн; 2 — площадка
прослушивания; 3 — боковые педали; 4 —
центральный излучатель звука; 5 — боковые
излучатели звуков (по: Herman, 1980)
Рис.
2. Схема эксперимента с афалинами
«отставленного выбора по образцу
акустических сигналов» 1 — вход в
экспериментальный бассейн; 2 — площадка
прослушивания; 3 — боковые педали; 4 —
центральный излучатель звука; 5 — боковые
излучатели звуков (по: Herman, 1980)
Результаты приведенных выше исследований свидетельствуют о том, что у дельфинов наиболее развитой оказалась слуховая оперативная память. Это согласуется с морфологическими данными о более сложном строении слуховых отделов центральной нервной системы китообразных (Богословская, Солнцева, 1979). У дельфинов оперативная слуховая память по сравнению со зрительной обладает большей емкостью, дольше удерживает новую информацию, отличается пластичностью при кодировании событий, «считывании» и реализации следовых процессов. При сравнении уровня развития зрительной и слуховой памяти у дельфинов и обезьян было выяснено (Herman, 1980), что приматы в отличие от дельфинов гораздо лучше справляются со сложными задачами, где им приходится идентифицировать зрительные стимулы и плохо обучаются сопоставлять разделенные во времени слуховые сигналы (Worsham, D'Amato, 1973). Так, опыты, проведенные на капуцинах, в которых использовался незнакомый для них набор новых предметов, доказали, что обезьяны с задержкой до 8 мин могли выбирать стимул — образец из набора от 2 до 8 объектов (D'Amato, 1973). Сопоставляя данные обучения дельфинов и обезьян по методу отставленного выбора по образцу, Л. Герман (1980) приходит к выводу, что зрительная оперативная память у обезьян столь же хорошо развита, как слуховая у дельфинов. Изучение у собак роли различных рецепторов в продолжительности запоминания комплексного восприятия местонахождения пищи (Беритов, Айвазашвили, 1967) показали, что наибольшее значение для сохранения в памяти местонахождения пищи имеет вкусовое и обонятельное восприятия, меньшее значение имеют зрительное и вестибулярное восприятия. Таким образом, у разных видов животных возможности памяти (восприятие, фиксация и воспроизведение) определяются уровнем развития их сенсорных систем, избирательностью выбора и считывания информации со следами предыдущих событий, а также особенностью эффекторных систем в воспроизведении сигнала (след памяти может быть воспроизведен лишь в пределах, ограниченных возможностями исполнительных органов). На основании приведенных выше данных можно заключить, что дельфины успешно хранят и перерабатывают большой объем, информации, что дает возможность сравнивать их с приматами. При этом наиболее быстрое, лабильное воспроизведение следовых процессов и сопоставление их с текущими событиями происходит в пределах слуховой системы. Лабораторные методы исследования, к сожалению, пока не позволяют нам составить более полного представления о возможностях использования дельфинами ресурсов своей памяти в естественной жизни. Можно согласиться с мнением Дж. Лилли (Lilly, 1978), что развитие мозга дельфина обусловлено большим объемом информации, которую он воспринимает и обрабатывает. В настоящее время можно говорить о трех возможных источниках необычайно большой по объему информации, которыми могут располагать китообразные. Одним из возможных источников большого объема информации может явиться восприятие акустических образов окружающих предметов. Это предположение было впервые высказано Дж. Лилли еще в 1963 г. Предполагается, что мозг дельфинов также оперирует акустическими аналогами, как мозг человека — зрительными. Сложнейшие звуковые сигналы дельфинов и других китообразных, которые никак не поддаются расшифровке, могут содержать не слова и фразы, как у людей, а некое подобие голограмм. Это также могло бы привести к многократному увеличению передаваемой и обрабатываемой информации. Активная эхолокация, как полагают В. А. Протасов, Б. Ф. Сергеев (1974), требует развитой оперативной памяти, так как для получения исчерпывающей информации о подводных объектах необходимо иметь возможность сравнивать зондирующую посылку с эхосигналом, возникшим при отражении сигнала от лоцируемых тел. Сложная социальная организация сообществ у китообразных, поддерживающаяся в первую очередь с помощью звуковых сигналов при идентификации членов сообщества, установления контактов внутри группы, поиска полового партнера и т. д., также связана с восприятием и долговременным хранением большого объема слуховой информации. Другим источником информации можно считать хеморецепцию. С того времени, как Л. И. Суховской (Suchovskaya, 1972) было доказано наличие вкусовых луковиц на языке дельфинов, а работами В. Е. Соколова, В. Д. Кузнецова (1971), В. Д. Кузнецова (1978) была экспериментально подтверждена возможность восприятия малых концентраций некоторых веществ в воде, предположение о существовании у зубатых китообразных хеморецепции (Яблоков, 1957, 1961) превратилось в установленный факт. Учитывая, что запахи в водной среде должны сохраняться много дольше, чем в воздухе (Яблоков и др., 1972), роль хеморецепции у китообразных по объему улавливаемой информации может быть сравнима, по мнению автора, с ролью зрительного анализатора у животных, обитающих на суше. Третий источник информации следует из особенностей социальной организации их группировок. В каждом стаде дельфинов, видимо, могут находиться одновременно особи пяти-шести поколений. Этот факт становится очевидным, если учесть, что продолжительность жизни у большинства видов дельфинов составляет около 25 лет, а половая зрелость наступает в 2 — 5 лет. Подобное соотношение одновременно живущих поколений можно встретить только у высших приматов (продолжительность жизни шимпанзе около 60 лет, а половозрелость наступает к 6 — 10 годам) и слонов. Указанная особенность состава социальных группировок у китообразных и высших приматов может в немалой степени способствовать передаче «традиций» и накоплению большого объема информации от поколения к поколению.
Некоторые подходы к изучению способов общения у дельфинов
Энтузиазм Дж. Лилли, направленный на изучение дельфинов, и его попытки найти способы общения человека и дельфина (Lilly, 1961 — 1978) побудили многих ученых заняться этой проблемой. Появились серии работ, в которых исследовались механизмы звукового общения дельфинов между собой, выяснялось, могут ли дельфины обмениваться осмысленной информацией, существует ли у них язык символов (Dreher, Evans, 1964; Lang, Smith, 1965; Busnel, 1966; Bastian, 1967; Batteau, Markey, 1968; Caldwell, Caldwell, 1972 c; Herman, Arbeit, 1973; Белькович, Казнадзей и др. 1978; Марков, 1978; Herman, 1980; Reiss et al., 1981). Одну из самых интересных работ в этой области выполнил Дж. Бастиан (Bastian, 1967). Применив методику оперантного обучения, Дж. Бастиан хотел выяснить, может ли один дельфин, наблюдая за условными оптическими сигналами, сообщить с помощью акустических сигналов другому дельфину, визуально от него изолированному, какой из двух манипуляторов следует нажать, чтобы оба получили пищевое поощрение. В опыте участвовали две афалины — самка и самец, которые были обучены нажимать на левый или правый рычаг в зависимости от того, какой условный сигнал включался (мигающий свет или непрерывный). В первой серии опытов дельфинов обучали в разных отсеках бассейна, разделенных сетью, сквозь которую дельфины могли видеть и друг друга, и сигнальные лампы, находящиеся в отсеке самки. В отсеке самца над рычагами помещалась лишь одна лампа, включение которой служило сигналом начала эксперимента. Когда у обоих дельфинов был выработан условный рефлекс, их разделили перегородкой, через которую они не видели друг друга, но акустическая связь между ними сохранялась. Самец мог знать о начале эксперимента по включению лампы в его отсеке, а условные сигналы (указывающие левый или правый рычаг следует нажимать) включались только в отсеке самки. Несмотря на то что самец не видел сигнальных ламп, он вскоре стал правильно (в 90% случаев) нажимать рычаги в соответствии с тем сигналом, который подавался самке. Изменение сигнального значения условных раздражителей, т. е. результат «обратного обучения» показал, что и в этом случае самец стал правильно нажимать на педаль. Только после введения звуко- и светонепроницаемой перегородки самец стал случайно нажимать на педали. Результаты столь удачно начатого эксперимента, естественно, привели исследователей к мысли, что самка с помощью акустических сигналов сообщала самцу, какой из рычагов, левый или правый, следует нажать, чтобы получить вознаграждение. Тщательный анализ магнитофонной записи звуков самки показал, что самец действительно ориентировался на звуки, издаваемые самкой при выборе рычагов. Однако доказательств тому, что самка понимает связь между своими сигналами и поведением самца, не было получено. После того как самца перевели из отсека в бассейн и убрали ширму, самка продолжала издавать те же эхолокационные щелчки, на которые ориентировался самец при выборе манипулятора. Дж. Бастиан полагает, что у самки в начале опытов случайно или по какой-то неясной причине выработался рефлекс на включение непрерывного света издавать серию щелчков. У самца также выработался условный рефлекс на серию щелчков, издаваемых самкой, нажимать на правый рычаг в то время, когда он мог видеть самку и сигналы лампы. Автор полагает, что в основе этого сложного согласованного действия лежит самодрессировка самки издавать разные звуковые сигналы в ответ на включение различных световых сигналов, а также самодрессировка самца выбирать рычаг в соответствии с услышанным звуковым сигналом самки. Опыты Дж. Бастиана не дали ответа на вопрос о существовании у дельфинов языкового смыслового общения, но выясненное в эксперименте умение дельфинов справиться со сложной задачей выработки тонкой диффереицировки условных сигналов при совместной работе может свидетельствовать о высоком уровне развития их высшей нервной деятельности. Данные этих опытов показывают, насколько сильно развит у дельфинов общественный инстинкт. Звуки, производимые партнером, видимо даже не относящиеся к коммуникационным, являются достаточно сильными раздражителями, на основании которых могут формироваться рефлексы второго порядка у членов сообщества. Непосредственное изучение акустических сигналов, которыми обменивались два дельфина вне визуального контакта, было проведено Т. Лангом, Дж. Смитом (Lang, Smith, 1965). Они поместили двух дельфинов афалин в разные бассейны и установили между животными двустороннюю акустическую связь с помощью гидрофонов и подводных динамиков. Время опыта (30 с) было разбито на 16 интервалов длительностью от 80 до 150 с каждый. В период интервалов с четными номерами между бассейнами устанавливалась акустическая связь, во время интервалов с нечетными номерами такая связь отсутствовала. Типы издаваемых свистов были разделены на 6 групп (А — Е). Записи показали, что в периоды отсутствия акустической связи голосовая активность дельфинов резко падала по сравнению с периодами включения связи. В начале опытов при отсутствии связи регистрировали свисты трех типов — А, Б, Г. В период включения связи также присутствовали свисты А, Б, Г и добавлялся еще один тип — Е. По мере увеличения числа опытов менялся характер звуков в периоды включения связи. Присутствовали все 6 типов свистов и наблюдалась определенная последовательность типов свистов: Б — Г — Б — Г. При этом все свисты Б издавала самка, а свисты Г — самец. Спустя 4 месяца запись звуков, издаваемых самкой, была несколько раз проиграна самцу. На свист самки Б самец отвечал свистом типа Г и замолкал каждый раз на том месте, где воспроизводился свист самки Е. На этом основании авторы сделали вывод, что свисты типа Б и Г — это зов другого дельфина; свист А — приглашение вступить в голосовое общение. Более сложный свист Е, по мнению авторов, может нести определенную смысловую нагрузку, так как этот звук присутствовал только во время двусторонней связи и не воспроизводился дельфинами при механическом одностороннем воспроизведении. Основной вывод, который можно сделать из проведенных Т. Лангом и Дж. Смитом экспериментов с двусторонней «телефонной» связью двух дельфинов: они активно обмениваются звуковыми сигналами, которые, видимо, несут разные функциональные нагрузки. Но могут ли дельфины передавать с их помощью какую-либо смысловую информацию на основании этих опытов, сказать нельзя. Итак, ни эксперименты Дж. Бастиана, ни опыты Т. Ланга и Дж. Смита не выяснили возможности языкового общения у дельфинов. Главной неразрешенной проблемой в экспериментах Дж. Бастиана остается возможность влияния дельфинов друг на друга с помощью эхолокационных звуков. До тех пор, пока не будет устранен этот фактор, трактовка опытов не может быть однозначной. Е. Н. Панов (1980) высказывает интересную мысль о том, что объединение обеих методик могло бы внести некоторую ясность в понимание результатов опытов Дж. Бастиана. Было проведено немало специальных акустических исследований, связанных с попытками расшифровок языка дельфинов (Dreher, Evans, 1964; Busnel, 1966; Caldwell, Caldwell, 1968a, 1972c; Белькович, Казнадзей, и др., 1978). Одним из интересных исследований явились работы В. И. Маркова (Марков, 1978; Марков, Островская, 1978). Авторы выделили в «словаре» афалины 7 исходных сигналов, из которых формируется 31 элемент более сложного состава. Последние комбинируются друг с другом, образуя большое количество различных сигналов. В работах В. И. Маркова продемонстрировано наличие (высокого уровня синтаксической организованности звуковых сигналов черноморской афалины. У дельфинов было обнаружено до 6 уровней комбинирования первичных сигналов. Обнаруженная организация звуковых сигналов подтверждает, что звуковое общение между дельфинами может нести сложную информацию. Возможности установления контактов с дельфинами с помощью свистовых сигналов, передаваемых человеком, впервые были осуществлены в опытах Д. Батто. Эксперименты Д. Батто, П. Маркей (Batteau, Markey, 1968), предпринятые с целью выяснения «языковых» способностей дельфинов, выявили у них удивительные возможности выполнять точные команды на различные свистовые сигналы. Д. Батто сконструировал электронный прибор, преобразующий звуки человеческой речи в свистовые, подобные тем, что издают дельфины. Запись велась постоянно, и по графикам можно было визуально сравнивать между собой свисты дельфинов и копирующего устройства. Опыты проводились на двух афалинах. Подавались определенные команды типа: «вперед», «направо», «налево», «выйди из обруча», «ударь ластом по мячу» и т. д. За время работы оба животных научились правильно исполнять 15 таких команд: толкать мяч, проплывать сквозь обруч, поднимать хвост, издавать эхолокационный щелчок и т. д. Эти эксперименты показывают, что поведением дельфинов можно управлять с помощью свистов. Хотя животные воспринимают услышанное не как речь, а просто как условный звуковой сигнал, они точно выполняли требуемые действия. Второй эксперимент, проведенный с этими же дельфинами, ставил цель выяснить, может ли дельфин правильно выполнять команду, которой его специально не учили, но он наблюдал процедуру тренировки со вторым дельфином. На команду «издай эхолокационный щелчок» не обученный этому ранее дельфин сразу же правильно отреагировал, издав щелчок. Когда этой же самке подали команду «подними хвост», она им шлепнула по воде. Этот опыт еще раз подтверждает удивительно развитые имитационные способности у дельфинов, сформировавшиеся в результате их стадного образа жизни. Группой сотрудников Гавайского университета (Herman, Arbeit, 1973; Herman, Gordon, 1974; Herman, 1980) на трех афалинах были проведены исследования, в которых выясняли уровень усваивания животными простых языковых звуковых стимулов. Эксперименты были разработаны только для выяснения возможности восприятия различных комбинаций звуковых символов, а не для их воспроизведения. Отправной точкой постановки данных экспериментов было допущено, что у человека в онтогенезе раньше формируются связи, обеспечивающие понимание языка, чем его воспроизведение. Для дельфинов была разработана кодовая слуховая система, в которой окружающие дельфинов в бассейне предметы и действия, имеющие к ним отношение, были обозначены определенными, произвольно выбранными звуковыми сигналами. Эти сигналы производились с помощью электронного генератора и транслировались в воду. В первой серии опытов дельфина по кличке Ки обучали при демонстрации разных объектов в бассейне (мяча, спасательного круга и пластикового цилиндра) выбирать один из них при действии соответствующего этому предмету звукового сигнала. После закрепления навыка дельфина начали тренировать на выполнение парных звуковых команд из двух слов, обозначающих объект — действие, например: «коснись мяча», «возьми мяч», «принеси мяч». Дельфин в течение 4 месяцев обучился выполнять 9 комбинаций трех действий с тремя предметами. В конце опытов дельфина обучили по соответствующему звуковому сигналу приносить объекты только определенному экспериментатору. В следующей серии экспериментов участвовали два дельфина. Их тренировали в одном бассейне. Имя каждого дельфина обозначалось звуковым сигналом. Опыт проходил следующим образом. Одного из дельфинов по имени вызывали на стартовую площадку. Услышав «свой» сигнал, он нажимал на педаль, расположенную на его площадке. Далее следовала команда-инструкция, состоящая из двух слов: существительное — глагол, сформированная по той же схеме, что и у Ки. Если инструкция выполнялась дельфином правильно, подавался звук, обозначающий «да» (правильный ответ), когда дельфин ошибался, включали сигнал «нет». В этой серии экспериментов одного дельфина тренировали на выполнение звуковых команд, некоторые команды другому дельфину подавались с помощью жестов. Словарный запас дельфинов был доведен до 25 слов (существительных и глаголов). Оба дельфина оказались способны различать 44 парных комбинаций команд, составленных из этого словарного запаса. После усвоения парных команд дельфинам стали предъявлять команды, состоящие из трех слов, например «правый мяч выброси». Дельфин по этой команде выбирал мячик, расположенный от него справа, и выбрасывал из воды. Трехсложная команда типа «мяч принеси обруч» обозначала, что дельфин должен поместить мячик рядом с обручем. По команде «обруч принеси мячик» дельфин должен был отнести обруч в то место, где находился мяч. Такие сложные команды правильно выполнялись в среднем в 50% случаев. Это значительно превышает вероятность случайного выбора, так как число возможных ответов было очень велико. Правильные ответы на 600 — 700 команд из двух слов составили 80%. Число правильных ответов по одной команде падало с увеличением числа объектов (6 — 11), из которых производился выбор. Проведенные исследования показали, что дельфины способны в лабораторных условиях к случайно выбранной элементарной символизации окружающих их явлений и предметов. Однако, как полагает автор (Herman, 1980), эти эксперименты не означают, что в природе у дельфинов существует подобная символизация окружающей их среды. Недавно опубликовано сообщение (Reiss et al., 1981) о начале исследований возможности дельфинов с помощью сложных двигательных навыков оповещать экспериментатора о событиях окружающей их обстановки в океанариуме. Три афалины были обучены инициативно использовать кодовые сигналы для получения игрушек, добывания пищи, установления тактильных контактов с человеком. Во время обучения использовали не только пищевое поощрение, но и игровое, применяли также некоторые формы отрицательных подкреплений их нежелательных действий. Первые этапы обучения прошли успешно. Можно выделить основные направления исследований способов общения у дельфинов. Один путь — это изучение структуры самих звуковых сигналов, выделение элементарных исходных единиц — «фонем», анализ последовательности чередования разных типов сигналов и т. д. Другой подход направлен на выяснение вопроса, могут ли дельфины обмениваться содержательными сообщениями о характере и расположении окружающих их предметов, сигнализирующих о возможности получения пищи и др. И третий подход — это попытки установления контактов между человеком и дельфином с помощью сигналов-символов, обозначающих предметы и явления окружающей их среды; сюда же входит обучение дельфинов человеческой речи. Третий подход наиболее широко используется при изучении «языкового поведения» у обезьян и дельфинов. Попытки обучения шимпанзе членораздельной человеческой речи (Hayes, Hayes, 1952; Lieberman, 1968) не привели к желаемым успехам. Исследователи пришли к заключению, что, несмотря на удивительно развитую способность к подражанию, голосовой аппарат шимпанзе не приспособлен к воспроизведению речи человека. При попытках обучения дельфинов человеческой речи (Lilly, 1962) также не были достигнуты большие успехи, чем при обучении обезьян. Опыты Д. Батто, П. Маркей (Batteau, Markey, 1968), Л. Германа (Herman, 1980) являются одним из подходов к изучению у дельфинов способности к символизации внешнего мира. Этот путь исследований, как известно, привел к успешным результатам при обучении шимпанзе азбуке жестов, используемых глухонемыми в Америке (Gardner, Gardner, 1972). Исследования Б. и Р. Гарднеров показали, что шимпанзе, обученные азбуке жестов, могут использовать знаки-символы в качестве средства общения с экспериментаторами и другими шимпанзе. Шимпанзе по кличке Уошо, выучив несколько жестов в конкретной обстановке, могла вследствие генерализации употреблять эти жесты в другой ситуации. Так, жест «открыть», выученный по отношению к ящику с игрушками, шимпанзе самопроизвольно применяла по отношению к двери и водопроводному крану. Таким образом, использование шимпанзе их словарного запаса представляет творческий процесс и не является результатом простого заучивания слов по методу условного рефлекса. Замечательные исследования супругов Б. и Р. Гарднеров обнаружили у животных способность к символизации окружающего мира — тех качеств, которые лежат в основе языка человека. Однако это открытие еще не подтверждает представления о том, что у шимпанзе реально существует язык, подобный человеческому. Е. Н. Панов (1981) подчеркивает принципиальное отличие человеческого языка от языка животных: если язык человека является средством познания мира, то язык животного — лишь средство коммуникаций. Исследования Л. Германа (Herman, 1980), направленные на изучение уровня способности афалин к символизации окружающих предметов и явлений с помощью звуковых символов, находятся на первом этапе. Выяснилась лишь возможность обучения дельфинов реагировать определенными действиями на различные комбинации звуковых символов. Цель дальнейших экспериментов, как сообщает Л. Герман, установить двустороннюю связь человека с дельфином, используя звуковые символы. Таким образом, в настоящее время вопрос о возможности установления двусторонней связи с дельфинами посредством языковых символов на том же уровне, что и с шимпанзе, остается открытым. Можно надеяться, что в ближайшее время будет получен ответ на этот вопрос, уже много лет волнующий исследователей. Другим плодотворным направлением в исследовании коммуникационных возможностей китообразных является изучение биологического значения сигналов при взаимодействии животных в естественных группировках. Такие работы в настоящее время широко развертываются в отношении изучения звуковых сигналов как усатых, так и зубатых китообразных. Возможности общения с помощью языковых символов у животных тесным образом связаны с уровнем развития их интеллекта, или рассудочной деятельности. Поэтому в следующем разделе мы остановимся на некоторых экспериментах, которые могут продемонстрировать способность дельфинов к решению простых логических задач.
Изучение некоторых форм рассудочной деятельности
Выше, при описании ряда экспериментов с дельфинами, неоднократно шла речь об их интеллекте, или рассудочной деятельности, по определению Л. В. Крушинского (1977). В этом разделе мы рассмотрим проблему интеллекта дельфинов. Прежде всего остановимся на интересных работах К. Прайор. В опытах, проведенных на двух морщинистозубых дельфинах (Steno bredanensis), К. Прайор с соавторами (Pryor et al., 1969) применили для их обучения метод, ранее не использовавшийся на животных. В эксперименте вознаграждались не как обычно, отработанные, ранее сформированные движения дельфина, а только новые, до этого не поощрявшиеся. Поняв суть задачи, дельфины начинали демонстрировать не только движения, свойственные их виду, но изобретали новые, не характерные для морщинистозубых дельфинов. В проведенном исследовании авторами было показано, что дельфины могут усвоить принцип предложенной им задачи, а не только обучиться еще одной новой реакции. В этом огромное значение проведенной работы. Г. Бейтсон (Bateson, цит. по: Pryor, 1975), присутствовавший при экспериментах, определил описанное выше поведение у дельфинов как обучение высшего порядка, или «вторичное обучение». К. Лоренц (1981) оценивает результаты экспериментов с другой точки зрения. Он считает, что осмысление животными того факта, что вознаграждаться будет не какое-то определенное движение или система движений, но любое движение, которое никогда прежде не поощрялось, требует совершенно неожиданной для животного степени абстрактного мышления. К. Прайор полагает, что с помощью этих опытов ей удалось выявить присущее поведению высших животных свойство — оригинальность. Это свойство поведения дельфинов позволяет по-иному подойти к определению существа интеллекта животных. Первыми исследователями, которые поставили проблему оценки умственных способностей у дельфинов в современной литературе были А. Ф. Мак-Брайд и Д. О. Хебб (McBride, Hebb, 1948). Всесторонне и подробно проанализировав поведение дельфинов в стаде, их игры, сексуальные отношения, ориентировочно-исследовательское поведение, приручение, адаптацию к неволе, авторы пришли к заключению, что с точки зрения эмоционального поведения и действий, обусловленных внешними причинами, дельфины по своему умственному развитию занимают промежуточное положение между собакой и шимпанзе. Хотя авторы и отмечают отсутствие методов, которые могли бы всесторонне оценить развитие умственных способностей у дельфинов, они указали путь, который может привести к успеху в изучении этого вопроса. Они полагали, что нельзя использовать условнорефлекторный критерий для суждения об умственных способностях животных: не скорость обучения, а решение логических задач может служить показателем разума у животных. Тридцать лет спустя сходную точку зрения высказал X. Джерисон (Jerison, 1978). По его мнению, способность к обучению не может служить критерием для изучения интеллекта у животных, поскольку образование условнорефлекторных связей является универсальным приспособлением для всех видов животных. Он предлагает разработать тесты, с помощью которых можно было бы исследовать способность животных к абстракции. Исследователями высказываются различные мнения о возможности познания и изучения интеллекта у животных. Так, Е. О. Вильсон (Wilson, 1975), Дж. Гарсиа (Garcia, 1978) подчеркивают, что поведение животных, в том числе и интеллект, следует изучать только в рамках специфики экологии каждого вида. Авторы полагают, что вряд ли можно найти такую методику эксперимента, с помощью которой можно было бы сравнивать интеллект животных, принадлежащих к разным видам. У каждого вида животных, по их мнению, существует специфический набор поведенческих реакций, приспособленных к особенностям условий своего существования, необходимых для выживания. В то же время Л. Герман (Herman, 1980), обсуждая выявленные способности дельфинов и обезьян к оперированию случайными символами, обозначающими предметы и явления окружающей среды, полагает, что мозг этих животных преадаптирован к высокому уровню познавательных возможностей. Потенциальные возможности мозга дельфинов и человекообразных обезьян гораздо выше, чем это необходимо для существования в естественных условиях. На избыточность потенциальных возможностей мозга указывали еще А. Н. Северцов (1922), К. Лешли (Lashley, 1949) и Б. Ренш (Rensch, 1960). К. Лешли подчеркивал, что мозг животных, способных к элементарной абстракции, обладает избыточным резервом нейронов. Л. В. Крушинский (1958), рассматривая развитие рассудочной деятельности животных, полагает, что эта форма высшей нервной деятельности не может быть охарактеризована только как частное приспособление к условиям существования. Рассудочная деятельность, обусловленная сложнейшей нейронной организацией мозга, развилась как общая адаптивная функция — арогенез. Один из первых экспериментальных подходов к изучению элементарной рассудочной деятельности у животных в сравнительно физиологическом плане был осуществлен Л. В. Крушинским и его сотрудниками (см. обзор: Крушинский, 1977). По определению Л. В. Крушинского, в основе любой, самой сложной формы рассудочной деятельности животного лежит способность улавливать закономерности, связывающие предметы и явления окружающей среды, и возможность оперировать этими закономерностями. Для оценки уровня развития рассудочной деятельности у дельфинов были выбраны два критерия: способность к решению экстраполяционных задач и оценка некоторых геометрических свойств предметов (оперирование геометрической мерностью фигур). Изучение способности черноморских афалин экстраполировать направление движения исчезающих за преградой раздражителей при различных условиях их предъявления проводилось в отсеке размерами 8X10,5 м2, куда была помещена звуко- и светонепроницаемая ширма длиной 6 м, высотой 2 м, на одну треть погруженная в воду, с окном шириной 0,8 м в середине (Крушинский и др., 1982). Окно и пространство от ширмы до дна отсека были забраны сетью. Слева и справа от ширмы имелись проходы шириной около 2 м, позволявшие дельфину оплывать ширму с любой стороны. При (проведении эксперимента дельфину, находившемуся на стартовой позиции напротив окна ширмы, предъявлялся в окне натуральный (рыба) или другой привлекавший его раздражитель (мяч). После подхода животного к окну по сигналу (всплеску о воду раздражителя) и при попытке охватить приманку последняя на держателе отодвигалась вдоль ширмы и скрывалась из поля зрения животного. Таким образом, животному предлагалось решить задачу отыскания раздражителя: после получения информации о первоначальном направлении движения раздражителя экстраполировать его последующий путь за ширмой и обойти ее с соответствующей стороны. В следующем опыте приманку продвигали в противоположном направлении, а в дальнейшем направление ее перемещения меняли без строгого чередования. Были проведены 4 серии опытов, различавшихся характером раздражителей и средой, в которой их предъявляли. В экспериментах первой и второй серий приманкой служила рыба. В первой серии ее передвигали за ширмой под водой на глубине 0,2 м (22 предъявления). Дельфин правильно решил задачу, допустив только две ошибки. Во второй серии рыбу 15 раз предъявляли и продвигали за ширмой над водой на высоте 0,2 м. Это не отразилось на решении задачи дельфином, допустившим лишь одну ошибку. В третьей и четвертой сериях опытов, в которых в качестве приманки использовался мяч, передвигавшийся в 8 пробах по поверхности воды и в 10 — на высоте 0,2 м над поверхностью, дельфин во всех случаях решил задачу правильно. Таким образом, афалина четко проявила способность к экстраполяции направления скрывавшихся за преградой как натуральных, так и искусственных раздражителей, предъявлявшихся под водой и над водой (что исключало возможность их восприятия за преградой), т. е. показала высокий уровень элементарной рассудочной деятельности при решении простого варианта экстраполяционной задачи. Более сложный эксперимент, выявляющий способность животных к оперированию мерностью фигур, основан на том, что из двух фигур, предъявленных животному для выбора — объемной и плоской, только объемная фигура обладает свойством вмещаемости. Животные должны понять, что объемная фигура может быть вмещена только в объемную, но не в плоскую фигуру и в этой фигуре она может быть перемещена из одной точки пространства в другую. Опыты были проведены с двумя молодыми черноморскими афалинами-самками (Крушинский и др., 1972). Игру с мячом они часто предпочитали очередному кормлению, и поэтому в качестве приманки была выбрана их любимая игрушка — мяч.
 Рис.
3. Дельфин перед экспериментальной
установкой
Фото Л.В. Крушинского
Рис.
3. Дельфин перед экспериментальной
установкой
Фото Л.В. Крушинского
В эксперименте, который проводили обычно раз в день или через день, одному из дельфинов предлагалось решить, к какой из двух геометрических фигур (объемной или плоской) следует подплыть, чтобы найти мяч. Экспериментальная установка помещалась над водой. На высоте 1 м от поверхности воды укреплялась горизонтальная штанга, по которой от центра одновременно раздвигались плоская и объемная пара фигур. В центре установки находился непрозрачный подвижной экран, поднимая который, можно было скрыть фигуры от дельфинов. К обеим фигурам прикреплялись педали. Нажимая на них рострумом, дельфин мог повернуть их вокруг подвески на 180° и из объемной фигуры выпадал мяч. Нажатие на педаль (а в некоторых опытах — обливание фигур водой изо рта) служило индикатором выбора дельфином данной фигуры (рис. 3). Опыт происходил следующим образом. При открытой ширме дельфину демонстрировали пару фигур, и в это время экспериментатор играл с ним мячом, привязанным к леске. Затем поднимали экран, отгораживая от дельфина фигуры, и мячик прятали в объемную фигуру, отстегивая его от лески. Это занимало 30 — 40 с. Экран опускали, и, если дельфин был ориентирован строго по центру бассейна, фигуры раздвигали со скоростью 30 см/с. Дельфин мог подплывать под фигуру и рассматривать ее спереди из воды и с воздуха. При правильном решении задачи дельфин подплывал к объемной фигуре, нажимал рострумом на педаль и получал мяч. В каждом опыте ему предлагали новую пару геометрических фигур. Всего было использовано 16 пар фигур: девять плоскогранных и семь — тела вращения. Фигуры были одного цвета, фронтальные проекции которых всегда совпадали. Каждая фигура применялась в эксперименте с одним дельфином не более двух раз с промежутками между опытами в 3 — 4 недели в целях исключить обучение на фигуры данной формы. Так как фигуры предъявлялись не в воде, а над водой, то необходимой предпосылкой решения этой задачи являлась способность дельфинов к зрительному различению геометрических фигур и восприятию глубины пространства. Уже давно известно, что дельфины успешно обучаются зрительному дифференцированию изображений геометрических фигур как под водой, так и над водой (Kellogg, Race, 1966). Точно рассчитанные прыжки из воды к высоко поднятой над водой цели, способность точно забрасывать предметы, в сетку свидетельствуют о том, что дельфины хорошо воспринимают глубину пространства в воздухе. Поэтому задачу, предложенную нами дельфинам, можно было считать адекватной для их зрительного восприятия. Результаты опытов с афалинами показали, что оба животных с первого раза правильно решали задачу с геометрической мерностью фигур. Особенно интересны были два первые опыта с одной из самок по кличке Василиса, когда фигуры не имели педалей. После раздвижения фигур в обоях случаях Василиса подходила к объемной фигуре и обливала ее водой — реакция выпрашивания мяча. Поскольку афалина не получала в этих опытах мяч, то в последующих опытах она просто уходила от экспериментальной установки. Впоследствии во всех экспериментах к фигурам стали прикреплять педали. Суммарные данные опытов, проведенных с двумя афалинами, продемонстрировали явное преобладание правильных решений (из 48 опытов — 37 правильных выборов, р<0,001). Следующие доводы, с нашей точки зрения, исключают возможность объяснения выбора дельфинами объемных фигур в результате обучения. Во-первых, каждый опыт проводился с разными по форме фигурами. Во-вторых, два первые опыта с Василисой, когда фигуры были без педалей, показали, что оба раза она правильно решила задачу, не получая мяча. В-третьих, неправильные решения задачи не были приурочены к началу опытов, как это обычно бывает в опытах по обучению дифференцированию геометрических фигур (Kellogg, Rice, 1966). Ошибки в экспериментах с мерностью фигур равномерно распределялись по всей серии опытов. Появление ошибочных выборов плоской фигуры у обеих афалин совпадало с перевозбуждением животных в связи с разными причинами. Проведенные эксперименты с афалинами показали, что они способны без предварительного обучения строить программу поведения, в основе которой лежит способность к оперированию мерностью фигур, т.е. они имеют представление об элементарных геометрических свойствах предмета — вмещаемости объемных фигур.
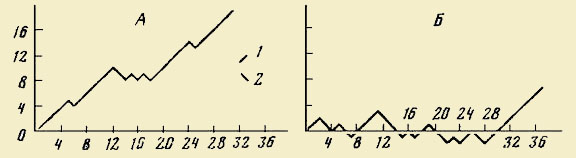 Рис.
4. Индивидуальные кривые накопления
правильных ответов
А — при решении
задачи оперирования эмпирической
мерностью фигур афалиной Василисой; Б
— во время выработки дифференцирования
геометрических фигур у афалины Леди. 1
— правильный ответ; 2 — неправильный
ответ. По оси ординат — разность между
числом правильных и неправильных
ответов; по оси абсцисс — Число (N)
предъявлений задачи
Рис.
4. Индивидуальные кривые накопления
правильных ответов
А — при решении
задачи оперирования эмпирической
мерностью фигур афалиной Василисой; Б
— во время выработки дифференцирования
геометрических фигур у афалины Леди. 1
— правильный ответ; 2 — неправильный
ответ. По оси ординат — разность между
числом правильных и неправильных
ответов; по оси абсцисс — Число (N)
предъявлений задачи
Описанную выше задачу предлагали другим животным: обезьянам (Cercopithecidae), собакам, кошкам и морским львам (Zalophus californianus). В результате проведенных опытов (Крушинский и др., 1972; Дашевский, 1972; Крушинский и др., 1976; Крушинская, Дмитриева, 1974) было показано, что только обезьяны и афалины способны к ее решению без специального обучения. В процессе многочисленных предъявлений задачи у собак удалось выработать дифференцирование объемных фигур от плоских. Однако, как показали контрольные эксперименты, даже в результате длительного обучения собаки оказались неспособными к улавливанию (абстракции) свойства вмещаемости объемных фигур, они научились лишь различать объемные и плоские фигуры (Крушинский и др., 1976). Приведенные исследования дают основание считать, что афалины способны решать элементарные логические задачи, требующие экстраполяции направления движений раздражителей и оперирования эмпирической мерностью фигур. Это свидетельствует о наличии у этих животных способности к абстракции. Следует также отметить, что эксперименты были построены не на традиционном пищевом подкреплении, а на игровом. Результаты опытов подтверждают мнение целого ряда исследователей, что дельфины (афалины) обладают высокоразвитым интеллектом, сопоставимым с таковым у обезьян.
Заключение
Приведенные в настоящей главе данные свидетельствуют о чрезвычайно высоком уровне развития мозга китообразных и весьма пластичных и сложных формах их поведения. Мощная теленцефализация китообразных не может, по-видимому, быть объяснена лишь высокой специализацией этих животных к водному образу жизни и развитой у них способностью к эхолокации, как полагают некоторые исследователи (Кесарев, 1974; Колдуэлл, Колдуэлл, 1978; Томилин, 1980), а связана в первую очередь с развитием их высших психических функций: сложных форм обучения и рассудочной деятельности. Исследование высшей нервной деятельности дельфинов методом условных рефлексов показало, что по основным видам условнорефлекторных связей дельфины существенно не отличаются от других высших наземных позвоночных. Это подтверждает известное положение в теории обучения, согласно которому в основе приобретения индивидуальных навыков у всех позвоночных лежат одни и те же процессы (Skinner, 1956; Воронин, 1969). Переделка сигнального значения раздражителей при их дифференцировании рассматривается (Bitterman, 1965) одним из показателей качественного различия способности к обучению у позвоночных животных, обладающих разным уровнем организации нервной системы. Однако применение этой методики для оценки познавательных способностей дельфинов не выявило у них существенных отличий от других позвоночных. Изучение более сложных форм поведения, таких, как имитационное научение, показало, что своего наивысшего развития оно достигает у дельфинов и шимпанзе, проявляясь в подражании орудийной деятельности человека. Подражание различным формам орудийной деятельности человека, проявляющееся в виде игр и сбора пищевых объектов, могут служить одним из убедительных доводов в пользу высокого уровня их рассудочной деятельности. Имитационное поведение дельфинов является предпосылкой для возникновения в сообществах китообразных определенных традиций, которые передаются от поколения к поколению. Способность к имитационному обучению и образование условнорефлекторных реакций на тождество сигналов (выбор по образцу) у дельфинов должна рассматриваться как выражение функций сложного обобщения. Исследование возможности выработки у дельфинов наличного и отсроченного выбора по тождеству сигналов выявило у них способность к решению этих задач того же уровня, что и у обезьян. Процесс образования выбора по тождеству сигналов осуществляется животными не на основе жесткой системы «сигнал — реакция», а путем обобщения одного или нескольких признаков раздражителя. Такая форма обобщений у животных классифицируется исследователями как «интеллектуальные формы поведения». Способность дельфинов обучаться отсроченному выбору звукового стимула-образца среди нескольких тестовых сигналов является показателем высокого уровня лабильности их оперативной памяти и надежности в реализации следовых процессов. Уровень развития оперативной памяти у дельфинов можно рассматривать как одну из предпосылок к отвлеченной символизации предметов и явлений окружающей их среды. Многочисленные исследования, направленные на изучение «языковых» способностей дельфинов, до сих пор не подтвердили гипотезы о существовании у дельфинов особых способов коммуникаций, принципиально отличающихся от сигнализаций других животных и тем более тех, по которым их можно было бы приблизить к языковому общению человека. В настоящее время исследуются возможности усвоения дельфинами звуковых символов-признаков предметов и явлений окружающего их мира, тех качеств, которые лежат в основе человеческого языка. Выяснена возможность обучения дельфинов реагировать определенными действиями на различные комбинации случайно выбранных звуков, что рассматривается как первый этап на пути к установлению двусторонней связи человека с дельфином на основе использования языковых символов. Как известно, такой путь исследований привел к успешным результатам при обучении шимпанзе азбуке жестов (ASL). Обученные по системе ASL шимпанзе могли использовать эти знаки-символы в качестве средства общения с экспериментаторами и другими обезьянами. Возможно ли будет осуществить полностью намеченную программу исследований по установлению двусторонней связи человека и дельфинов, зависит от ряда причин. Успех исследований в большой мере зависит от адекватности выбранной формы эксперимента физиологическим возможностям воспринимающих и эффекторных систем дельфина. Высокий уровень развития слухового анализатора дельфинов и способности к имитации искусственных звуковых сигналов широкого диапазона могут служить залогом успешного проведения запланированной программы исследования. Вместе с тем нельзя исключить из рассмотрения возможность существования у дельфинов систем кодирования информации, принципиально отличающихся от привычных представлений человека. Например, сложная информация может кодироваться не последовательностью символов во времени, как принято в языке человека (и, видимо, в системах общения у обезьян), а одновременной передачей набора составляющих частотного спектра звуков или каким-либо другим необычным для человека и пока нераскрытым способом. Очевидно, в этом случае навязываемая дельфину экспериментатором «человеческая» система символов может для него оказаться совершенно непривычной. И, наконец, возможность установления двусторонней связи человека и дельфина на основе «языковой» символизации зависит от того, обладают ли дельфины уровнем развития умственных способностей, необходимым для оперирования отвлеченными символами. Исследование рассудочной деятельности у дельфинов на основе решения ими простых логических задач (экстраполяция направления движения раздражителя и оперирование имперической размерностью фигур) можно рассматривать как один из этапов подхода к познанию интеллекта этих животных. Экспериментально было показано, что дельфины и обезьяны способны к решению наиболее сложных задач, требующих соизмерения пространственных свойств объемных и плоских предметов, и на основании уловленных при этом закономерностей строить адекватное поведение в новой ситуации. Рассматривая предложенные тесты как объективные параметры для оценки рассудочной деятельности животных, можно заключить, что дельфины и по этому критерию стоят на том же уровне, что и обезьяны. Обобщая данные морфологических исследований и экспериментального изучения поведения китообразных, можно сказать, что они в совокупности однозначно свидетельствуют о больших потенциальных возможностях их мозга для оперирования большим объемом отвлеченных символов.
Практичне заняття № 5. Біопсихологічні передумови виникнення психіки людини.
Мета: аналіз основних поглядів на психологічну сутність антропогенезу і особливості психіки вищих антропоїдів, вивчення теоретичних та експериментальних даних по дослідженню інтелекта антропоїдів, які розкривають найбільш суттєві особливості структури діяльності цих тварин, що мають значення для походження свідомості людини в філогенезі, а також порівняльний аналіз деяких сторін розвитку інтелекта в онтогенезі людиноподібних мавп та дитини.
Форма заняття: семінар.
Обладнання: методичні рекомендації, навчальні посібники.
План заняття
І. Проблема походження трудової діяльності.
1. Еволюція гаптичних і сенсорних функцій вищих ссавців.
2. Предметна діяльність мавп. Фирсов Л.А. «Обезьяны, берущие палки».
3. Передтрудова предметна діяльність викопних мавп.
4. Знаряддя тварин і знаряддя праці людини.
5. Резникова Ж.И. Исследование орудийной деятельности как путь к интегральной оценке когнитивных возможностей животных.
А) Визначення «орудийная деятельность» (Еллок, Гудолл, Бек, Ладигіна-Котс);
Б) Використання знарядь ссавцями;
В) Використання знарядь птахами;
Г) Експериментальне дослідження когнітивних аспектів знаряддєвої діяльності тварин;
Д) Розвиток знаряддєвої діяльності в онтогенезі;
Е) «Культурні» аспекти знаряддєвої діяльності тварин.
6. Матеріальна культура і біологічні закономірності.
ІІ. Проблема зародження суспільних відносин і членороздільної мови.
Лурия а.Р. Эволюционное введение в общую психологию. Лекция 6. Границы поведения животных и психологическая деятельность человека.
А) Три відмінності поведінки тварин;
Б) Три відмінні особливості психологічної діяльності людини;
В) Теорії виникнення свідомості людини.
Групова поведінка мавп і зародження суспільних відносин.
Демонстраційне маніпулювання.
Мова тварин і членороздільна мова людини.
Формування мови людини.
Завдання для самостійної роботи по підготовці до заняття:
Опрацювати літературу за питаннями плану семінару.
Підготувати короткий конспект-відповідь на питання семінару.
З’ясувати та записати пояснення основних ознак мови людини за Хоккетом (вокалізація, направленість, швидке затухання, повний обернений зв’язок, змінність, спеціалізація, семантичність, довільність, культурна спадкоємність, навчаємість, дискретність, заміна, відкритість/продуктивність, відхиленість, рефлексивність)
Література
Линден Ю. Обезьяны, человек и язык: Пер. с англ. Е. П. Крюковой под ред. Е. Н. Панова — М.: Мир, 1981. 272 с.
Фабри К.Э. Основы зоопсихологии. М.: Изд-во Моск. ун-та, 1993
Правоторов Г.В. Зоопсихология для гуманитариев. Новосибирск: ЮКЗА, 2001
Мак-Фарленд Д. Поведение животных. Психобиология, этология и эволюция. М.: Мир, 1988
Леонтьев А. Н. Проблемы развития психики. М.: Изд-во Моск. ун-та, 1972
Тинберген Н. Поведение животных. М.: Мир, 1969
Дьюсбери Д. Поведение животных. Сравнительные аспекты. Пер. с англ./Перевод Полетаевой И.И. – М.: Мир, 1981. – 480 с.
З.А.Зорина, И.И.Полетаева. Элементарное мышление животных. М.: Аспектпресс. 2003 г., стр. 19 – 63.
О.Т. Губко. Основи зоопсихології. Навчальний посібникик. К.: Світогляд, 2006, - 190 с.
Хейс Ники. Принципы сравнительной психологии /Пер. с англ.. – М.: «Когито-Центр», 2006. – 304 с.
Г.Г. Филиппова. Зоопсихология и сравнительная психология: учеб. пособие для студ. высш. учеб. заведений /Г.Г.Филиппова. – 2-е изд., стер. – М.: Издательский центр «Академия», 2004. – 544 с.
Хрестоматия. Сравнительная психология и зоопсихология / Сост. и общая редакция Г.В.Калягиной. – СПб: Питер.2001. – 416 с.:ил. – (Серия «Хрестоматия по психологии»).
Зорина З.А., Полетаева И.И., Резникова Ж.И. Основы этологии и генетики поведения. Изд-во МГУ, 1999. Переиздание.
Кликс Ф. Пробуждающееся мышление. У истоков человеческого интеллекта. М.: Прогресс,1983
Ладыгина-Котс Н.Н. Предпосылки человеческого мышления. М.: Наука, 1965
Рогинский Г.З. Навыки и зачатки интеллектуальных действий у антропоидов. Л., 1948.
Теми рефератів
Використання мавпами знарядь.
Вивчення свідомості у тварин.
Свідомість: походження та сутність.
Наслідування мавп.
Сигнальна система приматів.
Еволюція пластичності поведінки тварин.
Дослідження вищих форм поведінки та психіки тварин в руслі порівняльно-психологічної парадигми.
Психіка молодого шимпанзе.
Досліди В. Келера.
Дослідження Е.Г.Вацуро вищої нервової діяльності шимпанзе.
Вищі психічні функції антропоїдів.
Функції органів чуття мавп.
Психологія нижчих мавп.
Життя мавп в групах.
Експеримент подружжя Кеті та Кейт Хейс.
Особливості інтелекту шимпанзе в порівнянні з інтелектом людин
Лурия а.Р. Эволюционное введение в общую психологию. Лекция 5. Интеллектуальное поведение животных
В прошлом занятии мы дали характеристику двух типов поведения животных: один мы назвали типом сенсорной психики или инстинктивного поведения, второй – типом перцептивной психики или индивидуально – изменчивого поведения. Для первых этапов развития поведения – этапов сенсорной психики и инстинктивного поведения – характерен тот факт, что животные приспосабливаются к окружающим условиям среды, проявляя известные врожденные программы поведения в ответ на отдельные сенсорные стимулы. Одно какое – нибудь воспринимаемое свойство, например, блеск воды у комара, вибрация у паука вызывают сразу же целую сложную врожденную, закрепленную в видовом опыте программу поведения. Эта программа поведения может быть очень сложной и малоподвижной, она приспособлена к мало меняющим условиям. По этому типу построено поведения низших позвоночных и насекомых.
Второй тип поведения формируется с изменением условий существования и с развитием коры головного мозга. Он проявляется особенно отчетливо у высших позвоночных, и в частности у млекопитающих. Для этого типа поведения характерен тот факт, что животное начинает воспринимать комплексные раздражители, идущие из среды, отражать целые ситуации, регулировать свое поведение посредством субъективных образов объективного мира и приспосабливаться к меняющимся условиям. У животного на этом этапе развития комплексный раздражитель уже не просто реализует врожденные репертуары инстинктивного поведения, а вызывает акты, приспособленные к предметному миру. Поэтому ведущим на этом этапе начинает становиться индивидуальное поведение; оно проявляется в отсроченных реакциях, о которых речь шла выше, в образовании условно – рефлекторных актов, в тех навыках, которые, видно формируются на основании анализа среды, совершаемого животными.
Сегодня мы сделаем еще один шаг дальше и будем говорить к уже о наиболее сложных формах индивидуально – изменчивого поведения животных и о тех формах, которые сближаются с интеллектуальным поведением.
Рядом с инстинктивными и простыми формами изменчивого поведения у животных существует еще одна форма поведения, которая представляет собой интерес. Животные обнаруживают некоторые формы подлинно разумного интеллектуального поведения.
Возникает вопрос: в чем основа этих форм поведения? В каких формах это поведение проявляется у животных, в чем состоят его границы?
Позвольте сегодняшнюю лекцию посвятить рассмотрению этих вопросов.
Предпосылкой для инстинктивного поведения является отражение отдельных свойств внешней среды, которое действует на механизм, пускающий в ход врожденный инстинктивный акт.
Предпосылкой для сложных форм индивидуально – изменчивого поведения является перцепция, то есть отражение целых комплексных форм сложных ситуаций среды. На основе этого образа отражаемой действительности возникают индивидуально – изменчивые формы поведения.
Мы условно можем назвать эту стадию – стадией индивидуально изменчивых форм предметного поведения, то есть поведения, приспособленного к условиям внешней среды.
Что же составляет основу для наиболее сложных форм индивидуального поведения животного, интеллектуального поведения? Основу интеллектуального поведения, по – видимому, составляет восприятие сложных отношений между предметами внешнего мира. Это и есть дальнейшее усложнение форм отражения, которое ведет к появлению более интересующих нас форм поведения. Сначала животное отражало отдельные свойства и эти свойства пускали вход заложенные от природы врожденные видовые механизмы. Затем животное начинало воспринимать целые образы предметов действительности и приспосабливаться к ним; возникли индивидуально – изменчивые формы предметного поведения, которые могут быть проиллюстрированы в навыках. Но есть третья, очень существенная форма отражения, которая очень слабо выявлена у низших животных и выявляется все больше и больше у высших животных. Это – отражение не отдельных слов, не отдельных предметов и ситуаций, а сложных отношений между отдельными предметами. Она составляет основу интеллектуального поведения.
Позвольте привести несколько примеров. Мы начнем с анализа наиболее элементарных форм отражения, которые немецкий психолог Кёлер проводил с курами.
Перед курицей размещались два квадрата: один квадрат был светлосерый, а другой – темносерый. На оба квадрата были насыпаны зерна, но только на одном, более темном квадрате они лежали свободно, а на другом, более светлом, они были приклеены так, что курица, которая пыталась клевать эти зерна, не получала эффекта. Постепенно курица приучалась направляться к более светлому квадрату.
Возникает вопрос: реагирует ли курица на абсолютный цвет квадрата, или же на относительно более светлый квадрат.
Для того, чтобы ответить на этот вопрос, Кёлер предъявил курице два других квадрата – такой же темносерый и еще более темный. Прежний, более темный квадрат становился в новой паре относительно более светлым. К какому же квадрату шла курица? Оказывается она сразу же направлялась к более светлому квадрату, который был отрицательным, и игнорировала квадрат, которого прежде не было. Следовательно, она реагировала не на абсолютный цвет квадрата, а на отношение между двумя квадратами. Для того, чтобы окончательно подтвердить свое предположение, Кёлер сделал третий опыт: он дал светлосерый квадрат, который в первом опыте был положительным, и рядом с ним еще более светлый квадрат, почти белый, которого раньше не было. В этом контрольном опыте курица никогда не шла на светлосерый квадрат, и, наоборот, шла на белый, бывший ранее положительный квадрат, никогда не фигурировавший в ее прежнем опыте.
Таким образом, курица явно реагировала не на цвет, а на отношение между двумя цветами. Значит, уже на довольно раннем этапе развития существует восприятие не только свойств, но и их отношений, существует какой – то элементарный тип анализа ситуации и выделяются признаки не конкретные, а признаки, соотносящие один предмет к другому, типа разницы потенциалов.
Этот пример, ставший классическим, указывает еще на очень элементарное явление. Но можно привести другой пример восприятия отношений, который показывает эти же факты в значительно более интересных и сложных формах.
Опыт, который я сейчас приведу, принадлежит нашему советскому физиологу – профессору кафедры нервной деятельности университета Л. В. Крушинскому и называется опытом с экстраполяционным рефлексом. В данном случае речь идет тоже о восприятии отношений, но уже не пространстве, а восприятии отношений во времени. Аппарат, на котором демонстрируется указанный опыт, состоит из двух непрозрачных труб. В одну из них на глазах животного вводится на веревке приманка – кусок мяса или пачка зерен для птицы. Эта приманка движется в закрытой трубе. Животное видит, как приманка входит в трубу, видит, как приманка выходит в свободное отверстие и снова скрывается во второй трубе. Как ведет себя животное в этом случае? Как показали опыты, разные по уровню развития животные реагируют неодинаково. Те животные, которые стоят на более низком этапе развития (например, курицы) реагируют так: они бросаются на приманку проходящую через просвет и пытаются ее схватить, несмотря на то, что она прошла мимо, иначе говоря, реагируют лишь на непосредственное впечатление.
Отличие от них, животные, которые стоят на более высоком уровне, дают совершенно иную реакцию: они смотрят на приманку, проходящую через просвет, затем бегут к концу трубы и дожидаются, когда приманка появится у открытого конца.
Из птиц так делают хищные; так всегда делает кошка и собака.
Значит все эти животные реагируют не на непосредственное впечатление, а производят экстраполяцию, то есть учитывают, где появится данный предмет, если он движется. Они предвосхищают движение предмета, и в этом предвосхищающем поведение состоит особенность высокоразвитых животных.
Значит наряду с реакцией на непосредственное впечатление у высших позвоночных существует известный тип предвосхищающего поведения, то есть реакции с учетом отношений между тем, где предмет находится в данный момент и где он будет в последующем.
Это поведение уже является типом разумного поведения, которое резко отличается как от инстинктивного, так и обычных, более элементарных форм индивидуально – изменчивого поведения.
Эти формы поведения и стали предметом изучения целого ряда психологов и физиологов. Больше всего в изучении этого поведения стал немецкий психолог Кёлер, о котором я уже упоминал выше; много сделал и американский психолог Черкс и грузинский психолог Бериташвили.
Очень много интересных фактов было добыто в этом отношении и советским физиологом Протопоповым. Сейчас я расскажу вам о некоторых из этих исследований.
Первая группа исследователей вот такого интеллектуального поведения – это методика так называемого обходного пути. Она заключается в следующем.
Животное помещали в ящике, у которого одна стенка состоит из решетки. Перед решеткой располагается приманка. Приманка расположена так, что непосредственно животное не может его достать.
Как ведут себя в этом случае животные, стоящие на ступенях эволюционной лестницы?
Я начну с примера, приведенного академиком Беритовым. Курица, помещенная в только что описанную загородку, воспринимает зерна и просто бьется о сетку, никак не может отвлечься от непосредственного образа приманки; корова в аналогичных условиях довольно вяло стоит, тыкается мордой в перегородку и не делает никаких попыток обойти загородку. А вот собака ведет себя совершенно иначе; она несколько раз пытается достать приманку непосредственно. затем делает совершенно обратное – бежит от приманки, огибает загородку и берет приманку. Конечно так же поступает и обезьяна.
Сложное поведение, выступающее в последнем случае, разбивается на три фазы; первая фаза – фаза первоначальных проб, попыток; если эти попытки удаются, животное тормозит свои непосредственные реакции. После этого начинается вторая фаза: животное начинает исследовать ситуацию, и, когда это исследование заканчивается, оно совершает последний акт – бежит не к приманке, а от нее, и это, казалось бы, бессмысленное поведение получает свой смысл только потому, что в его результате животное достигает истинной цели.
Таким образом, если на первых этапах филогенетической лестницы поведение носит элементарный, непосредственный характер, если оно определяется непосредственным восприятием отдельного свойства, сигнала (блеск для комара, вибрация для паука) или комплексным отражением непосредственно – воспринимаемого предмета(когда животное, например, в опыте отсроченных реакций, бежит к ящику, в котором скрыта приманка), то здесь поведение животного принимает уже сложный характер и начинает состоять из цикла последовательных взаимно подчиненных звеньев. В таких случаях мы, несколько упрощая факты, говорим, что поведение животного приобретает многозначных характер; оно начинает с непосредственных попыток, включает ориентировочно – исследовательскую деятельность и завершается рядом организованных, вспомогательных операций, в результате которых оно достигает цели. Некоторые авторы с основанием говорят, что действие здесь разбивается здесь на три фазы: непосредственные пробы и ориентировка в окружающей среде, в этой первой фазе создается ориентировочная основа будущего действия и формируется общая схема тех путей, которые могут достичь цели; исполнительной операции, в течение которой животное осуществляет выработанную схему действия, и поэтому третья фаза, во время которой животное сличает достигнутый эффект с искомым намерением, и либо заканчивает действие (если оно согласуется с исходным намерением), либо продолжает его (если такой согласованности не наступает) /П. К. Анохин/.
Недаром исследователи называют эту последнюю фазу фазой акцепции действия и считает ее важнейшем звеном саморегулирующего поведения животных.
Такой сложный характер действия, имеющего предварительно ориентировочную основу и распадающего на ряд последовательных взаимноподчиненных операций, можно назвать структурой интеллектуального поведения.
Некоторые американские авторы применяют другой термин, который взят ими из теории игр и нередко применяется для анализа сложных форм деятельности. Они говорят, что всякое интеллектуальное действие имеет стратегию и тактику. Стратегией является общая схема действия, тактикой – существование этого акта с помощью вспомогательных операций.
Начиная с самых элементарных форм интеллектуального поведения животного и кончая самыми сложными формами интеллектуального поведения человека, интеллектуальные акты всегда отличаются наличием такой ориентировочной основы действия, такой стратегии и тактики.
Позвольте мне сейчас обратиться к некоторым классическим опытам, в которых изучалось интеллектуальное поведение животных. Эти опыты были проделаны Кёлером, и получили известность как элементарнейшие опыты с употреблением орудий. Употребление орудий есть всегда типичное интеллектуальное действие. Ведь для того, чтобы употребить орудие и обратиться, например, к молотку, с помощью которого человек отбивает кусок камня, которым он затем пользуется, он вынужден совершать акт, имеющий известную стратегию и распадающий на ряд операций. Поэтому употребление орудий может быть типичным примером не только простого предметного действия, но и сложного интеллектуального акта. В силу этого, Кёлер и обратился к исследованию того, могут ли у обезьян быть в употреблении орудия, иначе говоря – способны ли обезьяны к более сложной форме интеллектуального действия.
Опыт был поставлен следующим образом.
Первый простой опыт. Обезьяна в клетке, передняя стенка решетка. вне клетки расположена приманка, которую обезьяна никак не может достать рукой; сбоку лежит палка, которая расположена ближе, чем приманка. Может ли обезьяна воспользоваться палкой для того, чтобы достать приманку? Опыты показали следующее: сначала обезьяна всячески пытается изловчиться, чтобы достать приманку рукой – стратегии еще нет, есть прямые попытки достать приманку рукой; затем, когда эти попытки оказываются тщетными, она останавливается и наступает следующий этап: обезьяна оглядывает ситуацию, берет палку, подтягивает ее к себе и палкой достает приманку.
Второй опыт носит более сложный характер. Приманка находится еще дальше. С одной стороны лежит короткая палка, которой приманку ни как не достанешь, а с другой стороны, немного дальше – длинная палка, которая годна для того, чтобы достать приманку. Исследователь ставит перед собой вопрос: может ли обезьяна сначала взять короткую палку, а затем с помощью короткой палки достать достать длинную и с помощью длинной палки достать приманку? Оказывается, что для обезьяны эта задача гораздо сложнее, но все же доступна. Обезьяна очень длительно делает непосредственные попытки достать приманку, истощается, затем оглядывает поле, и, как описывает Кёлер, берет первую палку, и ее помощью достает вторую, и второй палкой – приманку. Очевидно, в этот момент, говорит Кёлер, у обезьяны возникает схема будущего действия, схема решения и общая стратегия действия. Кёлер говорит даже, что обезьяна испытывает нечто вроде того, что испытываем мы, когда говорим «ага, поняли», и называет этот акта «ага – переживем».
Третий опыт еще более сложный. Он построен так же, как и второй опыт, с той только разницей, что палка находится в разных полях зрения. Когда обезьяна глядит на одну палку, она не видит второй, когда глядит на вторую – не видит первой. В этом случаи задача для обезьяны оказывается почти неразрешимой. Нужно, говорит Кёлер, чтобы обе палки и приманка были в одном поле зрения, чтобы их отношение могло быть наглядно воспринято. Только при этих условиях, если обезьяна наглядно воспринимает отношение всех трех предметов, у нее может появиться наглядная гипотеза решения и возникает соответствующая стратегия.
Последняя серия опытов Кёлера, которую мы упомянем – это тоже опыты с употреблением орудия, но эти опыты несколько иначе построены. Высоко на крючке подвешена приманка, обезьяна должна ее достать; на площади разбросаны ящики.
Решение заключается в том, чтобы взять ящик и поставить его на другой ящик, сконструировать башню и достать плод.
Поведение обезьяны носит приближенно такой характер: сначала бесконечное прыганье к приманке, попытка непосредственно, прямо достать ее. В таких попытках обезьяна обнаруживает удивительную виртуозность. После этого обезьяна утомляется, как описывает Кёлер, осматривает ситуации и затем вдруг обращается к ящикам. Она берет ящик, ставит один ящик на другой, причем, интересно, что она ставит ящики иногда так, что зрительно составляют одно вертикальное целое, но не могут удержаться друг на друге. Затем обезьяна стремится влезть на ящики; иногда ей это удается благодаря ее ловкости, и тогда она достает высоко подвешенный плод. Следовательно, и в этом опыте обезьяна может употреблять известные орудия для того, чтобы достичь цели.
И здесь ее поведение разбивается наряд последовательных звеньев: сначала беспомощные попытки и какая – то ориентировка в окружающей действительности, затем временный отход от непосредственного решения цели, торможение непосредственных попыток и обращение к орудиям – к ящикам, и, наконец, исполнение той схемы, которая была найдена обезьяной.
До сир пор речь шла об употреблении готовых предметов в качестве орудий. Возникает, однако вопрос: может ли обезьяна изготовить орудие? Для решения этого вопроса Кёлер поставил последняя серию опытов.
Обезьяна ставилась перед задачей достать далеко лежащий предмет; ей давались две палки, но каждая из этих палок была слишком коротка, чтобы достать нужную цель. Единственное решение заключалось в том, чтобы вставить одну палку в другую, удлинить палку и с ее помощью достать цель. Следовательно, здесь речь идет не об операции употребления готовых орудий, а об известной операции синтеза, изготовление нового орудия. Доступна ли обезьяне такая операция?
Такие опыты проделывал, как Кёлер, так после него и другие исследователи. Эти опыты показали, что трудности, возникающие перед обезьяной, были здесь значительно больше. Если для первого опыта ее нужны были десять минут, то для этого опыта ее нужны были часы. Как показал Кёлер, обезьяна все – таки иногда могла изготавливать орудия, синтезируя одно орудие из двух элементов. Остается последний вопрос – может ли обезьяна хранить орудия или делать орудия впрок? Ведь когда мы делаем орудие, мы их делаем заранее с тем, чтобы использовать их в нужном случае. Даже первобытный человек готовит нож или молоток впрок и применяет их лишь тогда, когда в этом возникает необходимость.
Может ли обезьяна ориентироваться на будущее и создавать такие орудия заранее, впрок?
Оказывается она не может это сделать. Если опыт на синтетическое поведение составления орудия давал относительно ясные положительные выводы, то последний опыт на делание орудий впрок и сохранения орудий – дал резко отрицательные результаты. Обезьяна, применив палку, никогда не сохраняет ее, она ее выбрасывает. Даже составив палку, она никогда не хранит ее как орудие, которое в дальнейшем может быть использовано. Значит, орудие применяется обезьяной только в непосредственной наглядной ситуации. Если поведение обезьяны в этой непосредственной наглядной ситуации оказывается близким к интеллектуальному поведению человека, то оно резко отличается от него в двух отношениях; обезьяне гораздо труднее комбинировать орудия в одно целое и обезьяне совершенно недоступно создание орудий впрок и хранение орудий, как постоянных средств для того. чтобы осуществлять те или иные действия. Таким образом можно сказать, что у обезьяны, у которой есть интеллектуальное поведение в наглядной ситуации, но нет интеллектуального поведения, близкого к человеческому, в том отношении, что она никогда не имеет дело с орудиями, в которых известный способ решения задачи и который сохраняется для известного употребления. Орудие возникает у обезьяны в процессе ее непосредственного действия и сразу же исчезает. Она живет в мире приманок, действующих на ее условия среды, но в отличие от человека, не живет в мире орудий, в мире средств, с помощью которых эти условия могут быть достигнуты.
Я изложил эти факты, касающие интеллектуального поведения животных.
Какие же механизмы лежат в основе описанных мною форм поведения? Если в описании этих форм поведения отдельные ученые согласны друг с другом, то в интерпретации, в теоретическом анализе они сильно расходились. Я позволю себе остановиться на четырех теориях, которые показывают, какие пути проходила наука в интерпретации полученных результатов.
Первая теория принадлежит американскому психологу Иерксу. Ее называют теорией инсайт, что по-русски означает озарение, проникновение, изобретение.
Теория эта заключается в том, что обезьяна отличается от остальных животных тем, что у нее есть признаки разума; она пытается решить задачу, она чувствует бесплодность своих попыток и вдруг ее озаряет мысль. Эта мысль и есть инсайт; это и лежит в основе ее интеллектуального поведения.
Как оценить эту теорию? Эта теория была хороша тем, что она обратила внимание исследователя на те факты, которые раньше игнорировались, то есть на те факты, сближающие поведение животного с человеческим поведением.
Но отрицательная сторона этой теории заключается в том, что, описывая некоторые факты, она не раскрывает их механизмов, но просто описательно костатирует нечто близкое к интеллекту, давая в конечном счете идеалистическую интерпретацию.
Второй теорией является теория ряда американских авторов, различных по своим подходам, но объединяющихся тем, что они отличают вообще интеллектуальный характер за этими описываемыми мной фактами; они считают, что все эти факты, не говоря о назначении интеллектуального у животных, что они говорят только о сложных формах навыков, встречающихся у животных, к этим авторам относится Торндайк и из современных – очень интересный крупный американский психолог Скиннер. Скиннер провел ряд опытов, по своей структуре очень близких, к тем, что делал Кёлер. Эти опыты его заключаются в следующем.
Животное помещалось в специальный ящик, дверь которого запиралась и приманка оказывалась за дверью. Единственный способ достать приманку состоял в том, чтобы нажать на педаль или рычаг, тогда дверь открывалась и животное получало приманку. Скиннер назвал эти опыты «опыты с инструментальным обуславливанием». Он прослеживал, что вырабатывается это, казалось бы разумное поведение, при котором животное нажимает на рычаг и открывает дверь, и пытался объяснить это условно – рефлекторными механизмами. В тем, по его мнению, заключались эти механизмы? Животное сначала проявляло беспорядочные пробы, но когда случайно его движение попадало на рычаг и дверь отворялась, это движение закреплялось; в результате возникла большая вероятность проявления такого движения по сравнением с движением, которое не подкреплялось. Значит, говорил Скиннер, целесообразное движение, с помощью которого животное решало свою задачу, являлось результатов подкрепления случайных проб. Как по теории Дарвина, случайные мутации закрепляются, если они оказываются биологически целесообразными, так и нужные движения, появляясь случайно, закрепляются, если они подкреплены приманкой, иначе говоря, если они оказываются целесообразными.
Что характерно для этой теории? То, что она отрицает интеллектуальный характер этих форм поведения, и, что нет принципиальной разницы между образованием навыка и интеллектуальным поведением; и тут и там сторонники этой теории видят отбор удачных движений и закрепление тех из них, которые подкрепляются.
Как оценить эту теорию? Она являлась безусловным прогрессом по сравнению с прежними теориями, потому, что она имеет аналитический характер и исходит не из антропоморфических представлений, то есть не из кажущей близости этого поведения к поведению человека, но подвергала анализу составные элементы поведения, она, как бы исторически, подходит к сложным формам поведения. Тем самым, эта теория представляет интересный прогресс, ибо что может быть научно прогрессивным, чем пытаться отходить от простого впечатления, от фактов и заменять впечатление анализом фактов?
Однако эта теория имеет и отрицательные стороны. Она просматривала самое существенное, что есть в интеллектуальном поведении животного и что отличает вот эту форму двухфазного приспособительного поведения от более элементарных форм поведения животных.
Очень существенную критику этой теории дал русский физиолог Протопопов. Он тоже исходил из этой аналитической теории, принципиально такой же, как и теория Скиннера, исследовал животных в аналогичных ситуациях. Свои опыты он назвал «стимул – преграда», потому, что стимул или известная цель давались им в условиях преграды. Но Протопопов показал,, что те движения, из которых отбираются нужные, вовсе не такие случайные. Движения животных, которые находятся в ситуации стимул – преграда, вовсе являются не случайными, не хаотичными. Животные отражают предметное строение ситуации, и этот предметный характер ситуации направляет их движение. Поэтому нажим на педаль, которой отворяется дверь, является не случайным, а результатом действенного анализа, в котором движения животного уподобляются ситуации, в которой оно находится.
Мы дали краткую характеристику этой теории, оценив ее положительные и отрицательные стороны, она аналитична и научна по своим исходным задачам, но она механистична потому, что недооценивает своеобразия форм интеллектуального поведения животных, и тем самым оставляет вопрос не до конца раскрытым.
От этих двух теорий отличается третья теория, которая принадлежит тому же автору, начавшему основную работу над инстинктами обезьян – Кёлеру. Эта теория называется перцепторной теорией интеллектуального поведения или структурной теорией.
Интересно, что Кёлер принадлежит к числу тех психологов, которые всю жизнь специально занимались механизмами восприятия.
Эту теорию структурного характера восприятия Кёлер и использовал для объяснения интеллектуального поведения обезьян.
Как вы помните, наблюдая поведения обезьяны, поставленной в сложные условия, Кёлер видел, что она сначала пытается решить задачу непосредственно, рвется к прямому решению цели, затем истощается и неподвижно сидит и смотрит на ситуацию. Вот в этот момент рассматривания ситуации, когда тормозятся ее движения и обезьяна смотрит на окружающую ситуацию, и, по мнению Кёлера, возникает «ага – переживание» или решение. Корни этого решения задачи, по Кёлеру, нужно искать не в явлении «инсайта», а в закономерностях целостного восприятия обезьяны. Обезьяна смотрит на цель, на палку, и эта цель и палка как бы объединяются в ее наглядном восприятии; этим замыкается известная целостная связь, в восприятии создается наглядная ситуационная связь между палкой и целью. Если эта связь замкнута в восприятии, она оказывается замкнутой и в действии. Ключом к интеллектуальному поведению обезьяна являются, по Кёлеру, не законы ее движения, а законы ее восприятия. Принципиально говоря, законы интеллектуального поведения, по Кёлеру, несколько не отличаются от законов целостного восприятия, и именно это, по мнению Кёлера, является ключом к механизмам, стоящим в основе интеллектуального поведения обезьяны.
Теория Кёлера описана им в очень интересной книге «Интеллектуальная деятельность высших обезьян», переведенной и на русский язык.
Что имеется положительного в этой теории? То что теория описывает факты несравненно тоньше и лучше, чем описывает механистическая теория, и останавливается на своеобразии фактов, мимо которых проходила механистическая теория.
Что же касается недостатков данной теории? То, что она считает, что интеллектуальных акт проявляется у обезьяны не в действии, а в пассивном восприятии. Именно поэтому она гипотетична, недоказуема; она предполагает, что наиболее сложные психические акты совершаются у обезьяны в тот момент, когда она ничего не делает и просто смотрит; однако очень сомнительно, что именно в этот пассивный момент восприятия происходят сложнейшие процессы, которые на ранних этапах развития должны быть непосредственно связаны с активной деятельностью.
Исходя из этого, была выдвинута четвертая теория. Она принадлежит немецкому психологу Бюлеру и исходит вот из каких соображений.
Бюлер не отрицает, что те факты, которые описал Кёлер, правильны, но считает трактовку этих фактов недостаточной. По его мнению, Кёлер описывает обезьяну, которая рассматривает внешние объекты так, как будто бы она никогда в жизни раньше с этими ситуациями не встречалась. Однако, справедливо говорит Бюлер, у обезьяны имеется богатый прошлый опыт. Обезьяна – это животное, которое живет на деревьях. Ей случалось в прежнем опыте добывать плод, пригибая к себе ветку, или пользоваться веткой для того, чтобы сшибать плод. Нельзя, говорит Бюлер, объяснить интеллектуальное поведение обезьяны, не учитывая исторически ее прежний опыт. Перенос этого прежнего опыта неизбежно должен участвовать в решении сложных задач у обезьяны. Обезьяна выделяет выделяет из своего прежнего опыта то, что имело успех, и эти элементы прежнего опыта включаются в ситуацию решения данной задачи.
Если Кёлер объяснил интеллектуальное поведение обезьяны законами восприятия, то Бюлер объясняет интеллектуальное поведение обезьяны законами памяти, то есть законами воспоминания элементов прежнего опыта, переносом прежнего опыта в новую ситуацию.
Ценным и положительным в теории Бюлера является экологический подход к жизни обезьян, то есть анализ интеллектуального поведения проводит, исходя из прежнего биологического опыта данного вида. В этом отношении теория Бюлера стоит значительно выше предыдущих теорий.
Слабой стороной этой теории является недостаточный анализ, которому Бюлер подвергает факты, с которыми он имеет дело. Бюлер только предполагает факт переноса, который должен иметь место в поведении животного, но еще не предполагает опытов, которые бы изучали эти механизмы достаточно подробно.
Я изложил вам четыре теории, авторы которых пытаются подойти к интеллектуальному поведению высших животных.
Как подходим мы, советские психологи, к этой проблеме, пытаясь взять положительные стороны перечисленных теорий и избавиться от их отрицательных сторон?
Я позволю себе кратко сформулировать наш подход в нескольких положениях.
Первое положение. Описание качественных особенностей интеллектуального поведения животных – имеет большое значение для понимания структуры наиболее сложных форм поведения.
Интеллектуальное поведение животных никак не может быть понято как закрепление случайных, беспорядочных попыток решения задачи.
Именно поэтому каждая из упомянутых мною теорий сделала свой существенный вклад в анализ интеллектуальной деятельности животных. Теория инсайта Иеркса обратила внимание на формы поведения животных, которые выходят за пределы простых, постепенно формирующихся навыков, но еще идеалистически трактовала эти факты, неправомерно сближая поведение животных с поведением человека и не давая анализа этих форм поведения. Механистические теории попытались проанализировать механизмы, лежащие в основе образования интеллектуальных операций, но игнорировала специфику этих высших интеллектуальных форм поведения, страдая грехом сведения качественно новых форм к более элементарным. Они игнорировали качественно новые формы поведения, возникающие лишь на высших этапах эволюции и находили у обезьян такие же законы выработки навыков, как и у мышей, не представляя, что в процессе эволюции возникают качественно новые формы поведения, принимающие характер интеллектуальных. Опыты Кёлера впервые были подвергнули интеллектуальное поведение животных точному экспериментальному анализу – и за это мы благодарны ему; однако, Кёлер неправомерно отрывал процесс интеллектуального приспособления животного от его активной деятельности, недоказательно считал, что корни интеллектуального поведения можно искать в момент, когда животное замолкает, тормозится и ничего неделает.
Наконец, Бюлер правильно экологически подходил к анализу истории формирования интеллектуальных процессов, но еще не давал их настоящего структурного анализа.
Вот наша оценка ряда зарубежных теорий интеллектуального поведения животных.
Из чего мы исходим, когда пытаемся научно подойти к интеллектуальному поведению животных? Прежде всего мы исходим из того, что всякая форма приспособления животного к окружающим условиям среды есть известная активная деятельность, протекающая, однако, по рефлекторным законам. Иначе говоря, животное имеет известные потребности, оно отражает условия внешней среды, хранит программу своего прежнего поведения, осуществляет известные направленные пробы, корректирует эти пробы, если они не дают нужного эффекта, но всегда приспосабливаются к условиям внешней среды в конкретной практической деятельности, иначе говоря, в конкретных двигательных актах. Животное никак не решает раньше что – то в уме с тем, чтобы потом осуществить это в деятельности, – оно пытается решить задачи в процессе активного приспособления к среде.
Интересно, что исходя из этого, И. П. Павлов, ничуть не отрицая наличия интеллектуальных форм поведения обезьян, назвал мышление обезьяны ручным мышлением. Итак, первое положение нашего подхода к интеллектуальной деятельности заключается в том, что всякое приспособление к внешней среде осуществляется ими в процессе активной рефлекторной деятельности.
Второе положение заключается в признании того, что строение этой активной деятельности на разных этапах эволюции неодинаково и что только с точки зрения эволюции можно подойти к формированию у высших животных интеллектуальных форм поведения.
На начальных этапах эволюции мы имеем дело с непосредственным изменением протоплазмы под влиянием внешних условий. Это медленные кратковременные формы пластичного изменения протоплазмы, которые осуществляются постепенно, сохраняя лишь краткие промежутки времени.
На следующим этапе – сенсорной психики приспособление к внешним условиям среды пускается в ход отражением отдельных сигналов или отдельных свойств, действующих на среду и вызывающих программы инстинктивного поведения.
На дальнейших этапах, о которых шла речь на последних лекциях, сенсорное, элементарное отражение внешней среды занимается сложным перцептивным комплексным отражением, и животное начинает вырабатывать индивидуально – изменчивые формы поведения, приводя его в соответствии с внешним предметным миром. Именно, на этом этапе и возникает предметная деятельность животных; в зависимости от того, с какими предметами животное имеет дело, возникают те формы навыков, о которых мы говорили в прошлой лекции.
Эта выработка приобретенных форм деятельности, соответствующих предметным условиям среды, является существенным этапом для развития сложных индивидуальных форм поведения. Механизм этих форм поведения и изучается физиологией высшей нервной деятельности.
Что же возникает на еще более сложных этапах эволюции?
Как мы говорили выше, здесь возникает отражение не только отдельных свойств или предметов, но и целых отношений между предметами. Но это отражение отношений между предметами возникает не пассивно, а всегда в процессе деятельности деятельности животного.
Эта деятельность, в процессе которой возникает отражение сложных отношений, называется ориентировочной или ориентировочно исследовательской деятельностью. Эта ориентировочно – исследовательская деятельность еще очень мало выражена на низших этапах развития позвоночных; она начинает занимать все большее место лишь на позднейших этапах эволюции, развиваясь вместе с формированием сложных структур коры головного мозга.
У собак, которые охотятся за дичью, эта ориентировочная деятельность приобретает уже достаточно сложную форму; но в особо сложную форму она разрастается у обезьян. Если наблюдать за обезьяной в клетке, то можно видеть, что она ни минуты не сидит в покое. . Обезьяна, как говорит Павлов, занимается все время «бескорыстной» ориентировочной деятельностью: она имеет дело не только с вещами, которые годны для пищи, которые можно сунуть в рот и съесть, но и с любыми вещами, которые ощупывает, обнюхивает, которые практически анализирует, в которых ориентируется.
Поэтому не простое впечатление от предметов, а эта ориентировочно – исследовательская деятельность является той основой, из которой вырастает интеллектуальное поведение.
Павлов ставил целый ряд опытов над обезьянами. Один из его опытов заключается в следующем. Обезьяне давался ящик, в котором была скрыта приманка, в ящике имелась треугольная прорезь. Обезьяне давались палки с разными сечениями – круглым, квадратным, треугольным. И Павлов смотрел, как она выбирает палку, которой можно открыть замок, соответствующий треугольному отверстию. Такого типа опыты показывают, что сначала обезьяна делает попытки не специфические, затем начинает обнюхивать, ощупывать предметы и, наконец, в процессе ориентировочной деятельности возникает правильный выбор адекватного предмета.
Имеются все основания думать, что возникновения правильного решения есть результат не предварительного умственного действия у обезьяны, а происходит в процессе «ручного мышления», то есть в процессе непосредственной ориентировочной деятельности. Исходя из этого, мы будем искать подходы к интеллектуальному поведению обезьяны не просто в восприятии, не просто в памяти, но в ориентировочно – исследовательской деятельности обезьяны, в процессе которого выделяются, сопоставляются нужные признаки. Если эти признаки соответствуют нужным, действие оказывается успешным и оно кончается, а если они оказываются не соответствующими нужным, действие продолжается.
Эта схема построения и привела физиологов и психологов к новой концепции поведения животных. Эта концепция отличается от классической концепции, трехчленной рефлекторной дуги (стимул, внутренняя реакция и переработка). Адекватной для объяснения поведения животных является не трехчленная дуга, а четырехчленная схема, или схема сложного рефлекторного кольца, в которую включается сложная ориентировочно – исследовательская деятельность животного.
Известные задачи, поставленные средой, вызывают у обезьяны сложную ориентировочную деятельность, приводящую к ряду проб; эти пробы выливаются в ряд операций, которые постепенно сравниваются с исходными условиями. Если выработанные пути согласуются с исходными условиями – задача разрешена, действие прекращается. Если такая согласованность не имеет место и действие не соответствует исходным явлениям – оно продолжается. В результате возникает такой сложный процесс: проба, операция, сличение и выход, если пробы соответствуют исходным. Американское психологи называют эту схему Т – О – Т – Е, то есть это проба – операция – проба – результат. Видный советский физиолог, профессор Анохин называет – механизмом акцептора действия. Соответственно этому последнему представления, определенные условия внешней среды ставят перед животным соответствующую задачу, и у животного возникает известный образ того, как нужно было бы решать задачу. Если действие не соответствует намеченной цели, возникает рассогласование операции и исходного измерения; в этом случае в мозгу идут обратные сигналы о несоответствии действия, и действие снова продолжается. Если же действие оказывается согласованным с исходным намерением – попытки дальнейших проб прекращаются.
Аппарат намерения, или образа, или акцептора действия, иначе говоря, механизм сличения намерения и данной операции, является важнейшим элементом для любых и особенно сложнейших саморегулирующих форм поведения, и его нужно иметь ввиду, анализируя сложное строение интеллектуальной деятельности.
Таким образом, возникает четырехчленная структура сложнейшего поведения животного, раздражитель – центральная переработка, совершающаяся в процессе ориентировочной деятельности – создание известной схемы решения – действие, адекватное существованию нужной задачи.
Так возникает сложнейшее поведение как саморегулирующаяся система. С этим нам с вами придется сталкиваться еще много раз, поэтому мы сегодня ограничимся указанием на эту сложную структуру высших форм поведения животных.
Лекция 6. Границы поведения животных и психологическая деятельность человека
Материал сегодняшней беседы я рассматриваю как чрезвычайно важный. Он должен послужить введением к целому большому циклу занятий, которому практически будет посвящен весь оставший курс психологии.
Вопрос, который я хочу осветить сегодня, заключается в объяснении того, в чем состоят особенности человеческого сознания и чем строение психологической деятельности человека отличается от структуры поведения животных.
Для того, чтобы подойти к этой теме, резюмируем сначала в самых кратких форматах то, что мы знаем о поведении животного и попробуем, исходя из этого, ответить на вопросы о том, каковы границы, за которые не может перешагнуть даже самая высокоразвитая форма поведения животных.
Мы осветили в течение ряда лекции основные черты, характеризующие поведение животных. Мы показали, как от инстинктивных, наследственно – программных форм поведения животное постепенно переходит к индивидуально – изменчивой форме приспособления, в который сигнал, полученный из среды, не просто запускает врожденную программой деятельность, а в которой животное программирует все свое поведение соответственно восприятию целых ситуаций и формированию субъективного образа объективного мира.
Мы показали, что с развитием ориентировочной деятельности и сложных форм отражения соотношений между вещами у животных формируются наиболее высокие формы интеллектуального поведения. Мы показали, что эти формы интеллектуального поведения имеют трехфазный характер. Первая фаза является предварительная ориентировка животного в условиях среды, предварительное ознакомление с задачей анализа существенных элементов ее; второй фазой – исполнение той гипотезы или той программы, которая возникла в процессе предварительной ориентировки; третьей фазой действия – сличение полученных результатов с исходными намерением и коррекция ошибочных действий, если они имеют место.
Мы показали, что это сложнейшая интеллектуальная форма поведения животных осуществляется в процессе действенных проб, который, как было показано в трех последних лекциях, имеет характер отражения известных сложных форм отношений между предметами, которые животное воспринимает, выделение возможных способов решения, торможение побочных неадекватных способов решения и выработка тех программ поведения, которые приводят к нужной цели.
В последний раз вам был сообщен очень существенный экспериментальный материал, который показывает, что животные могут не только употреблять известные средства или «орудия» для решения практических задач (хотя отличие их от настоящих орудий мною будет освещено после), но могут при определенных условиях и составлять орудия, например, составить одну длинную палку из двух или даже выделять эти орудия из однородного материала (выгрызать или выламывать нужную палку из деревянного диска для того. чтобы после употребить эту палку как средство достать приманку). Значит, животное может не только употреблять готовые средства, но может выделять из окружающей среды нужные средства, причем, что самое интересное, такое выделение орудий становится настолько самостоятельной формой деятельности, что обезьяна может часами, не отвлекаясь, заниматься попытками выделить нужное орудие (например, выломать палку из очень прочного диска), чтобы после того, как орудие выделено, непосредственно применить его как средство достать приманку.
Следовательно, в данном случае деятельность животного носит уже совсем не интеллектуальный характер, не характер просто элементарного условного рефлекса или привычного, удержавшегося из прежнего опыта навыка, – она представляется сложной ориентировочной деятельностью, в процессе которой выделяется известная программа, животное подчиняется этой программе, этому образу будущего средства, которое оно должно выделить из имеющегося в его распоряжении материала. Все эта создает доминанту у животного, иногда оттесняет из его непосредственного внимания даже конкретную цель, которую животное на время забывает, пока не выделит средство, позволяющее ему получить приманку.
Таким образом, на высшем этапе высшие животные с развитием коры головного мозга, с мощными зонами, обеспечивающими синтез сигналов из разных рецепторных зон, с развитой синтетической деятельностью, могут выполнять очень сложные формы поведения, программировать свое поведение сложными, возникшими в ориентировочной деятельности образами.
Все это может создать впечатление, что грани между животными и человеком стираются, и животные могут обеспечивать такие сложные формы интеллектуального поведения, которые начинают быть очень похожими на сложные интеллектуальные, разумные формы поведения человека.
Однако это впечатление, которое с первого взгляда может показаться очень очевидным, оказывается неправильным. существует ряд коренных отличий в поведении животного от собственного поведения человека.
Позвольте сейчас остановится на вопросе о границах поведения животного.
Рассмотрим внимательно поведение высших человекообразных обезьян, не столько для того, чтобы сблизить их поведение с поведением человека, а столько для того, чтобы выяснить, что отделяет их от поведения человека. Попробуем, следовательно, описать известные поведения даже самых развитых животных.
Первое отличие заключается в том, что поведение животного всегда осуществляется внутри определенной биологической деятельности, внутри определенного биологического мотива.
Животное никогда не делает ничего, что не обслужило бы известную биологическую потребность, что выходило бы за рамки определенного биологического смысла. Всегда всякая деятельность животного в конечном итоге либо мотивирована сохранением индивида, либо мотивирована продолжением рода. Деятельность животного либо обслуживает инстинкт питания (то есть она делает что – нибудь, чтобы получить пищу), либо инстинктом самосохранения (оно производит действие, чтобы спасти себя от опасности), либо инстинктом продолжения рода. Значит, всякая деятельность животного, сколько бы она не была расчленена, всегда осуществляется в пределах биологически осмысленной деятельности.
Я не могу забыть, как примерно 35 лет тому назад мне удалось в Голландии слышать доклад одного известного психолога – Гельба, который сформулировал парадоксальное, но очень правильное положение. Он сказал: «Животное не может ничего делать бессмысленного, делать бессмысленное способен только человек!»
Если вдуматься в эту формулу, можно увидеть, что она таит в себе действительно правильное рассуждение.
Животное не может делать ничего, что выходило бы из пределов биологического смысла, в то время как человек 9/10 своей деятельности посвящает актам, не имеющим прямого, а, иногда, даже и косвенного биологического смысла.
Это положение великолепно показано в рефлекторной теории. Чтобы образовать условный рефлекс у животного, надо включить его в известную безусловную биологическую деятельность.
Может, существует только один момент, в который животное как будто выходит за пределы этого правила: его мощное развитие ориентировочно – исследовательской деятельности. Наблюдая высших обезьян, И. П. Павлов отметил их отличие от более низко стоящих животных, собак, кошек, тем более от кроликов, морских свинок. Если собаке или кошке нечего делать, она засыпает; если обезьяне нечего делать, она начинает исследовать, то есть ощупывать, нюхать или перебирать шерсть, перебирать листья и так далее. Все это время она занята тем, что Павлов назвал «бескорыстной ориентировочно – исследовательской деятельностью». Однако, и это перебирание предметов, рассматривание, обнюхивание может быть тоже трактовано как определенный безусловный ориентировочно – исследовательский рефлекс. Если это так, значит перебирание, обнюхивание, которое постоянно обнаруживает ничем не занятая обезьяна, то же являются биологической инстинктивной деятельностью.
Следовательно, первое отличие в поведении животного заключается в том, что всякое его поведение не выходит за пределы инстинктивной биологической деятельности и носит биологически мотивированной характер.
Второе отличие животного от человека несколько сложнее. Мы говорим, что животное может употреблять и даже выделять орудия. Но сейчас нам надо сделать известную поправку или уточнение этого факта, который с первого взгляда сближает поведение обезьяны с деятельностью человека. Животное, употребляющее и выделяющее орудия, всегда делает это в конкретной наглядно – действенной ситуации и никогда не закрепляет выделенное орудие, не сохраняет орудие впрок. На прошлой лекции, когда я излагал по выделыванию животным длинной палки из круглого диска, вы могли видеть, что животное никогда не оставляет эту палку впрок, для употребления в следующем опыте. Было многократно показано другими исследованиями, что даже применив известное орудие, животное начинает искать новое орудие каждый раз, когда дается новая задача.
Животное не обладает абстракцией от наглядной ситуации; внутри наглядной ситуации может выделить орудия, однако, вне этой наглядной ситуации животное никогда не может выделить орудия и зафиксировать постоянные средства, которые ему когда – нибудь понадобятся.
Оказалось далее, что обезьяна даже не пользуется одним орудием для того, чтобы с помощью этого орудия заготовлять другое и что ее орудийная деятельность не носит сложного, многократного опосредственного характера.
Когда в прошлый раз нам было дано сравнение поведения обезьяны и ребенка (эта лекция была прочитана Т. Хрустовым и давала обзор его опытов с применением «орудий обезьянами), вы видели, что если ребенок с большой легкостью начинает применять камень для того, чтобы расщепить диск, и ему достаточно одного показа, одного только намека для такой деятельности; наоборот – обезьяна не только не применяла орудий для того, чтобы сделать другие орудия, но даже бесконечный показ не подхватывается обезьяной. Она никогда не реагировала на подсказываемые человеком сложные, состоящие из цепи звеньев действия по употреблению одних орудий для того, чтобы изготовить другие.
Можно поэтому сказать, что животные не живут в мире постоянных вещей, имеющих постоянное значение. Вещь приобретает для него значение только в конкретной данной ситуации, в процессе деятельности. Один раз доска может быть для обезьяны подставкой, на которую она вскакивает, чтобы достать высоко подвешенный плод, в другой раз она может играть роль рычага, если ее нужно что – то достать; в третий раз – роль деревяшки, которую обезьяна сломает для того, чтобы изгрызть ее и так далее. Постоянного значения вещь для нее не имеет значения.
Поэтому можно сказать, что если человек живет в мире орудий, то обезьяна живет в мире средств для действия.
Таким образом, для обезьяны мир вещей – это вещи, которые могут приобрести определенное значение в процессе деятельности, но никогда вещи не представляют собой постоянно фиксированного для своей функции объекта. Это и составляет второе отличие животного, в частности обезьяны, от человека.
Третье отличие заключается в том, что животное может действовать только в пределах наглядно воспринимаемой ситуации. оно не может, в отличие от человека, абстрагироваться от наглядной ситуации и программировать свои действия соответственно отвлеченному принципу.
Для того, чтобы это положение стало ясным, я позволю себе сослаться на один опыт, который считаю, пожалуй, замечательнейшим из всех опытов, проведенных над животными, иллюстрирующих отличие поведение животного от психологической деятельности человека.
Опыт был проведен голландским психологом Бойтендайком. Перед животным (это могла быть кошка, собака или обезьяна) ставился ряд банок; под одной из этих баночек клалась приманка. Животное видело, что приманка положена под первую баночку. Совершенно естественно, что когда от первого опыта Бойтендайк перешел ко второму и снова предъявлял тот же самый закрытых баночек (естественно, что для чистоты опыта запах был исключен), животное направлялось к первой, подкрепленной в прежнем опыте баночке, открывало ее; однако под этой баночкой приманки нет. Опыт ставился по следующему принципу; каждый раз приманка перемещалась в следующую баночку; значит, животное должно было идти не к той банке, которая была подкреплена, а, наоборот, к банке, еще не подкрепленной, но, которая по всей видимости (если только усваивается принцип следующий»), будет подкреплено. Когда животное не находило ничего в первой банке, то оно шло ко второй и находило здесь приманку. Когда в следующий раз приманка переносилась в третью банку, животное направлялось опять к предыдущей банке и так далее.
Несмотря на большое количество проб Бойтендайку так и не удавалось получить у животного сразу реакцию на «следующую», еще не подкрепленную банку. Следовательно, животное усваивало известный опыт, всегда направляло свое поведение к ранее подкрепленному стимулу; ставить же принцип «следующий», то есть направить свое поведение не в сторону в прошлом подкрепленного места, а в сторону того места, которое должно было стать подкрепленным в будущем, животное не могло.
Иначе говоря, у животного не было того элементарного представления, которое характеризует поведение человека.
Интересно, что этот опыт Бойтендайка, как проверил, с очень большой легкостью становится доступным для ребенка даже двух с половиной лет, который обладает элементарной речью и который этот принцип может схватить достаточно легко.
Следовательно, этот опыт показывает, что поведение животного целиком детерминируется наглядно воспринимаемой ситуацией, опытом прошлого, но животное не может никогда выйти за пределы этой наглядной ситуации, абстрагироваться от нее и схватить отвлеченный принцип, иначе говоря, не может регулировать свое поведение предвидением, возникающим в результате усвоенного принципа действия.
То, что я вам сказал, как будто находится в противоречии с тем фактом экстраполяции рефлексов, о которых я говорил в прошлой лекции. Но это противоречие только кажущееся.
В опытах Л. В. Крушинского животное производит экстраполяцию наглядного движения предмета во времени; в опыте Бойтендайка оно должно схватить отвлеченный принцип, что не удается ему.
Эта невозможность абстрагироваться от наглядной ситуации и является очень четко выявленной особенностью поведения животного, и этот опыт я считаю таким важным потому, что он действительно кладет водораздел между поведением животного и поведением человека. Поведение животного в настоящем определяется его прошлым наглядным опытом. Поведение человека в настоящем может определяться образом его будущего, тем принципом, который будет экстраполировать его поведение в будущем. Поэтому только животное остается, как говорил Кёлер, рабом своего зрительного поля, рабом своего впечатления, а человек легко эмансипируется от него.
Невозможность отвлечься от непосредственно воспринимаемой ситуации или прежнего наглядного опыта является, таким образом, еще одним существенным признаком, отличающим поведение животное от психологической деятельности человека.
Обратимся к последнему признаку, чрезвычайно существенному для того, чтобы отличить поведение животного от психологической деятельности человека.
Если программирование поведения у животного всегда ограничено только двумя фактами, у человека к этим факторам прибавляется еще третий, не существующий у животных. Как мы уже говорили, поведение у животных определяется либо наследственно отложившимися видовыми программами, либо непосредственным личным опытом, иначе говоря, либо видовым, безусловным, либо условным рефлексом, получившимся в результате индивидуального опыта животного. Эти два факта и определяют поведение животного, они являются факторами его психологического развития. Еще не существует такой собаки, которая, получив известный опыт решения задачи, подошла к другой новой собаке и на ухо сказала ей: «А решить задачу нужно так». Нет такого животного, которое может передать свой опыт другому животному. В лучшем случае животное может передать свой опыт так, как передает его матери детенышам; с помощью непосредственного действия и подражания. Передача опыта или формулы опыта, принципа действия от одной особи к другой у животных невозможна.
В отличие от этого психологическая деятельность человека характеризуются тем, что человек наряду с этими двумя формами поведения (программированным наследственно и программированным личным опытом) обладает еще третьей формой поведения, которая становится все более и более доминирующей и у нас с вами начинает занимать доминирующее место: такой формой является передача общественного опыта от одного человека к другому человеку. Все обучение в школе, все усвоение знаний, все усвоение методов работы есть по существу передача индивиду опыта поколений, иначе говоря, передача общественного опыта от одного человека к другому. Если животное рождается на свет только с наследственной программой и обогащает ее личным условно – рефлекторным опытом, то человек, рождаясь с гораздо более бедными, чем у животного, инстинктивными программами, развивает свой психологический процесс не только с помощью своего индивидуального опыта других поколений через посредство воздействия других людей, усваивая опыт от других людей.
Усвоение общественного опыта не существует у животного, но существует у человека.
Вот в этом заключается последнее коренное отличие психологической деятельности животного от психологической деятельности человека.
Я осветил границы наиболее сложных форм поведения животных и показал, чем оно отличается от поведения человека. Займемся сейчас анализам основных особенностей психологической деятельности человека и разбираемся так же в том, в чем психологическая деятельность человека отличается от поведения и психологической деятельности животных. Подойдем к только что освещенному вопросу с другой стороны – со стороны анализа психологической деятельности человека.
Психологическая деятельность человека коренным образом отличается от психологической деятельности животного как по своей структуре, по своим функциональным особенностям и по своему генезу.
Я сейчас остановлюсь на трех отличительных особенностях психологической деятельности человека, характеризуя ее структуру, его функции и его генетические особенности.
Чем отличается психологическая деятельность человека от поведения животного по своему содержания и строению?
Прежде всего это отличие заключается в том, что вероятно в подавляющей своей части поведение человека не протекает внутри биологических мотивов и не определяется биологическими потребностями.
Если вы проанализируете психологическую деятельность человека, начиная с психологической деятельности ребенка и, тем более психологическую деятельность подростка или взрослого, то вы должны будете признать, что подавляющая масса актов поведения человека вовсе не определяется биологическими мотивами; силами, движущими поведение человека, вовсе не является голод, пищевой рефлекс, оборонительный рефлекс, половой рефлекс и так далее. деятельность человека определяется по своему содержанию другими мотивами, которые приобрели в науке очень невразумительное название «духовных» мотивов, вероятно, просто потому, что как – то надо было отличить их от мотивов животных, а подходящих понятий сразу не было найдено.
Примером такого поведения может служить поведение учения. Все вы учитесь вовсе не из каких – либо биологических мотивов. Мотивы ваши могут быть разными: одни учатся потому, что их интересует само содержание науки, и ими, следовательно, движет познавательный интерес; другие учатся потому, что хотят получить положение в жизни, и ими движут, следовательно известные мотивы престижа. Но какие бы ни были мотивы, они не являются биологическими.
Если проанализировать каждый отдельный поступок человека, то окажется, что и эти поступки в очень небольшом проценте являются актами, которыми движутся не биологическими мотивами.
Попытка свести поведение человека к биологическим мотивам, рассматривать это поведение как результат оборонительного или пищевого рефлекса есть величайшая вульгаризация психологии.
Значит, поведение человека по своему содержанию отличается от поведения животного тем, что оно программируется другими мотивами и потребностями, выходящими за пределы биологических и носящих характер так называемых духовных потребностей. В чем же состоят эти духовные потребности и как материалистически к ним подойти.
В отличие от животного, человек может ставить себе цели, и эти цели или задачи могут носить очень расчлененный характер, далеко выходя за пределы непосредственно биологически обусловленных актов. Даже если человек, вернувшись домой и будучи страшно голодный, захотел бы открыть шкаф, где хранится пища, а ключа от шкафа не оказалось, он идет к соседям, спрашивая, нет ли подходящего ключа, или к слесарю, чтобы заказать ключ, или находит орудие, с помощью которого можно взломать дверцу шкафа, – во всех тех случаях его действия не носят прямого биологического характера. Его задача заключается в том, чтобы вычленить ряд действий, которые последовательно осуществляет, чтобы постепенно подойти к выполнению цели; эти действия распадаются на ряд определенных, иерархически подчиненных друг другу операций, каждая из которых может не иметь прямого отношения к цели.
Учение, труд, трудовая деятельность – все заполнено у человека такими действиями, которые человек осуществляет сознательно или имеет ввиду то значение, которое эти действия получат и которые никакого биологического биологического смысла не имеют. Если бы я хотел изобразить такую деятельность в известной схеме, то я бы сказал, что деятельность животного, как бы сложна и извилиста не была, всегда ведет к достижению какой-то биологической цели, она превращается в известные действия, которые только в отношении с другими действиями, с целым циклом действий, а иногда и с действиями других людей может привести к известной цели и получить окончательный эффект. Таким образом, деятельность человека строится как расчлененная цепь взаимоподчиненных отдельных актов, которые лишь в конечной стадии могут привести к известному биологическому смыслу, но, которые сами по себе такого биологического смысла не имеют.
Отсюда, что очень существенно, – деятельность человека, определяемая целями, которые он сам ставит, задачами, которые он сам осознает, превращается в сознательную деятельность, регулируемую предвидением будущего, что у животного совершенно отсутствует. И вот этот расчет на будущее, то есть случаи, когда действие пускается в ход известным предвидением будущего, составляет подавляющее большинство конкретных актов нашего поведения.
Таким образов, первое отличие психологической деятельности человека заключается в том, что она далеко не всегда протекает в пределах биологических мотивов, и в том, что деятельность человека носит сложный иерархический построенный характер. Она расчленяется на ряд сознательных действий, каждое из которых несет в самом себе известные мотивы и получает свой смысл лишь в конечном итоге, иногда в индивидуальной работе, а иногда в работе других людей приводит к соответствующей цели.
Эта сложная иерархически построенная структура деятельности, в которой выделяются отдельные действия, получающие свой смысл только из того, что они соотносятся с конечной целью, а иногда приобретают смысл только в деятельности других людей, характеризует сознательную деятельность человека, отличая его от непосредственного биологического поведения животных.
Вторая отличительная черта психологической деятельности человека является его иное отношение к непосредственным влияниям воздействующей на него среды. Я уже сказал, что животное – раб непосредственно воспринимаемой ситуации, что поведение животного в настоящем определяется либо непосредственным восприятием этой ситуации, либо прошлым опытом, которое животное получило в этой ситуации. Поведение человека определяется другим: оно может определяться более глубоким знанием ситуации и предвосхищение того будущего, тех последствий, которые могут возникнуть в связи с его действиями.
Позвольте привести несколько примеров, которые дадут мне возможность прийти к соответствующему выводу. Ясный осенний день, в октябре. Вы отправляетесь на далекую прогулку. Небо ясное, солнышко греет. Вы решаете в 12 часов дня выйти из дома с тем, чтобы вернуться в 7 – 8 часов вечера. И вот вы отправляетесь в осенний солнечный день в далекую прогулку, ориентируясь вовсе не на то, что сияет солнце, что оно пригревает, и делаете как бы бессмысленную вещь: вы берете теплое пальто или непромокаемый плащ, хотя в непосредственном восприятии никаких мотивов для этого нет.
Чем вы руководствуетесь? Вовсе не своим восприятием; вы руководствуетесь более глубоким знанием закономерностей: вы знаете, что это осень, октябрь, что вернетесь поздно вечером, что октябрьским днем может быть дождь, а вечером, наверное, будет холодно. Вы определяете свое поведение не наглядными впечатлениями от ситуации, а планируете его согласно знанию законов природы, предвосхищая будущее. Как сейчас часто говорят, вы ведете себя, прогнозируя большую вероятность появления холода или дождя, и реагируете на отсутствующий дождь или холод, исходя из знания тех закономерностей, которые таит в себе осенний, хотя и солнечный день.
Второй пример, который должен показать, что это прогнозирование будущего может иногда даже вступать в конфликт с прямыми биологическими потребностями.
Представьте себе человека, который страшно хочет пить. Он подходит к водоему и готов зачерпнуть воды и выпить ее; но ему говорят: «Знаете, в этом водоеме обнаружен холерный эмбрион, имейте это в виду!». Человек может скорее умереть от жажды, чем начать пить из этого зараженного водоема.
Это значит, что прогнозирование последствий известного действия может быть даже сильнее, чем непосредственная биологическая потребность.
Какое бы действие человека вы не взяли, нормальный человек, если он только не находится в состоянии аффекта, если его поведение не снижено, ведет себя совершенно отлично от того, как ведет себя животное: его деятельность не подчиняется непосредственному восприятию, а анализу этой ситуации, а анализу этой ситуации, в результате которого сигналы от этой ситуации доходят до него в перешифрованном виде. Эта перешифровка получаемой информации и дает человеку возможность подчинять свое поведение не непосредственному впечатлению, а знанию закона или правила, отражению внутренней необходимости, которое лежит за непосредственным восприятием.
Философски рассуждая, здесь впервые возникает поведение, свободное, в отличии от поведения рабского. Животное – раб воспринимаемой ситуации, человек – хозяин воспринимаемой ситуации. Животное подчиняется непосредственному впечатлению, человек подчиняется перешифрованным сигналам, то есть сигналам, которые исходят не от непосредственного восприятия от той необходимости, которые человек расшифровал, познал и которым подчиняется.
Если человек в моем первом примере не возьмет с собой плащ или пальто и отправится на прогулку поздней осенью налегке, его поведение будет неразумным, несознательным, несвободным потому, что он подчиняется непосредственному впечатлению. Если он сделает обратное, его поведение будет сознательным, разумным и свободным, то есть будет абстрагироваться от непосредственного впечатления.
Если в моем последнем примере человек, желающий пить, выпьет воду из зараженного водоема, его поведение будет несознательное и несвободное, реактивное. А если человек воздержится от этого и, несмотря, на то, что он испытывает величайшую потребность выпить воды из этого водоема, все – таки не выпьет, его поведение будет сознательным, разумным и свободным, ибо оно исходит из познанной необходимости.
Таким образом, поведение человека, в отличие от поведения животного, может подчиняться не непосредственному впечатлению, а анализу закономерностей, которые могут скрываться за непосредственной ситуацией. Поэтому поведение человека, который поступает не реактивно, соответственно впечатлению или потребности, а соответственно познанной необходимости, может оцениваться как поведение свободное.
Таким образом поведение человека по структуре сложное иерархическое, по функции – произвольное или свободное, основанное на перешифровке непосредственных впечатлений. Этим оно резко отличается от поведения животных. Если, как я говорил 2, 5 – ий летний ребенок легко справляется с опытом Бойтендейка, то это потому, что он перешифровывает ситуацию и реагирует не на впечатление от подкрепленного прошлым объекта, а на абстрактной принцип: «Ага, приманка в следующем». Вот почему то, что является границей для человека.
Третья особенность человека заключается в том, что, как я уже рассказывал, поведение человека не определяется только двумя факторами – наследственными программами и личным опытом, – поведение человека определяется той информацией, которую он получает от других людей. Усваивая общечеловеческий опыт, человек получает возможность использовать знания и средства уже в готовом виде. Этот факт имеет огромное значение.
То, что человек живет в митре готовых вещей, в которых материализовался прежний опыт, что он усваивает в школе таблицу умножения и законы рычагов – все это говорит о том, что человек не вынужден заново изобретать каждый раз те вещи, которые были изобретены и применялись в процессе истории.
Следовательно, человек обладает третьим фактором развития – программированием своего поведения с помощью усвоения общечеловеческого опыта, и это резко отличает его от животного, которое ничего не получает из опыта других животных в готовом виде. Тот факт, что человеку не нужно самому формировать опыт, а этот опыт поступает к нему в готовом виде, в известной общественной информации, представляет следовательно, третью существенную особенность психологического процесса человека. Эта особенность связана уже не со структурой или функцией, а с происхождением, генезисом его сложных форм психологической деятельности.
Я кратко остановился на трех основных особенностях психологической деятельности человека, расчлененной по своей структуре, свободной от непосредственного впечатления по своей функции и основанной на своей информации, которая в готовом виде дается ему, передавая общественный опыт поколений по своему генезису.
Когда я сделал эти вещи, я по существу определил основные особенности сознательного поведения человека. Под сознательным поведением мы имеем в виду такое поведение, которое подчиняется известному анализу ситуации и ориентируется на предвидение будущего, соотнося каждый факт поведения с другими возможными актами, приводя настоящее в связь с будущим.
Возникает, однако, важный вопрос, который заключается в следующем: как же можно трактовать происхождение вот таких сложных сознательных форм психологической деятельности, которые явно отличают психологическую деятельность животного от психологической деятельности человека.
В чем же заключается сущность этой сознательной психологической деятельности, как трактовать материалистически, научно трактовать сознание человека? Среди всех загадок природы, которые были поставлены одним из крупных естествоиспытателей прошлого века, фигурирует и эта загадка, считавшаяся им неразрешимой: он считает, что есть несколько мировых загадок, ответ на которые мы никогда не откроем, и одной из таких загадок является естественная история сознания.
Как же подойти к ответу на этот вопрос? Как объяснить тот факт, что из интеллектуального, импульсивного поведения животных возникает сознательная деятельность человека?
На этот вопрос разные теории давали три разных ответа.
Первая теория открытого идеализма давала такой ответ: сознание присущи человеку как имманентное, характеризующее его внутреннее свойство. Этим свойством человек отличается от животного коренным образом, этим он противопоставляется животному. В еще более резкой форме это было сформулировано Декартом: животные есть механизмы или машины, они работают по законам природы, человек есть существо духовное, он действует по законам разума. Сознательность объясняется тем, что человек обладает кусочком бессмертной души, которую вложил в него бог, положив этим отличие человека от животного.
Эта точка зрения господствовала в течение многих веков и в скрытом виде еще продолжает господствовать во многих зарубежных психологических теориях. Она может быть как угодно завуалирована, но психолог – идеалист думает так: животное работает по биологическим законам, а человек действует по субъективным законам, заложенным в его душевной жизни, – он существо сознательное. Такая точка зрения была настоль сильна, что Сеченов меньше чем 100 лет назад говорил так как о злейших врагах науки об «обособителях» в психологии, то есть о тех, кто обособляет психологию от всякой физиологии и биологии, желая найти в психологии духовную сущность и отрицая эту духовную сущность у животных.
Вы ясно видите, что эта идеалистическая имеет по существу агностический характер, потому что она представляет отказ от всякого объяснения сознательной жизни человека, отличающейся от сознательной жизни животных.
Вторая теория – происхождение сознания – это диалогическая теория.
Если первая теория хочет вывести сознание из глубины духа, то вторая теория пытается вывести сознание непосредственно из естественной эволюции животных. Эта точка зрения позитивистов – что это точка зрения механических материалистов. Исследователи примыкающие к этой позиции, считают, что уже у животного можно видеть известные формы инстинктивной жизни, которые приближаются к сознательным, и эти сторонники биологической концепции стараются, наоборот, стереть грань между человеком и животным.
Так, например, Дарвин считал (и для его времени это было правильной и актуальной задачей), что он должен найти в поведении животных такие формы, которые сближают их с человеком; поэтому он очень очеловечивал животных – очень антропоморфизировал их, считая, что и животные обладают теми же формами интеллектуального поведения, что и человек.
Исследователи, пытающиеся вывести из биологической эволюции, не находят иного выхода, как попытки трактовать сознание как сложнейшую форму инстинкта, который может быть свойственен и животным.
Естественно, что эти попытки приводят в тупик потому, что факты, которые мы знаем здесь, заставляют не смазывать разницу между человеком и животным, а в полный голос говорить о различии сознательной деятельности человека и инстинктивного поведения животных.
В чем же состоит правильное решение вопроса о сознательном поведении человека?
Решение вопроса о природе сознательного заключается в том, что выводит это сознательное поведения не из глубины духа, не из естественного развития животных, а из новых общественных форм поведения, которые отличают жизнь человека от жизни животных.
Человек отличается от животного тем, что он живет в условиях общественного существования. Его формы жизни – это общественные формы жизни, и, именно из этих общественных форм жизни могут быть выведены те особенности поведения, которые мы называем духовными.
Этот путь является важнейшим для советской психологии: чтобы объяснить внутреннее психологическое, мы должны рассмотреть внешние формы жизни, общественную жизнь человека и найти объективные внешние причины, которые позволяют перейти от животных форм существования к общественным формам психологической жизни.
Позвольте мне совсем кратко на этом остановиться.
С переходом от животного существования к общественной истории возникают два важнейших события. Во – первых, это переход к общественному труду и употреблению орудий, и, во – вторых, переход к языку, к общению с помощью слова.
Эти два факта и лежат в основе появления новых форм поведения. Из них, из общественного труда и из языка возникают те особенности сознательной формы психологической деятельности, которые отличают человека от животного.
Позвольте сегодня остановится на первом из них – употребление орудий и общественного труда.
Мы видели, что животные могут употреблять известные внешние средства и даже выделять эти средства в тот момент, когда они заняты соответствующей деятельностью. Но у них никогда не возникает устойчивых форм трудовой деятельности и постоянного изготовления орудия впрок. Иначе у человека: с возникновением прямохождения у человека освобождается рука. И с дальнейшим развитием мозга и, прежде всего, сложных форм мозговой коры, позволяющих объединять совместную работу отдельных анализаторов. Благодаря этому у него возникают условия для мощного развития ориентировочной деятельности, которые и приводят к возникновению орудий. В курсе антропологии вы уже встречались с анализом наиболее простых форм орудий труда. Это раньше были осколки или готовые камни, которые никак не материализовали операции, которые с ним производятся. Лишь в позднейшие эпохи орудия приняли уже более сложный характер: в этих более поздних орудиях уже существует сторона, за которую орудия держат и которая приспосабливается к руке; режущая сторона, предназначенная для выполнения операций – рубить, снимать кожу, скрести, колоть. Когда появляются такие орудия, возникает, следовательно, материализованная операция: операция «резать», «колать», «скрести» становится постоянной материальной особенностью предмета. В таких материальных особенностях орудия мы уже находим закрепившиеся, постоянные операции. Именно в связи с этим возникает первое огромное отличие среды человека от поведения животного. Животное существует в мире случайных вещей, которые приобретают свое значение лишь в данной операции; человек начинает жить в мире постоянных предметов и орудий, в которых материализованы, фиксированы его операции. Тем самым, у человека возникает передавать операцию от одного поколения к другому, и этот факт материализации операции в орудии становится фактов первостепенной значения.
От сюда вытекает второй важный факт: эти орудия начинают изготавливать впрок, и тем самым действие изготовления орудий начинает отрываться от непосредственной деятельности по добыче пищи. Изготовление орудий, отламывание и обтесывание камня, с первого взгляда может показаться бессмысленной деятельностью, потому что оттачивать камень не значит получать пищу; однако в дальнейшем этот камень, который оттачивается, сможет быть использован для получения пищи, и в соотношении с этой конечной целью данное действие становится осмысленным. Таким образом, возникает первое сознательное действие, иначе говоря, первое отделение операции изготовления орудий от конечной деятельности – добычи пиши с помощью этого орудия.
С изготовлением и употреблением орудий возникают материализованнные операции, но возникает первая абстракция: человек абстрагируется, отвлекается от непосредственной пищи – добывающей деятельности и все сосредотачивается на изготовлении орудий, которые получают свой смысл только для того, что оно дальше будет применено для добывания пищи. Изготовление орудий тем самым является первым решающим фактором для того, чтобы понять возникновение действия, оторванного от непосредственной цели.
Второе, о чем следует кратко сказать, представляет собой факт не меньшего значения.
Труд человека никогда не осуществляется им в одиночку, он всегда осуществляется в обществе. Человеческое общество тем и отличается от животной стаи, что люди трудятся совместно и в процессе этого общественного труда разделяются свои функции. вначале разделение функций носит еще биологический характер: женщина поддерживает огонь, мужчина идет на охоту. Но разделение функций разбивает цельную всю деятельность на определенные звенья, каждое из которых может выполняться отдельным человеком; поддерживать огонь – занятие женщины – деятельность, которая приобретает свой смысл только для того, что дальше на этом огне можно будет изжарить пищу, которую добывал мужчина.
В этом разделении операций – величайшие шаги к расчленению деятельности на отдельные подчиненные звенья, получающие свой смысл из соотношений с конечной целью, в формировании абстрактной сознательной деятельности.
Разделение общественного труда иногда принимает такие формы, которые иногда заставляют человека совершать действия, обратные биологическим мотивам.
А. Н. Леонтьев приводит один замечательный пример.
Общественно – организованная охота в первобытном обществе иногда так построена, что одна группа людей отгоняет дичь для того, чтобы дичь попала в засаду, в то время как другая группа людей поджидает и убивает дичь, отгоняемую первой группой.
Какой биологический смысл имеет работа первой группы людей – отгонять дичь? Никакого. Наоборот, это нечто противоположное биологическому смыслу. Но в общественно – организованном труде это бессмысленное действие приобретает свой смысл, если один человек отгоняет дичь, а другой получает возможность поймать ее. В общественном разделении труда отдельных людей начинают возникать такие действия, которые лишены прямого биологического смысла и которые приобретают свой смысл лишь из действий других людей.
В этой общественной организации труда и заключается корень возникновения сознательной деятельности человека.
То, что я вам сказал, можно сформулировать следующим образом. У человека, для которого характерен общественный труд, функция часто разделяется между двумя людьми: один человек начинает выполнять данную задачу, чтобы другой мог ее закончить.
Функция, которая у животных всегда осуществляется одним индивидом, разделяется здесь между двумя индивидами (двумя группами), и это есть специфика общественного труда, из которой возникают грандиозные психологические следствия.
Если женщина поддерживает огонь, а мужчина охотится; если один человек делает орудие, а другой применяет его; если один человек отгоняет дичь, а другой человек ловит дичь, – то этим самым создается новая общественная организация деятельности, а эта общественная организация деятельности и приводит к тому, что единая биологическая деятельность расчленяется, выделяются отдельные части этой деятельности или отдельные сознательные действия, которые приобретают смысл только в работе другого человека.
Эти сознательные действия и можно понять только из общественной организации деятельности человека.
В этом подходе и состоит марксисткое решение вопроса о происхождении сознания, которое я наметил вам лишь в самых кратких чертах.
Сознание происходит не из глубин духа и не из недр биологии – сознание происходит из общественной организации поведения человека и его психологической деятельности. Поэтому психология есть не только биологическая, но и социальная наука, и это ни на минуту нельзя забывать, изучая психологию.
Л.А. Фирсов обезьяны, берущие палки
Печатается по изданию: Л. А. Фирсов. И. П. Павлов и экспериментальная приматология.М.: 1982. Изд-во: Ленинградское отделение «Наука».
В современной физиологии поведения, этологии и зоопсихологии весьма активно разрабатывается проблема способности животных к использованию различных предметов. Большое внимание этому уделяется в дискуссиях отечественных и зарубежных исследователей (Williams, 1978), а также советских антропологов, зоопсихологов и философов. Единство мнений пока не достигнуто. Более того, мысль, что животное может оперировать орудием, все-таки считается неприемлемой, хотя еще К. Маркс (1961) указывал на необходимость различения примитивных животнообразных форм труда. Использование природных объектов в целях актуализации самых разных форм поведения животных общеизвестно. Наиболее часто оно проявляется в виде сложных биокомплексов поведения (Промптов, 1947), имеющих преимущественно безусловнорефлекторную природу: закладывание крабом песчинок в свой отолитовый аппарат после очередной линьки; разбивание стервятником страусового яйца камнем или съедобных предметов с твердой оболочкой каланом и капуцином1; раскрашивание шалашником своего сооружения размочаленным кусочком коры; овладевание шимпанзе удаленной от него приманкой с помощью отломанной ветки. Очевидно, что во всех случаях животное удовлетворяет какие-то свои потребности. Однако вряд ли можно считать сколько-нибудь полезным такое обобщение под одним термином «предметная деятельность». Ближайшее рассмотрение показывает, что в одном случае мы имеем дело с восполнением регулярно проявляемой утраты (песчинки), в другом — с использованием камня в стереотипных условиях пищевого поведения, вероятно, формируемого в порядке «социального наследования» (Дубинин, 1977), в третьем — с врожденным компонентом токового поведения самца; и, наконец, в четвертом — с использованием предмета для приближения к себе другого предмета (пища, игрушка) или для других контактов. К сожалению, подавляющее число авторов не различает этих уровней организации предметной деятельности в сравнительном аспекте. Так, некоторые исследователи удивительным образом путают предметную и орудийную деятельности, а под последней зачастую описывают определенные двигательные навыки, выработанные у различных животных в порядке дрессировки. По мнению Н. П. Дубинина (1977), «использование орудий в пределах работы первой сигнальной системы свойственно многим видам животных». Нам, однако, кажется, что за пределами антропоидов и человека искать орудийную деятельность у других животных нет основания. Или в таком случае следует признать, что орудийная деятельность имеется у всех животных, как об этом, например, пишет К. Э. Фабри (1980) и некоторые западные исследователи (Beck, 1972, и др.). Вместе с тем изучение способностей животных к использованию предметов окружающей среды показывает, что только антропоиды применяют обширный набор предметов в самых различных ситуациях. При этом обнаружено, что предмет может быть даже частично обработан, а разрозненные предметы — соединены. На наш взгляд, в перечисленных проявлениях этого важного свойства в поведении многих животных трудно не видеть существенного видового различия. Что же касается поведения шимпанзе и других антропоидов, то использование объектов окружающей среды оказывается для них не только регулярным и полиморфным, но отличающимся также подчеркнутой целенаправленностью выбора предмета из множества и активным приспособлением его для конкретных условий задачи. Таким образом, использование животными непищевых предметов — несомненный показатель степени их эволюционного развития. Как установлено за последнее десятилетие, орудийная деятельность семейства гоминид является более важным систематическим (таксономическим) признаком, чем многие другие (Нестурх, 1970, Якимов, 1972; Урысон, 1972; Бунак, 1980, и др.). К сожалению, в работах наших и зарубежных авторов под видом орудийной деятельности животных часто описывается поведение низших и высших обезьян и даже других позвоночных, сформированное у них в направленном эксперименте. Орудийной деятельностью считаются также такие генетически запрограммированные формы поведения, как устройство плотин бобрами, употребление галапагосским вьюрком колючек кактуса для выковыривания личинок и т. д. Нам представляется, что подобное смешение форм использования предметов, основанное на чистейшей аналогии, нельзя признать корректным. При сравнительно-физиологическом и онтогенетическом изучении спонтанной, не направленной экспериментом, предметной деятельности у низших (капуцины, макаки, павианы, мартышки гелады, саймири) и высших (шимпанзе, горилла, орангутан) обезьян обнаруживаются существенные отличия. В экспериментальных ситуациях, созданных нами в лабораторных условиях, а также в природных условиях лесистых островов и заказника в урочище р. Гумисты (Абхазская АССР) для шимпанзе, павианов гамадрилов и макак резусов, ни одна низшая обезьяна из числа указанных видов не использовала палку или иной предмет для целенаправленного преодоления дистанции в горизонтальном или вертикальном направлениях, что отвечало бы пониманию ими довольно примитивных причинно-следственных отношений. Опыты с палками для доставания пищевой приманки, находящейся вне клетки, были проведены Н. Л. Парфеновой и М. А. Ширинкиной по одной схеме на капуцинах (6), макаках резусах (22) макаках лапундерах (4), павианах гамадрилах (17), павианах анубисах (5), геладах (3), зеленых мартышках (4) в нашей лаборатории (Колтуши), а также на базе Пермского и Ленинградского зоопарков и в Сухуми на клеточных и свободноживущих обезьянах. Общее заключение, которое можно сделать на основании этих опытов, таково: без специальной дрессировки ни один вид исследованных низших обезьян не способен использовать предмет для целенаправленного приближения приманки. В специальной серии опытов, осуществленной по совету проф. А. М. Дубинского, капуцины не изменили своего поведения (игра, ориентировочно-исследовательская деятельность, разрушение) с палками даже после многих показов им навыков овладевания пищей при помощи палки с крючком, которым был обучен капуцин-демонстратор. Как известно, в опытах В. Я. Кряжева (1965) павианы гамадрилы действовали с помощью палок тоже после достаточно продолжительного обучения, но по подражанию они усваивали навык использования палки быстрее. Подобные опыты с шимпанзе разного возраста (Гамма, Бой — около 1 года, Вега — с 3 мес, Каролина — около 3,5 лет, Тролль — около 2,5 лет) показали, что в возрасте 2 — 2,5 лет в условиях лаборатории шимпанзе систематически проявляет способность использования предметов в самых различных ситуациях. По-видимому, к указанному сроку у шимпанзе созревают нервные механизмы, позволяющие вести себя в определенных ситуациях качественно другим образом по сравнению не только с другими животными, но даже с низшими обезьянами. Это качество — целенаправленное использование предметов, а также выбор наиболее подходящего предмета из множества. На детенышах шимпанзе, родившихся в Колтушах и воспитывавшихся с первого дня в изоляции от взрослых обезьян (Лель, Чингис) или с матерью (Бой-2), использование различных предметов в связи с определенной целью было отмечено даже несколько раньше двухлетнего возраста. Любопытно, что между первой и последующей попытками детеныша Боя-2 достать палочкой привлекший его внимание предмет прошло три месяца: он как бы забыл свое первое действие. Вторая попытка в отчетливой форме проявилась во время экспедиции 1978 г. на Обезьяньем острове. Внимание Боя-2 приковала медицинская шапочка, которую он несколько раз с успехом стянул с головы экспериментатора и бурно играл с нею. После того как в очередной раз шапочка была у него изъята, ее бросили в воду недалеко от берега. Бой-2 некоторое время смотрел на шапочку, потом протянул к ней руку и даже сделал шаг в воду, но тут же отступил назад. Только после всего этого детеныш поднял тонкую сухую хворостинку и дотянулся до шапочки. Действия его были во время доставания вполне направленными, и за две минуты шапочка была добыта и опять использована в игре (наблюдение от 17 июля 1978 г.). Примечательно, что шимпанзе в своем ареале, как об этом пишет Дж. Лавик-Гудолл (Goodall, 1963; Lawick-Goodall, 1974), применяют природные объекты для целенаправленных действий тоже не сразу и даже не тогда, когда они уже способны активно передвигаться, играть с предметами, лазить по деревьям. Целенаправленное использование палочек и других «удлиняющих» руку предметов у диких шимпанзе проявляется в возрасте около 3 лет. Вполне возможно, что возраст несколько завышен. Одним из важных факторов в обнаружении этой формы поведения, по мнению Лавик-Гудолл, является подражание детенышей взрослым особям. Вероятно, это так, коль скоро детеныш шимпанзе неотступно следует за своей матерью и другими взрослыми особями, внимательно следит за их поведением и в особенности за новыми элементами поведения, но, как обнаружилось на наших детенышах-изолятах, целенаправленность действий с предметом возникает и без подражательного толчка. Многообразие и тонкость операций шимпанзе с предметами не подлежит сомнению и, вероятно, эта деятельность у них развита намного выше, чем у гориллы и орангутана, отличающихся некоторой флегматичностью, и даже у молодых особей. Достаточно подростку шимпанзе дать новую незамысловатую игрушку, как он моментально включает ее в игру. Такая черта в поведении антропоидов неоднократно была отмечена И. П. Павловым. Рассказывая, например, о своем впечатлении от знакомства с прибывшими в Кол-туши Розой и Рафаэлем, он замечает: «...первый факт, который чрезвычайно поражает, первый вывод, который я делаю из наблюдения за этими обезьянами — это чрезвычайно высокое развитие у них исследовательского рефлекса» (Павловские среды, 1949, т. 2, с. 68; разрядка моя, — Л. Ф.). И далее: «Совершенно независимо от еды, обезьяны охвачены постоянным стремлением исследования» (с. 69; разрядка моя, — Л. Ф.). Павлову же принадлежит тонкая индивидуальная характеристика шимпанзе по их отношению к предметам: «„Роза" умнее „Рафаэля". Она представляет сравнительно высокий тип „интеллигентности", в то время как „Рафаэль'' — просто утробистый господин. Единственно, что его привлекает, — это еда. У „Розы", наоборот, еда на втором плане. У нее часто превалирует сильное желание поиграть или даже „помастерить" — открыть какую-нибудь коробку и т. д. Когда она занята, а вы предлагаете ей еду, то она ее отталкивает: цель занятий ее другая» (с. 584). Эти слова об «утробистом господине» Рафаэле нельзя понимать буквально, речь идет о преимущественном проявлении разных форм поведения в одинаковых экспериментальных условиях. Неукротимая тяга обезьян к предметам, особенно новым, к игре с ними, к использованию предметов для различных нужд (ковыряние в зубах, почесывание и очищение кожи, прикасание к различным животным и т. д.) отмечалась многими отечественными и зарубежными исследователями (Ладыгина-Котс, 1935; Yerkes, 1943, Вацуро, 1948; Рогинский, 1948; Воронин, 1952, 1957; Alcock, 1972; Beck, 1972, Фирсов, 1977, и др.). Материалы нашего исследования о взаимоотношении пищевой и игровой условнорефлекторной деятельности у детеныша шимпанзе мы смогли подтвердить в групповом эксперименте с изменением уровня мотивации у двух шимпанзе (Счастный, Фирсов, 1965). В задачах с отсроченным выбором из множества при игровом подкреплении, как правило, обнаруживался более высокий и стабильный уровень решений (Фирсов, 1972). Нам представляется, что способность антропоидов (шимпанзе) использовать предметы в различных ситуациях следует рассматривать в комплексе таких биологических качеств, как высокоразвитая ориентировочно-исследовательская деятельность, подражание, условнорефлекторная (долгосрочная) и образная память, обобщение и др. Разумеется, весь этот комплекс функций, важный для поведения, не может не стоять в теснейшей связи со структурно-функциональными особенностями головного мозга обезьян, в особенности антропоидов, анатомическим строением их кисти и органов чувств, в первую очередь зрительного анализатора. Что касается прогресса в строении головного мозга, то у шимпанзе и гориллы обнаружено усиленное развитие нижнетеменной области, которая оказывается увеличенной в несколько раз даже по сравнению с низшими обезьянами (Шевченко, 1971). Во время наших семи экспедиций в Псковскую и Ленинградскую области (1972 — 1980 гг.) мы имели возможность наблюдать, заснять на кинопленку, а также использовать для экспериментов способность шимпанзе пользоваться природными объектами (палки, стебли травы, камни, раковины и пр.). Одни предметы использовались ими как бы для увеличения естественной длины конечности, другие — для самообслуживания (почесывание, очищение, доставание до того места, куда не хватало длины конечностей), третьи — для усиления некоторых действий. Как мы смогли неоднократно убедиться в лаборатории и еще лучше в природе, физические свойства рычага хорошо известны уже подростку шимпанзе. При всем этом в случае необходимости предметы могут быть достаточно точно брошены в цель. Чтобы иметь представление о сложности и точности действий шимпанзе, следует рассказать об инциденте с двумя подростками Ладой и Невой — летом 1956 г. обезьяны смогли выбраться из надежно закрытого вольера. Лада и Нева находились в обширном вольере, где, кроме полок, был небольшой стол, надежно прикрепленный в углу вольера. Довольно основательная крышка стола (30 мм толщины) имела небольшой напуск вдоль длины стола. Эта деталь существенна для понимания происшедшего. Когда закончился рабочий день, лаборантка закрыла на два замка дверь вольера, занавесила большие окна, одно из которых находилось в 1,5 м от решетки вольера и покинула здание лаборатории. Как выяснилось впоследствии, она допустила ошибку: ключи оставила на столе, а не вынесла в коридор. Ближайший край стола, на котором лежала связка, располагался в 2,7 м от решетки вольера. Следовательно, связка ключей, до которых оказалось чуть меньше 3 м, безраздельно завладела вниманием обезьян. Первое, что они сделали (вероятней всего Лада, так как во время воспроизведения именно она дважды повторила выход из вольера), отбили нависший край столешницы, отчего получилась палка длиной около 1 м. Кто проделал эту главную операцию, которая не была нами воссоздана при последующем воспроизведении выхода, неизвестно. Обследование всей длины отщепа показало, что скрытых дефектов в толще доски не было, однако она имела множество следов от довольно основательных, уже постоянных, клыков обезьян. При помощи получившейся палки обезьяны дотянулись до ближайшей занавеси и, сдернув с подвесок, втащили ее в вольер. Далее, набросив штору, как лассо, на стол, обезьяны смогли придвинуть к себе связку ключей. Заключительный этап уже никаких усилий не требовал: с открыванием замка каждая из обезьян справлялась за считанные секунды. Поведение обезьян, после того как они оказались вне клетки, лишний раз говорит об их неукротимом «стремлении исследовать» (И. П. Павлов) все, что оказывается для них новым. Они были обнаружены поутру в здании антропоидника возле лабораторного стола, уставленного различной посудой и растворами. Они играли, ничего не разбив и, вероятно, ничего не приняв внутрь. При нашем появлении они не обратили на нас ни малейшего внимания и, словно пробудились ото сна, когда мы стали их звать к себе. Нам сразу же могут возразить, что отлом обезьянами куска столешницы именно в те часы, когда их внимание было мобилизовано забытой связкой ключей, простое совпадение. Прямых доказательств обратного нет, разве только то, что этот стол годами стоял на своем месте, по нему ежедневно прыгали эти же обезьяны и никаких попыток к разрушению столешницы никогда не было. Важна сама по себе поэтапность событий, которые Лада затем дважды продемонстрировала, доставая ключи. В проверочном наблюдении не было только момента отторжения куска столешницы. Воссоздав всю обстановку выхода, мы выделили в ней четыре главных эпизода: 1) отлом нависшего края столешницы; 2) доставание палкой занавеси и втягивание ее внутрь вольера; 3) набрасывание занавеси на связку ключей и подтягивание их к вольеру; 4) открывание замков и выход обезьян на свободу. Самым сложным действием в техническом смысле оказался третий эпизод. Прицельно набросить громоздкую занавесь на связку ключей — дело не простое. Лада бросала занавесь одним концом, удерживая в руках другой, не рискуя при неудачном броске не достать и ключей и занавеси лишиться. Вот, наконец, при очередном броске занавесь накрыла ключи и обезьяна со всеми предосторожностями, очень медленно тянет ее к себе. По звуку сдвигаемых ключей, вероятно, она определяет, что дело близится к завершению, но вдруг занавесь соскальзывает, а ключи остаются на столе, слегка сдвинутые в нужном направлении. Дальше следуют все новые и новые броски. Вот ключи упали со стола на пол, вот они уже покрыли половину расстояния между столом и решеткой, наконец, до них уже можно дотянуться рукой. Занавесь тут же отбрасывается, и ключи используются по прямому назначению. За всем поведением в проверочных испытаниях мы следили скрыто и появлялись в помещении, когда обезьяна начинала открывать замок. Если отбросить время на отбитие куска столешницы (это определить не удалось бы даже в случае повтора), то на все остальное требовалось около получаса. Этот случай в нашей лабораторной практике, детально разобранный и экспериментально воспроизведенный, во-первых, указывает на сложность действий шимпанзе, каждое из которых несет «орудийную» нагрузку, т. е. включает изготовление предмета, а также дальнейшее его использование (отбитие куска столешницы, доставание палкой, подтягивание с помощью занавеси, открывание ключами). Во-вторых, он не исключает того, что разрушение крышки стола имело в своем основании целенаправленный характер, тем более что откусывание лучин от доски для дальнейшего употребления их в конкретной экспериментальной задаче — явление для шимпанзе заурядное (Ладыгина-Котс, 1935; Хрустов, 1976). Это нами не только описано, но и заснято на пленку3. В связи с приведенным случаем уместно напомнить высказывание И. П. Павлова относительно предметной деятельности Розы и Рафаэля, которые «едой даже меньше занимаются, а больше например, механическим воздействием на окружающую природу» (Павловские клинические среды, 1954, т. 1, с. 260). Павлов писал, что шимпанзе свойственно не только проявление напряженного внимания к внешним предметам, но и обнаружение способа воздействия на них также с помощью предмета: «В помещении, где находились обезьяны, обрезали трубу и забили ее пробкой, так две обезьяны потратили три часа на то, чтобы эту пробку вытащить, и в конце концов достали гвоздь и гвоздем выковыряли эту пробку». А далее комментарий: «Человек несомненно двигался вперед не только благодаря слову, но и благодаря своим рукам» (там же). Приведем далеко неполный список использования предметов нашими шимпанзе в природных условиях Псковской области: обследование черепахи соломинками (Тарас, Бой, Гамма, 13 августа 1973 г.), перекатывание с помощью палочки личинки дубового шелкопряда (Гамма, 8 июля 1974 г.), обследование ежа с помощью небольших палочек (Тарас, Бой, Гамма, 14 июля 1974 г.), бросание палок и водорослей в аквалангиста (Бой, Гамма, 14 августа 1973 г.), выламывание длинной хворостины и заклинивание ею дверцы проблемного ящика (Тарас, 17 августа 1974 г.), бросание камней в чучело крокодила (Тарас, 24 июля 1978 г.) и многое другое. Следовательно, шимпанзе могут использовать как уже имеющийся готовый предмет, так и проделать несколько операций, чтобы затем воспользоваться подготовленными предметами для своих целей. Мы наблюдали это несколько раз у Сильвы и Читы, когда они делали из тонких веток черемухи, ивы и лещины небольшие окоренные палочки для ловли муравьев. В одном из опытов Тарас не мог дотянуться до столика, который был сооружен в воде на некотором расстоянии от берега. На столе лежала вкусная пищевая приманка. Походив вдоль берега, Тарас повернулся и исчез в ближайших кустах. Вскоре он вернулся с длинным байдарочным веслом. Это была несомненная репродукция когда-то увиденного предмета, с которым Тарас, по-видимому, мог играть, но в нужный момент экспериментальной ситуации весло было принесено, а с его помощью шимпанзе несколько раз пытался достать до столика, но длины весла не хватило; тогда Тарас положил его на воду и попытался пройти по нему, как по мосту. От первого же прикосновения ногой весло потонуло, Тарас вынул его из воды и стал играть с ним. Второй впечатляющий случай, когда этот же шимпанзе во время опыта использовал предмет нужной длины. Тарасу была дана задача добыть баночку компота из аппарата с самозахлопывающейся дверцой. Чтобы открыть дверцу, следовало потянуть за ручку, достаточно удаленную от аппарата. Длины двух рук обезьяны не хватало, чтобы потягиванием рукоятки одной рукой другой открыть дверцу и дотянуться до баночки с приманкой. Перепробовав всевозможные приемы, Тарас ушел от аппарата в сторону ближайших кустов. По дороге он поднял небольшую хворостинку и тут же ее бросил. Через пару секунд он потянулся к сухим веткам ольхового куста, отломил короткую тонкую веточку и снова бросил. После этого Тарас выломал довольно длинную и прочную хворостинку и с нею вернулся к аппарату. Не проявляя игры или других признаков отвлечения, он с силой потянул за рукоять тяги, открыл дверцу и вскоре довольно точно заклинил ее концом хворостины. Как только шимпанзе оценил все происшедшее, он тут же оставил рукоять и хворостину, стремительно отправился к аппарату, легко отжал дверцу и добыл баночку с компотом (22 июля 1973 г.). Важные моменты поведения Тараса в этом опыте, отснятые двумя киноаппаратами с разных точек, вошли в два кинофильма.2 В опытах, когда приманка (конфета, яблоко) опускалась на дно узкой и довольно глубокой ямки, Сильва воспользовалась ветками орешника, выломанными ею из куста, росшего от ямки в 11 м. До этого Сильва (3,5 лет) безуспешно испробовала много примитивных приемов, чтобы добыть лакомство из ямки (глубина 80 см) с помощью рук и ног. Любопытно, что из четырех шимпанзе, живших на острове, только Сильва пользовалась специально подготовленными палочками для доставания приманки из глубины. После короткого осмотра куста она выбирала подходящий ствол, пригибала его к земле и откусывала или отламывала от него несколько веток. С пучком зелени она направлялась к ямке, усаживалась и принималась за подготовку палочек. Для этого Сильва брала одну из веток и перекусывала ее в одном-двух местах, а получившиеся фрагменты очищала от листьев, небольших веточек и иногда коры. Из нескольких подготовленных таким образом палочек обезьяна почему-то брала только одну, другие бросала и приступала к доставанию приманки. Если палочка была неподходящей прочности или длины, Сильва снова шла к кусту и цикл подготовки повторялся. Очень важно, что во время первого опыта приманка добывалась только с помощью четвертой палочки, которая оказалась достаточной длины и прочности. После опыта все использованные палочки были нами собраны, измерены, описаны и сфотографированы (опыт от 15 и 18 июля 1973 г.). Другие шимпанзе для решения задачи с глубокой ямкой пользовались случайными предметами: сухими хворостинами или нетолстыми палками, найти которые в лесу не представляло особых трудностей. Вместе с тем после изучения ямки Тарас, Гамма или Бой быстро осматривали поляну, где до опыта специально все было чисто прибрано, и отправлялись к лесу. Они поднимали не все валявшиеся сухие ветки или палки, некоторые из них, подняв, бросали, а потом, остановившись на чем-то, поднимали этот предмет, удаляли лишнее и принимались за доставание. То обстоятельство, что шимпанзе подбирали с земли или отламывали от кустов и деревьев такие предметы, которые наиболее подходили по содержанию ситуации, а также быстро отбрасывали после короткого испытания менее подходящие, может указывать на их способность активно оперировать следовым образом и планом. Необходимо сказать, что в способности шимпанзе к подготовке предмета до того, как он использовался для решения конкретной задачи, или после одной-двух неудачных попыток такого решения мы убедились еще в лаборатории в процессе онтогенетического изучения у них орудийной деятельности в пищевой ситуации. В природе эта особенность их поведения проявлялась чаще. При этом шимпанзе не держали при себе изготовленного или уже использованного предмета. Каждый раз он выбирался вновь. Исключение представляет тот случай, когда Тарас принес весло, за которым он, скорее всего, пошел преднамеренно. Большей же частью можно было наблюдать обратное: приготовление нескольких палочек, а потом выбор из них только одной (Сильва), многократное последовательное изготовление полностью окоренных палочек для ловли муравьев (Сильва, Чита). Особенности обнаруженного поведения, конечно, нуждаются в анализе. Изучение орудийной деятельности шимпанзе в природных условиях позволило также сделать заключение о том, что свойства многих окружающих предметов достаточно полно и надолго запоминаются обезьянами и при необходимости используются вполне адекватно. В экспедициях 1978 и 1980 гг. мы имели возможность длительно наблюдать за поведением детеныша шимпанзе Боя-2, когда ему было 1,7—3,7 лет. В лесу он имел возможность общаться, кроме матери Сильвы, со всеми взрослыми шимпанзе, в том числе с могучим и агрессивным самцом Тарасом, который к своему сыну проявлял самые нежные чувства. В первой экспедиции, еще будучи детенышем, Бой-2 продемонстрировал способность к целенаправленному использованию природных объектов (хворостинка, палка). В экспедиции 1980 г. он оказался более мобильным, часто держался отдельно от матери, и можно было видеть, как он вначале делал грубые ошибки при подъемах по дереву, прыжках с ветки на ветку, при переходе с дерева на дерево. Однако эти промахи имели место только в начале экспедиции, затем же полностью исчезли, что указывает не только на обогащение его опыта знаниями об окружающей среде (трава, кусты, деревья, вода, палки, камни, животные и др.), но и на прочное запоминание этих знании. В течение многих лет в нашей лаборатории изучалась игровая, ориентировочно-исследовательская, подражательная деятельность, а также обобщающая функция, которые в экспериментах всегда связывались со способностью шимпанзе использовать различные предметы. Насколько эти обезьяны могут ориентироваться относительно того или иного предмета говорит метод «выбора по образцу», в свое время примененный Н. Н. Ладыгиной-Котс (1923, 1935). В опытах мы смогли убедиться в том, что подростки и взрослые шимпанзе способны зрительно идентифицировать объемные геометрические фигуры, изготовленные из дерева, с их изображениями в виде схем, выполненных на ватмане. В другом варианте работы обнаружилось, что шимпанзе без дополнительного обучения может осуществлять выбор на образец с интермодальным переносом со зрительного (эталон) на кожно-кинестезическую рецепцию (выбор объекта из набора) (Фирсов, 1972, 1973, 1977). В опытах, где специально преследовалась цель изучения предметной деятельности у низших и высших обезьян разного возраста, установлены важные закономерности поведения с использованием предметов, которые имели место у всех видов обезьян, невзирая на их возраст, во время игры, а также при обследовании предметов. У некоторых видов низших обезьян (капуцины, павианы и др.) и антропоидов, кроме этого, довольно часто проявляется использование предметов (щепочка, лучинка, листья, бумага, тряпка) для целей «самообслуживания»: целенаправленное же применение предмета с частичной его обработкой свойственно только антропоидам. Шимпанзе с определенного возраста оказываются способными соединить две короткие палки для того, чтобы преодолеть расстояние, большее, чем длина руки с каждой короткой палкой в отдельности. По поводу такого поведения шимпанзе в опытах В. Кёлера И. П. Павлов сказал: «Самым замечательным номером самой „умной" из его обезьян „Султана" было удлинение палки за счет двух палок, когда ни одна из палок в отдельности не доставала через решетку до банана» (Павловские среды, 1949, т. 2, с. 573). Наши опыты с составлением палок, проведенные на нескольких детенышах и подростках шимпанзе (Бой, Гамма, Тролль, Тарас, Бой-2), показали, что они ничем не отличаются от выдающегося Султана В. Кёлера (Kohler, 1930), чем опровергли этот «критерий гениальности» у шимпанзе. В первых пробах с палками или другими предметами, которые можно соединять, шимпанзе ведет себя, как обычно: обследует, играет, иногда почти сразу соприкасает предметы или даже стыкует их, но, как правило, достигнутого результата не оценивает (вероятно, в этом для него ничего нет необычного). В нашем фильме «Стратегия поиска в переменном лабиринте» (Институт физиологии им. И. П. Павлова АН СССР, операторы Ю. И. Левкович, В. Б. Лозовский, 1972 г.) имеются кадры, где показано отношение детеныша шимпанзе Тараса к длинной и двум коротким палкам. Длинной палкой он оперирует целенаправленно, и движения его точны. Когда же ему дали две короткие палки, которые можно торцами составить, то вначале он начал игру, в процессе которой обнаружил трубку на конце одной палки, запустил внутрь этой трубки палец, и, наконец, составил палку. Однако в следующий момент палки были разобщены, и Тарас, возбудившись, разбросал их. Подобная оценка результата сочленения оказалась и у других обезьян, что может говорить только об одном: соединение палок делается неосознанно и свидетельствует лишь о возможности шимпанзе осуществить эту операцию. Вне всякого сомнения, подобное поведение — бесцельное проявление возможности — свойственно также детям первых лет жизни. Вместе с тем достаточно нескольких проб (2 — 3), чтобы успешно составленная конструкция (например, из двух коротких палок) начала использоваться для решения конкретной задачи. В этом случае подросток шимпанзе, по-видимому, действует на основании хорошо развитого у него обобщения. Более того, сам факт составления объектов для дальнейшего применения в новой ситуации мобилизует внимание к дифференцированному набору предметов по «признаку соединимости». В наших экспериментах с шимпанзе 2 — 6-летнего возраста применялся набор предметов, состоящий из двух коротких палок А и Б и небольшого куска алюминиевой проволоки (прямого или согнутого в виде кольца). Обе палки имели поперечные сверления, расположенные нерегулярно вдоль их длины. Кроме того, палка А имела с обоих концов торцевые сверления, которых не было в палке Б. Так как этот набор давался обезьяне для возможного использования его в задаче с доставанием лакомой пищи, находящейся за пределами клетки, то было считанное количество вариантов, которые давали возможность сочленения предметов (палки и проволоки), и только один, который давал действительное удлинение конструкции и мог привести к успеху. Этим единственным способом было погружение конца проволоки в торцевое отверстие палки А. Все шимпанзе, которым предлагался такой набор (палки А, Б и кусок проволоки), умели совершенно точно оперировать длинной палкой для доставания предметов, лежащих вне клетки. Получив в свое распоряжение набор в первом опыте, Каролина, Тролль, Гамма, Бой, Тарас проявляли напряженную ориентировочно-исследовательскую деятельность с элементами деструкции (обкусывание палок, сгибание и поломка проволоки) или играли. Очень редко, взяв какой-либо предмет из набора, обезьяна тянулась в сторону приманки, но тут же оставляла эту попытку. Надо отметить, что у шимпанзе отличный глазомер, и обезьяна, еще не взяв предмета в руку, большей частью определяет, следует это делать или нет. Редкие попытки использования явно неподходящего предмета для решения задачи имеют скорее значение заместительной деятельности. Общая оценка использования обезьянами предметов указанного набора была положительной, и речь идет лишь о количестве испытаний, мотивированности и, возможно, возрастной характеристики шимпанзе. Так, Каролина, самая старшая из обезьян, после угашения игровой и ориентировочно-исследовательской реакции уже во втором опыте надежно соединила проволоку в торец палки А и достала приманку. Она ни разу не вставила проволоку в поперечные сверления палок и тем более не соединяла проволокой обе палки через поперечные сечения. Придание куску проволоки формы кольца также не тормозило ее деятельности; Каролина довольно быстро его расправляла, и большей частью получившуюся уплощенную дугу соединяла с торцом палки А. Любопытно, что при недостатке длины получившейся конструкции Каролина часто дополнительно разгибала конец проволочной дуги, чем достигала успеха в решении задачи (доставание пищевой приманки). По такому же типу шло освоение набора и применение составленной конструкции обезьянами Гаммой и Тарасом, но они показали более длительное угашение ориентировочно-исследовательской и игровой активности. Подростки Бой и Тролль достаточно долго дифференцировали палки А и Б, часто вставляли проволоку в поперечные отверстия палок, хотя такой конструкцией в дальнейшем почти не пользовались. Неуспех часто приводил к торможению операций с набором. Для повышения мотивированности обезьян приходилось экспериментировать через большие промежутки (неделю-две), а в день опыта — повышать пищевую возбудимость. Вполне возможно, что затруднение вызывалось еще недостаточным развитием подростков, однако со временем с этой задачей справились и они. На 10 шимпанзе в возрасте 2,5 — 15 лет исследовалось целенаправленное использование различных предметов с выбором из стандартного множества. Для этого в ситуации, когда приманка оказывалась вне клетки, обезьяне давался набор предметов и отмечался порядок их выбора для последующего использования. Следует еще раз подчеркнуть, что операции с палками всеми обезьянами выполнялись правильно и впервые обнаружились в возрасте около двух лет либо уже имелись у прибывших более взрослых шимпанзе (Рица, Каролина, Роза). Другой набор состоял из 11 предметов: длинная палка (80 см), «рогатина», короткая палка (30 см), кусок материи, кусок веревки, моток алюминиевой проволоки, палка А, палка Б, кусок алюминиевой проволоки, две короткие палки (по 30 см), годные для стыковки, кирпич. Во время испытаний этот набор предъявлялся обезьяне на полу вольера. Вне вольера на фанерном щите, выкрашенном в светло-серый цвет, укладывалась приманка на расстояние, чтобы ее невозможно было достать с помощью короткой палки, палок А или Б с учетом длины руки обезьяны. Эксперименты с выбором предметов проводились не чаще одного раза в неделю во второй половине дня, когда у обезьян имелась наиболее стабильная и высокая пищевая возбудимость. Использованный в текущем испытании предмет изымался. Полученные результаты удивительно сходны и отличаются только деталями. Во-первых, вне зависимости от возраста обезьяны брали в первую очередь длинную палку и быстро достигали успеха. Во-вторых, после изъятия из набора длинной палки, в зависимости от индивидуальных черт поведения, брались такие предметы, как моток проволоки (Тролль), «рогатина» (Каролина, Тарас), две короткие палки для составления в одну (Гамма), кусок материи, веревка (Бой). В-третьих, палки А или Б и кусок проволоки даже после значительного обеднения набора использовались неохотно; в-четвертых, короткая палка и кирпич никогда не использовались обезьянами и не только в том случае, когда эти предметы оставались к концу опыта, но и при выборе из двух предметов в самом начале эксперимента. Подробный анализ фактов, полученных в опытах с выбором предмета из множества, показывает, что шимпанзе на основании зрительной оценки множества — и, вероятно, собственной практики — способен классифицировать предметы по их функциональным качествам. Именно поэтому, нам кажется, короткая палка и кирпич игнорировались полностью с начала и до конца опыта. Следует к этому добавить, что после изъятия из множества наиболее удобного предмета (длинная палка) у шимпанзе наступала значительная генерализация. Так, например, Тролль чаще избирал моток проволоки, но он мог взять и кусок материи или две палки для составления, «рогатину» и т. д. Это же в равной мере относится и к другим обезьянам, имевшим свои изначальные склонности. Вместе с тем каждая из обезьян, кроме длинной палки и других предметов, которыми так или иначе можно было дотянуться до приманки, не использовали ни короткой палки, ни кирпича. За последний, впрочем, обезьяна могла хвататься, когда в результате возникшего затруднения она проявляла признаки недовольства или ярости. Одним из конструктивных подходов для объективного изучения предметной деятельности животных, в частности обезьян, мы считаем онтогенетический анализ этой формы поведения. Для проведения его мы предлагали стандартные задачи низшим и высшим обезьянам. Использование предметов антропоидами разного возраста изучалось в таких задачах, как доставание подвешенной приманки, выталкивание приманки из плексигласовой трубы, доставание лакомства из прозрачных вертикально поставленных сосудов с разным диаметром входного отверстия и др. Основная цель этих опытов состояла в изучении возрастного фактора испытуемых, а также процесса обработки (приспособления) предмета для дальнейшего его применения в экспериментальной ситуации. Первые и последующие опыты, поставленные на Каролине, Тролле, Гамме и Бое, показаны нами в кинофильмах «Думают ли животные?» и «Изучение орудийной деятельности у человекообразных обезьян (шимпанзе) разного возраста». Как и предполагалось, лидировала во всех испытаниях 4-летняя Каролина. Она с большим успехом делала лучины, обкусывая край тонкой доски (обязательно вдоль волокон), заостряя ее конец, что несомненно способствовало накалыванию кусочков фруктов, лежащих в сосуде. Несколько хуже действовал Тролль, возбуждавшийся и тративший на решение несколько больше времени, а детеныши Гамма и Бой при обилии всевозможных предметов пользовались в основном только собственными конечностями. Поэтому они успешно доставали сласти из вертикального сосуда, куда проникали их руки, но не могли ничего добыть из узкогорлого сосуда с помощью лучины. Таким образом, шимпанзе, достигший известного возраста (развития головного мозга), способен в окружающей среде обнаружить и выделить определенные предметы. Он достаточно точно дифференцирует их по потенциальным качествам, соединяет в устойчивые конструкции или, наоборот, разъединяет, а также частично подготавливает. Самое главное, что, вероятно, определяет предметную деятельность антропоидов — это способность обобщить несколько предметов в подклассы, а последние свести в классы. Это отчетливо можно видеть в экспериментах с выбором из множества. Метод исключения позволил нам убедиться, что шимпанзе, обобщая различные предметы, делит их на два класса: достаточные для овладения приманкой и недостаточные для этого. В опытах на низших обезьянах мы не смогли отметить целенаправленного использования предметов. Об этом же свидетельствуют и литературные данные. По-видимому, уровень предметной деятельности антропоидов следует расценивать как качественно новую форму поведения, возникшую в эволюции отряда приматов. Представляется, что гипотеза В. П. Якимова (1972) об австралопитеках, прямоходящих высших обезьянах, систематически использовавших орудия, в высшей степени ценна тем, что указывает на качественный уровень использования природных объектов как на устойчивый таксономический показатель. Трудно представить, как можно было бы оторвать целенаправленную предметную деятельность от самого существа ситуации, на которую антропоид активно воздействует. Разумеется, техника использования какого-либо предмета, с одной стороны, отражает собственно моторные возможности обезьяны, с другой — выявляет конкретные причинно-следственные отношения, которые предполагаются программой эксперимента или даже открываются самим испытуемым. Для иллюстрации сказанного приведем попытку воспроизведения «опыта с огнем», поставленного сотрудниками И. П. Павлова на Рафаэле (Штодин, 1947; Вацуро, 1948). Об этом опыте, впервые осуществленном П. К. Денисовым, подробно рассказывал И. П. Павлов на «Средах» (9 января и 6 марта 1935 г.). Уже тогда было отмечено, что навык тушения огня у Рафаэля прочно укрепился. Укрепление это шло и при дальнейшей работе с Рафаэлем, где практиковалось «тушение огня» в системе навыков. Именно эта длительность тренировки и хорошо известная способность обезьян устанавливать мощные кинестезические связи (Вацуро, 1948, 1952) заронили у нас сомнение в корректности выводов, сделанных на основании «опытов с огнем», поставленных на плотах. Этот хорошо известный опыт, по мнению Э. Г. Вацуро, будто бы показывал, что у шимпанзе нет общего представления о воде и что Рафаэль отчетливо различает воду бака, из которого он брал ее для тушения огня в проблемном аппарате, и любую другую воду, в том числе окружавшую плоты. В критическом эксперименте Рафаэль для тушения не зачерпнул воды из озера, что технически не представляло никаких трудностей, а с риском упасть в озеро навел переправу с одного плота на другой и принес воду в кружке, нацедив ее из знакомого бака. Так как воссоздание «опыта с огнем» делалось для кинофильма «У порога сознания» (режиссер Г. А. Бруссэ, «Леннаучфильм», 1954 г.), то мы решили воспользоваться этой возможностью, чтобы проследить за процессом ассоциирования в цепи событий таких предметов, как вода в баке — кружка — огонь, чтобы сделать контрольное испытание поведения шимпанзе с тушением огня на плотах. Для эксперимента был взят подросток шимпанзе Лада, отличавшаяся, как и Рафаэль, повышенным аппетитом и достаточной уравновешенностью. Результаты этого проверочного эксперимента вскрыли две особенности поведения антропоида. Во-первых, оказавшись в ситуации, где был бак с водой, кружка и аппарат с вкусной приманкой, доступ к которой преграждала завеса огня, Лада направила свое внимание на физические свойства аппарата, кружки и даже огня, а не на связь вода — огонь. К этому заключению можно прийти, проанализировав четыре решения в течение первого эксперимента, каждое из которых увенчалось успехом — добычей вкусного фрукта. Они заключались в следующем: а) опрокидывание аппарата с огнем; б) схватывание приманки рукой через завесу огня; в) укладывание пустой кружки на горящий валик из ваты и доставание приманки через образовавшуюся щель; г) удаление из аппарата металлической детали, в которой горел спирт. Лада переходила от одного решения к другому вследствие нашего запрета повторять предыдущее действие. Как видно, связь вода — кружка — огонь здесь отсутствует, хотя сама обезьяна обнаружила связь кружка — огонь. Во-вторых, оказавшись на плотах, Лада, уже имевшая навык наливать воду из бака в кружку и тушить огонь в аппарате, в контрольном испытании свободно зачерпнула кружкой воду из озера и залила огонь в аппарате. Сложную картину поведения Рафаэля с использованием предметов на каждом этапе описал И. П. Павлов. Она представляла собой ассоциацию нескольких навыков, которые уже были по отдельности образованы у Рафаэля в прошлых экспериментах. В контрольном опыте ему предлагалась задача на синтез этих навыков, с чем он превосходно справился. Цепь действий оказалась следующей: открывание ключом дверного замка — тушение огня на пороге выхода из комнаты на площадку — постройка на площадке вышки из нескольких ящиков — подъем на вышку и доставание плода. Комментируя этот опыт, Павлов сказал: «Совершенно ясно, до какой степени все это есть большая умственная работа» (Павловские среды, 1949, т. 3, с. 120). Итак, ключ, кружка с водой, постройка вышки — действия с предметами, благодаря которым антропоидом выполняется «умственная работа». Рука (и реже нога), следовательно, уже изначально вооружается предметом для чего-то, преследуя ближние или дальние, к сожалению, не всегда ясные экспериментатору, цели. По мнению Фишеля (Fischel, 1973), эксперименты, «что проводились с палками или ящиками, можно считать устаревшими» (с. 129). Он говорит об отсутствии количественной оценки поведения в ситуациях с выбором, что, конечно же, снижает их объективность. При этом для сравнения Фишель все же приводит опыты с палками и ящиками, поставленными с положительными результатами на капуцинах. Однако, несмотря на явную симпатию к низшим обезьянам, он утверждает: «...нельзя сказать, что мы имеем здесь дело с первичным решением задачи. Несомненно, в полученном результате на опыт падает большая доля, чем на понимание» (с. 133). И далее: «Исследование низших обезьян в эксперименте с палками и граблями показало, что они значительно уступают шимпанзе» (с. 133). Вывод его таков: «Сегодня можно с уверенностью сказать, что все низшие обезьяны обладают ограниченным, очень ограниченным пониманием» (с. 134). Таким образом, в некоторых работах западных и советских исследователей имеется усреднение низших и высших обезьян под единым и безликим термином «обезьяна». При этом важно привести мнение американского антрополога Ш. Уошберна (Washburn, 1967), что шимпанзе является незаменимым объектом для психологических исследований. Физиологический анализ орудийной деятельности невозможен без изучения меры участия в ней механизмов подражания различных видов памяти и функции обобщения (абстрагирования). Как подтверждают наши наблюдения, а также эксперименты в лабораторных и природных условиях, свойства отдельных предметов шимпанзе запоминает в совершенстве и надолго. При этом антропоиды обобщают не только содержание ситуаций, но и различные предметы, которые они в них использовали, обнаруживая в избираемых объектах качества, необходимые для решения проблемы (вес, длина, прочность, эластичность и пр.). Вполне вероятно, что обнаруженный уровень орудийной деятельности отражает способность шимпанзе и других антропоидов к более глубокому, по сравнению с другими животными, анализу связи предметов и явлений, что можно отнести за счет ароморфозных изменений головного мозга этих обезьян. Тот факт, что «большие полушария у обезьян развились больше, чем у других, причем развились именно в связи с разнообразием двигательных функций», был отмечен в свое время И. П. Павловым (Павловские среды, 1949, т. 2, с. 432). Напомним, что Н. А. Рожанскому (1957) принадлежит хорошо аргументированная классификация этапов развития коры головного мозга у позвоночных животных. Мозг обезьян он отнес к четвертому типу по таким морфологическим признакам, как наличие Роландовой борозды, лобных долей и др. При этом он указал также на ряд функций, которые заметно отличают обезьян от других млекопитающих: подражание, мимика, манипуляционная деятельность (точнее исследовательская), использование предметов. К сожалению, автор подошел ко всем видам обезьян обобщенно. В монографии А. И. Карамяна (1976) эти данные Н. А. Рожанского получили новое развитие на основании оригинального фактического материала. К этому следует добавить, что еще со времен В. М. Бехтерева стали известны факты сходного топического представительства в двигательной коре человека и антропоида большого пальца руки, равно как и движение отдельных пальцев передних (верхних) конечностей в ответ на электрическое раздражение моторной области полушарий. Нейроморфологам известно, что мозг человека построен по плану обезьяньего мозга. Работы последних десятилетий не только подтвердили этот вывод, но открыли в мозговой коре антропоидов (шимпанзе) поля 39 и 40, которые считались ранее характерными только для человека. Этот важный факт установлен Ю. Г. Шевченко (1971) на основании сравнительно-морфологического изучения нижнетеменной области коры игрунки, мартышки, гиббона, шимпанзе и человека. Значительно отличие площадей важнейших корковых областей у антропоида и человека. Так, площадь лобной коры по отношению ко всей коре у человека составляет 24,4%, у шимпанзе — 13,5%, площадь коры нижнетеменной области соответственно — 7,7 и 3,3%. В связи с большой долей участия в орудийной деятельности функции зрительного анализатора следует обратить внимание на то обстоятельство, что в процессе антропогенеза затылочная кора также претерпела значительные изменения. Так, у человека поле 19 больше 17-го. Поле 19 составляет у человека, орангутана и мартышки 4,5; 6,0 и 6,4% от площади всей коры, тогда как поле 17 занимает соответственно 3,0; 8,5 и 10,0%. Особенность поведения шимпанзе, связанная как с более широким использованием зрительной функции, так, по-видимому, и памяти, была замечена также И. П. Павловым. При разборе опытов с открыванием проблемного ящика при помощи палок-«ключей» разного сечения он не исключал, что «обезьяна в некоторых благоприятных условиях пользуется уже и зрительным образом» (Павловские среды, 1949, т. 2, с. 297). Измерение слепков эндокранов древнейших и древних гоминид убеждает, что первоначальное бурное развитие задних отделов мозга с их сенсорными функциями сменяется в дальнейшем прогрессивным увеличением передних его частей, в особенности префронтальной зоны, где, как показывают нервная клиника и эксперимент на животных, программируется сознательное, целенаправленное поведение (Лурия, 1969; Кочеткова, 1973; Батуев и др., 1979; Фирсов и др., 1979). По представлениям В. П. Якимова (1964), Робинсона (Robinson, 1972) и др., в эпоху третичного и начала четвертичного периодов наметился отчетливый процесс радиации австралопитековых на парантропов и австралопитеков. И если первым, тоже уже двуногим антропоидам, была свойственна в основном растительная диета и лишь возможное использование внешних предметов в виде орудия, то для плотоядных австралопитеков орудийная деятельность оказалась важным таксономическим критерием. По-видимому, здесь трудно было бы наметить критическую стадию перехода от предметной к орудийной деятельности, вероятно, она лежит, с одной стороны, на пути постепенного освобождения рук от опорной функции и взаимного влияния, с другой — развивающегося мозга и органов чувств и утончения моторных действий кисти. Именно особое устройство кисти шимпанзе создает возможность для обширного оперирования различными предметами. Как известно, схема Нэпира (Napier, 1963, 1967) показывает орудийную деятельность древнейших гоминид, однако три первых из шести стадий отчетливо усматриваются в поведении шимпанзе и, вероятно, это тождество, а не натянутая аналогия: первая — случайное использование орудий, вторая — намеренное использование орудий, третья — формирование орудия для непосредственного использования. Все стадии в Африке видели у шимпанзе Кортланд (Koortlandt, 1974), Дж. Лавик-Гудолл (Lawick-Goodall, 1974), а мы — на озерных островах Псковской области (Фирсов, 1977). В этом отношении, конечно, Дарт (Dart, 1957) правильно выделил эпоху «кости, зуба, рога», которая предшествовала каменной и, вероятно, сменила эпоху палки и других случайных природных объектов, не оставивших после себя никаких археологических следов. В этом аспекте представляет интерес пятая ступень классификации орудийных действий древнейших гоминид, предложенная В. В. Бунаком (1980): изготовление немногих стандартизованных форм орудий. Стандартизованность, или стереотипность изготовления орудий, например, олдувайского типа, скорей всего отражает повышенную прочность «социального наследования» (Дубинин, 1977) между поколениями древнейших гоминид, которые вряд ли могли полностью игнорировать другие внешние предметы. В этой связи полиморфность каменных орудий, которая расценивается С. А. Семеновым (1964) как показатель недостаточного интегрирования нервной деятельности, может говорить о том, что у homo habilis и более поздних гоминид функция обобщения благоприятствовала функциональному сглаживанию второстепенных признаков предмета либо, наоборот, использованию их отличительных признаков в целях ближайших потребностей. Все сказанное нисколько не реабилитирует представителей отряда приматов относительно их склонности к стереотипизации поведения, не исключая в огромной массе и человека разумного. Вполне возможно, что на протяжении антропогенеза, еще из глубин миоцена, это качество высшей нервной деятельности в какой-то степени умеряло все нарастающую способность антропоидов к пластическим формам поведения. Можно предположить, что иерархия этих двух тенденций психонервной деятельности обезьян и человека является фундаментальным механизмом адекватного или неадекватного (патологического) отражения действительности. Циркулярность, стереотипия, навязчивость, застойность, как известно, являются ведущими синдромами многих невротических и психических расстройств. Против приведенного мнения С. А. Семенова (1968) говорит и то обстоятельство, что после эпохи «немногих стандартизованных форм орудий» (Бунак, 1980) наблюдается как бы ее распад с обнаружением в камне многих свойств, до того не обнаруженных. Этот период орудийной деятельности можно трактовать как очередной перевес пластичного регулирования, несомненно связанного с накоплением морфологических сдвигов в структурах головного мозга, над стереотипными формами орудийного поведения, т.е. очередным прогрессивным шагом в процессе антропогенеза. В развитие наших представлений о функции обобщения у приматов (низшие и высшие обезьяны, ребенок) поведение шимпанзе и макак резусов исследовалось в процессе использования и уподобления ими по эталону природных объектов. На основании множества наблюдений и экспериментов, удостоверенных тщательной киносъемкой для многих кинофильмов, мы пришли к выводу, что предметная деятельность антропоидов в природных условиях отличается высокой пластичностью и целенаправленностью. При этом было обнаружено важное обстоятельство, что при неуспехе первого решения мы не видели, чтобы последующие действия шимпанзе были еще более ошибочными. В различных проблемных задачах на преодоление расстояния или тяжести, а также при выборе предмета по тождеству, шимпанзе использовали природные объекты в самом общем виде, дифференцируя их до начала действий с ними, в соответствии с содержанием ситуации (Фирсов, 1972, 1977; Фирсов и др., 1979). Подобное поведение шимпанзе Рафаэля отмечено И. П. Павловым в его первой информации об «опыте с огнем»: «Когда воды в кране не оказалось, он взял бутылку с водой и вылил ее на пламя. Чего вам надо больше?» (Павловские среды, 1949, т. 3, с. 17). Не может быть никакого сомнения, что в данном случае мы имеем дело не только с целенаправленным поведением, орудийностью его, но также с обобщением предмета в плане ближайшей потребности. Следовательно, предметная (до двухлетнего возраста) и орудийная деятельность шимпанзе являются генетически запрограммированными формами поведения, соответствующими филогенетическому и возрастному развитию их головного мозга. Таким образом, на основании собственных многолетних исследований предметной деятельности у разных видов низших и высших обезьян, проведенных в лабораторных условиях, а также в природных условиях Псковской, Ленинградской областей и Абхазской АССР, мы пришли к принципиально важному для нас заключению о качественно новом уровне этой деятельности только у антропоидов, т.е. об орудийном характере этой деятельности. По-видимому, ее можно рассматривать как основу животнообразных форм труда. Последовательный эволюционный, этологический и онтогенетический анализ способности животных к использованию природных объектов, думается, будет полезен для дальнейшей разработки гипотезы о становлении трудовой деятельности древнейшего человека. Сложные формы операций антропоидов, в особенности шимпанзе, с предметами, широкое их обобщение, а также обобщение ситуаций, в которых они применяются, целенаправленность этих действий и несомненное владение следовым образом и планом (инициацией) предстоящего использования, убедительно говорят о том, что орудийная деятельность возникла задолго до появления древнейшего человека и даже до австралопитеков и ныне живущих антропоидов (Yerkes, 1943; Семенов, 1964; Parker, 1968; Алексеев, 1972; Фирсов, 1972, 1977; Lawick-Goodall, 1974, и др.). На этом основании знаменитая фраза И. П. Павлова: «Когда обезьяна строит свою вышку, чтобы достать плод, то это „условным рефлексом" назвать нельзя» (Павловские среды, 1949, т. 3, с. 262), приобретает особо важное значение. Речь идет не о проявлении элементарного условного моторного акта, а о системной деятельности высокоорганизованного мозга со сложнейшей интеграцией функций многих церебральных механизмов. Мнение, высказанное Н. П. Дубининым (1977), о том, что животные предки человека, опираясь на итоги генетической эволюции, придали процессу труда целенаправленный характер, по-видимому, следует понимать таким образом, что целенаправленность может иметь как тактическое, так и стратегическое значение. Целенаправленность действий человека, актуализирующая сознательно построенный план и поддерживаемая критической оценкой уже сделанного, действительно, присущи только ему и являются стратегией трудовой деятельности. Вместе с тем целенаправленность использования предметов внешней среды, а также способность к частичному изменению этих предметов, является неотъемлемым новым качеством поведения ребенка первых лет жизни и высших обезьян (антропоидов).
ИССЛЕДОВАНИЕ ОРУДИЙНОЙ ДЕЯТЕЛЬНОСТИ КАК ПУТЬ К ИНТЕГРАЛЬНОЙ ОЦЕНКЕ КОГНИТИВНЫХ ВОЗМОЖНОСТЕЙ ЖИВОТНЫХ Ж.И. Резникова
Институт систематики и экологии животных со ран; Новосибирский государственный университет. Опубликовано: Журнал общей биологии, 2006, n1
В последние три десятилетия в этологии выделилась в качестве отдельной области когнитивная этология, изучающая познавательные процессы у животных и опирающаяся во многом на методы и подходы психологии (Griffin, 1976, 1992; Allen, Bekoff, 1997; Shettleworth, 1998; Резникова, 2000, 2005 [см. Введение и глава 8]; Bekoff, 2002). Проблемы когнитивной этологии связаны с наиболее сложными психическими процессами у животных и имеют непосредственное отношение к поиску эволюционных корней интеллектуальной деятельности человека. В течение столетий использование орудий считалось одним из самых надежных поведенческих признаков, отличающих человека от других биологических видов. Правда, внимательные наблюдатели давно обращали внимание на то, что животные могут использовать в своей деятельности разные предметы. Наблюдения за обезьянами, раскалывающими камнями орехи, и слонами, которые прогоняли ветками мух, встречаются в средневековых книгах. Дарвин (Darwin, 1871) привлек внимание научной общественности к использованию орудий животными и показал, что человек – не единственный вид, манипулирующий предметами для достижения цели.
С развитием этологии, во второй половине XX-го столетия, список видов, к которым приложимо понятие орудийной деятельности, был существенно расширен. Появилось множество вопросов: почему представители одних видов используют орудия, а других видов - нет; насколько животные компетентны в свойствах и связях предметного мира, в какой мере сказывается на проявлении орудийной активности животных индивидуальный и социальный опыт, и насколько она определяется генетической программой. Автору пришлось столкнуться с этими проблемами при обсуждении связи между коммуникативным и орудийным поведением муравьев, в дискуссиях, неоднократно возникавших на этологических конференциях (Reznikova, 1995, 2001; Reznikova, Ryabko, 1997).
Несмотря на то, что орудийное поведение животных неоднократно рассматривалось в монографических сводках и учебниках (Гудолл, 1992; Beck, 1980; McGrew, 1992, 2004; Резникова, 2000, 2005 [см. Введение и глава 8]; Зорина, Полетаева, 2001), задача анализа накопленных результатов в рамках парадигмы когнитивной этологии остается актуальной. Исследование орудийной деятельности является замечательным инструментом для экспериментаторов, изучающих пределы интеллектуальных возможностей животных. Использование этого подхода порождает все новые продуктивные гипотезы в области когнитивной этологии и сравнительной психологии. Пояснением может служить использование языков-посредников для изучения интеллекта животных, прежде всего, человекообразных обезьян (Gardner B., Gardner R., 1969; Savage-Rumbaugh,1986), а также попугаев (Pepperberg, 1987) и дельфинов (Herman, 1986). Вступив в “диалог” с животными, исследователи сумели не только оценить потенциал их коммуникативных возможностей, но и получить непосредственные “ответы” обезьян и попугаев на вопросы, касающиеся свойств, формы, количества предметов. Однако в диалог можно вступить только с потенциально “говорящими” видами, т. е. с такими, которым можно предложить адекватный язык-посредник. Представители “умелых” видов предоставляют исследователям дополнительные возможности для познания пределов когнитивных способностей. Решая поставленные экспериментаторами задачи с помощью орудий, животные дают возможность наблюдателю судить о том, как они выбирают предметы для изготовления орудий, как оценивают их свойства, предвидят ли результаты своих действий, улавливают ли закономерности перемещения предметов в пространстве и во времени.
В обзоре, на базе краткого описания феноменологии орудийной деятельности, проанализированы экспериментальные исследования когнитивных аспектов орудийного поведения животных. Систематизация последних достижений в данной области знаний дает основания рассматривать орудийное поведение животных как эффективный методологический инструмент для интегральной оценки когнитивных возможностей ряда биологических видов.
ТЕРМИНОЛОГИЯ И КРАТКОЕ ОПИСАНИЕ ОРУДИЙНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ В ЕСТЕСТВЕННЫХ УСЛОВИЯХ
Элкок (Alcock, 1972) предложил следующее определение орудийной деятельности: использование орудий состоит в манипуляциях с каким-либо неодушевленным предметом, созданным вне собственного организма животного и повышающим эффективность его действий, направленных на изменения положения или формы какого-нибудь другого предмета. Гудолл (Goodall, 1970) дает более краткое определение: использование тех или иных предметов внешнего мира в качестве функционального продолжения какой-либо части тела для достижения ближайшей цели. Бек (Beck, 1980) в книге “Орудийное поведение животных: использование и изготовление орудий” дает, в общем, сходное определение: использование объектов внешнего мира для изменения положения, формы или состояния других объектов, при этом пользователь удерживает предмет и контролирует его ориентацию и эффективность действия.
В терминологии, относящейся к данной области этологии, есть элементы неопределенности и разночтений, неизбежные для развивающейся ветви экспериментальной науки. В частности, нечетко определены различия между орудийной и конструктивной видами деятельности животных. В некоторых случаях границы между понятиями расплывчаты, поскольку весьма условны границы между “долговременными” и “ближайшими” целями в поведении животных. Иные, до сих пор не совсем ясные случаи использования предметов, такие, как “захоронительное” поведение слонов, забрасывающих ветками мертвых животных и подозрительные для них предметы (муравьи поступают так же с пятнами жидкого или вязкого субстрата), видимо, можно отнести к манипуляторной активности.
Н.Н. Ладыгина-Котс (1959) рассматривала в качестве конструктивной деятельности манипулирование предметами и изготовление сооружений (гнезд), а в качестве орудийной – использование предметов для достижения цели. При этом сложные формы орудийной деятельности включают подготовку и преобразование предметов, т. е. изготовление орудий. Мы будем придерживаться этой терминологии, тем более что она соответствует и устоявшимся в англоязычной литературе терминам: tool using (использование орудий) и tool manufacture (изготовление орудий).
Орудийная деятельность включает как сложные действия, сопровождаемые модификацией предметов, так и использование простых “подручных” средств для достижения цели. В последнем случае имеется в виду использование предметов, не измененных деятельностью животного: так, обезьяны почесывают спину палочкой или используют камень, чтобы расколоть орех. Такие предметы называют “натурофактами”, противопоставляя их артефактам, т.е. предметам, являющимся результатом “направленной деятельности”, таким, например, как прутик, заостренный зубами (Beck, 1980).
Изготовление орудий (toole manufacture) является одним из самых сложных проявлений когнитивной деятельности животных. Различают четыре различных по сложности способа изготовления орудий. Первый - самый простой и распространенный среди животных – отщепление (detach). Примером может служить сорванная ветка, которая используется без преобразования, скажем, для того, чтобы согнать муху или швырнуть во врага. Второй способ – редукция (subtract). Ветка, очищенная от листьев, изготовлена путем редукции. Такое орудие можно использовать, скажем, для “ужения” муравьев или термитов. Если же используется не сама ветка, а сорванные с нее листья (например, чтобы стереть кровь или грязь), то листья – это орудие, изготовленное первым способом, т. е. путем “отщепления”. Третий способ - комбинирование предметов (combine). Примером могут служить вставленные одна в другую палки. Четвертый, самый сложный из наблюдаемых у животных,- это преобразование (reshape). Этот способ требует от животных “представлений” о свойствах предметов, позволяющих им изготовить функциональное орудие. Сами действия могут быть несложными, важно, что они опираются на понимание животными причинно-следственных связей. Обезьяны, которые мнут и жуют листья, чтобы увеличить их адсорбирующие свойства и использовать как губку для добывания воды из дупла, используют преобразование предметов (Beck, 1980; McGrew, 2004).
Рассмотрим разнообразие примеров использования орудий представителями разных видов животных в ситуациях, близких к естественным, без вмешательства наблюдателей.
Использование орудий млекопитающими. Африканские слоны (Loxidonta africana) и индийские слоны (Elephas maximus) используют орудия в разных ситуациях: они с помощью хобота бросают ветки и камни в других животных и в наблюдающих за ними людей; используют ветки и палки, чтобы почесать труднодоступные части тела; вытирают пучками травы кровь и затыкают раны травой; отгоняют ветками мух (Douglas-Hamilton I., Douglas-Hamilton O.,1975; Moss, 1982; Chevalier-Skolnikoff, Liska, 1993;Masson, 1995). Использование опахал в борьбе с мухами является повседневной формой активности, и примечательно, что слоны обрабатывают ветки прежде, чем пустить их в дело. Индийские слоны, сорвав большую ветку, придерживают ее на земле передней ногой и с помощью хобота отламывают или откручивают часть нужного размера (Hart et al.,2001).
Опубликовано несколько примеров использования орудий грызунами. Самка гофера (Thomomys bottae) использовала камешки и затвердевшие кусочки пищи для выкапывания норы. Орудие служило ей как для разрыхления, так и для отгребания почвы (Katz, 1975). Самка мыши-малютки Micromys minutus после ряда безуспешных попыток выбраться из своего аквариума нашла решение, вскарабкавшись по стеблю овсюга, который находился в ее жилище. Позже она научилась прислонять стебель к стенке аквариума, так что он служил ей устойчивой лестницей (Zimmerman, 1952 - цит. по: Beck, 1980). Содержащиеся в неволе суслики Spermophilus beecheyi регулярно швыряли песок в приближающихся к ним змей, действуя передними лапами (Owing, Coss, 1977).
Каланы, или морские выдры (Enhydra lutris), демонстрируют сложные и многообразные приемы в использовании орудий. Фишер (Fisher, 1939) описала поведение калифорнийских каланов, которые подбирали камни со дна и использовали для того, чтобы отколоть от скал прикрепленных к ним моллюсков. В этой же работе было впервые указано на то, что морские выдры используют морские водоросли, обматывая их плети вокруг туловища, что позволяет им спокойно, не затрачивая усилий, удерживаться на одном месте. В последующих публикациях орудийное поведение каланов описывалось все более детально (Kenyon, 1969; Houk, Geibel, 1974; Дежкин, Мараков, 1968; Смирин В., Смирин Ю., 1991). Оказалось, что если калифорнийские каланы постоянно пользуются камнями для добывания пищи, то северные прибегают к орудиям редко и открывают раковины зубами. Молодые северные каланы, не справляющиеся с открыванием раковин, а также звери с поврежденными зубами помогают себе камнем. Взрослые выдры иногда играют, стуча камнями по скалам. Калифорнийские каланы не имеют себе равных среди животных по тщательности и по количеству операций с камнями. Чтобы расколоть раковину, звери держат ее передними лапами и стучат ею о камень, находящийся на груди. Требуется до 20 ударов, которые наносятся сериями со скоростью 2 удара в 1 с. Один из самых длинных наблюдаемых “пищевых сеансов” калана занял 86 мин, в течение которых было съедено 54 моллюска, а понадобилось для этого 2237 ударов камнем (Hall, Schaller, 1964). Один и тот же камень используется до 12 раз подряд, калан плавает с ним, держа под мышкой или в складках кожи. Животные, по-видимому, учитывают потерю веса камня под водой: для ныряния с “молотком” выбирают камни весом 800-3500 г, тогда как на поверхности воды, в качестве наковальни берут более легкие (400-650 г). Наковальня чаще всего нужна для раскалывания двустворчатых раковин, но иногда ее используют и для того, чтобы вскрыть панцири крабов, лобстеров и оболочку крупных морских ежей.
Медведи привлекают внимание исследователей орудийного поведения тем, что эти животные ловко балансируют на двух задних ногах и могут использовать свободные передние лапы. Большинство наблюдений орудийного (например, использование палок для сбивания плодов с деревьев, растущих рядом с клеткой), а также игрового манипуляторного поведения медведей сделано в зоопарках (обзор см.: Beck, 1980). В естественных условиях зафиксировано использование камней и глыб льда белыми медведями для убийства ластоногих (Перри, 1974).
Подавляющее большинство данных об орудийной деятельности животных в естественных условиях относится к приматам. Накоплено множество наблюдений за тем, как капуцины Cebus apella и C. capucinus , макаки -крабоеды и южно-африканские бабуины используют камни для разбивания орехов, фруктов с твердой кожурой, яиц, раковин найденных на берегу моллюсков, а также пытаются разбить орехи, стуча ими один о другой (Struhsaker, Leland, 1977; Beck, 1980). Для того чтобы зафиксировать регулярное проявление сложного орудийного поведения, требуются многолетние наблюдения. Так, в одном из национальных парков Сан-Пауло в популяции C. apella наблюдалось регулярно повторяющееся использование обезьянами пар камней (наковальня и молот) для раскалывания орехов пальмового дерева (Ottoni, Mannu, 2001). Для японских макак использование камней удалось зафиксировать в локальной популяционной группировке непосредственно с первого случая и пронаблюдать в течение 20 лет (Huffman, Nishie, 2001). Более простые действия, такие, как швыряние в нарушителей личного пространства палками, камнями, орехами, песком, а также фекалиями и горстями мочи, описаны для капуцинов, макак, ревунов, паучьих обезьян, колобусов, лангуров, бабуинов и некоторых других видов. Макаки-крабоеды в ботаническом саду Сингапура регулярно используют листья для того, чтобы удалять с фруктов муравьев, грязь и грибной налет (Chiang, 1967). В одном из национальных парков Кении верветки (Cercopithecus aethiops) употребляют листья в качестве губок, чтобы достать воду из углублений (Hauser, 1988).
Наиболее сложные проявления орудийной деятельности описаны для человекообразных обезьян. В начале XX-го века наблюдения Кёлера (Köhler, 1925) и Н.Н. Ладыгиной-Котс (1923) за шимпанзе в неволе привлекли внимание исследователей к тому, как часто обезьяны манипулируют предметами в самых различных целях. Например, животные использовали острые палочки и проволочки для доставания насекомых из щелей; шесты для того, чтобы достать высоко висящий предмет; палку как рычаг для открывания крышки люка; они почесывали тело палочками, камнями, черепками посуды; чистили острыми палочками и скрученной бумагой ногти и уши; использовали листья и куски ткани для оборачивания рук, если приходилось действовать шершавой палкой; употребляли листья, тряпки и бумагу для того, чтобы вытирать грязь и кровь со своего тела. Содержащиеся в вольере обезьяны употребляли предметы (палки, проволока, камни) и в качестве орудий воздействия на других животных. Они подманивали кур, швыряя куски хлеба, тыкали в них палкой или проволокой, бросали в кур камни, избивали палкой ящериц и с большим интересом наблюдали за реакциями объектов издевательства.
Первые систематические знания о том, как приматы используют орудия в естественной жизни, были добыты Ниссеном (Nissen, 1931). Дж. Гудолл (1992), и У. Мак-Грю (McGrew, 1992, 2004) на основании многолетних исследований создали настоящую энциклопедию орудийной деятельности антропоидов. В обобщающей статье коллектива авторов, посвященной “материальной культуре” шимпанзе (Whiten et al., 1999), перечислены способы использования орудий в естественных условиях. В списке присутствуют 39 позиций, из которых более половины связано с добыванием пищи с помощью веток (накалывание, выуживание, метание в жертву), листьев (использование как губок) и камней (метание, удары по раковине, панцирю или скорлупе).
Приматологи отмечают существенные различия в “суммах технологий” у разных видов антропоидов. В естественных условиях бонобо, гориллы и орангутаны практически не применяют орудий. Единственный пример регулярного применения орудий у орангутанов касается популяции, обитающей в национальном парке Суматры (Schaik, Dopyera, 1997). Здесь в пище орангутанов видное место занимает плод с местным названием “пувин”, покрытый снаружи чем-то вроде стекловолокна, так что его нельзя раскусить, не поранив язык и губы. Обезьяны научились преодолевать эту трудность, проделывая отверстия в плодах острыми палочками и вынимая с помощью тех же палочек кусочки сердцевины с семенами. Авторы объясняют наличие локального “очага культуры” в исследуемой популяции прежде всего ее необычно высокой плотностью, позволяющей распространение и закрепление орудийных навыков в сообществе. Нужно, однако, отметить, что в группах бонобо и горилл, даже при более высоких показателях плотности популяций, “очагов культуры” пока не выявлено. При этом не вызывает сомнений способность представителей всех трех видов манипулировать предметами и применять их по назначению в тех случаях, когда их обучают этому в неволе. Несомненно, шимпанзе гораздо более “технологичны”, чем все остальные антропоиды. Означает ли это, что они и наиболее “интеллектуальны”? Ответ скорее отрицательный. Исследования когнитивных возможностей животных с помощью языков - посредников выявили практически одинаковый потенциал у шимпанзе, горилл и орангутанов, а бонобо были признаны самыми “интеллектуальными” приматами по результатам, достигнутым в целом комплексе тестов (Waal, 1995). По крайней мере потенциальные способности к орудийной деятельности позволяют включать соответствующие задания в тесты для исследования когнитивных способностей всех видов обезьян.
Использование орудий птицами. Устоявшиеся представления об орудийной деятельности птиц, базирующейся на инстинктивных, ”запаянных” программах поведения, в последние годы поколеблены веяниями когнитивной экологии. Один из самых интересных пример в этой области связан с новокаледонскими галками (Corvus moneduloides). Они достают насекомых из отверстий и трещин в коре деревьев с помощью преобразованных частей растений. Хант (Hunt, 1996, 2000), изучив около полутора тысяч птичьих орудий, разделил их на две группы: 1) “грабли”, изготовленные из прочных листьев Pandanus sp., у которых часть листовой пластинки срезана, а часть разъята на полоски; 2) “крючки”, или “лопатки”, сделанные из хвоинок, у которых конусовидно срезана часть листовой пластинки. Если орудие оказалось эффективным и сохранило свои ловчие качества после первого использования, птицы носят его в клюве и используют повторно в течение дня. Галки изготавливают свои орудия, следуя определенному “плану”, и стандартизация инструментов связана скорее с памятью и опытом, чем со строгой привязанностью к наследственной программе (Hunt, Gray, 2004). В то же время орудийное поведение C. moneduloides специализировано и тем отличается от гибкого поведения ворон, которые манипулируют предметами, нередко используя их также и в качестве орудий (Зорина и др., 1986). Наряду со специализацией разнообразие используемых материалов и строгая латерализация (галки оказались “правоклювыми”) выделяют новокаледонских галок среди животных, использующих орудия (Weir et al., 2004).
Галапагосский дятловый вьюрок (Cactospiza pallida) стал в последние годы излюбленным объектом исследований когнитивных этологов (Tebbich, Bshary, 2004). На орудийное поведение этого вида впервые обратил внимание Гиффорд (Gifford, 1919), детальные исследования осуществили Лэк (Lack, 1947) и Эйбл-Эйбесфельдт (Eibl - Eibesfeldt, 1961). Для того чтобы доставать насекомых из-под коры и из отверстий в стволах и ветках, вьюрок использует заостренную палочку. Птица не просто обламывает веточку, но и преобразует ее: укорачивает и заостряет, действуя клювом и ногами, а также обламывает мешающие отростки. Помимо веточек вьюрки используют лучинки, которые отщепляют от подходящих древесных поверхностей, а также колючки кактусов. Бауман (Bowman, 1961), сопоставив особенности диеты и морфологии разных видов вьюрков, предсказал и затем обнаружил орудийное поведение еще у двух видов галапагосских вьюрков, C. Heliobates и Certhidea olivacea . Исследование межпопуляционных различий в поведении дятлового вьюрка показало, что в аридных местообитаниях, где насекомые прячутся в укрытиях, большинство вьюрков используют орудия регулярно и таким способом добывают более половины пищевых единиц. В местах с влажным климатом насекомые доступны на поверхности стволов и листьев, случаи использования орудий редки (Tebbich et al., 2002).
Внимание к поведению птиц, возможно, поможет исследователям выявить орудийное поведение у широкого ряда видов. Так, поползень (Sitta pusilla) с помощью зажатого в клюве кусочка коры помогает себе приподнять кору сосны и достает оттуда насекомых. Птицы прибегают к такому способу питания в тех случаях, когда их обычная пища (семена сосны) становится редкой. Это наблюдалось только в локальной популяции поползня в Луизиане, США (Morse, 1968). Описано орудийное поведение белокрылой клушицы (Corcorax melanorhamphus), обитающей в Австралии и добывающей моллюсков Velesunio ambiguus. Для того чтобы вскрыть раковину, клушица подбирает осколки или целые створки вскрытых раковин и колотит ими по моллюску. Производительность труда клушицы составляет 18 раковин в 1 ч (Hobbs, 1971). Наблюдения за белоголовыми орланами (Haliaeetus leucocephalus) показали, что они бросают камни на черепах, тычут зажатой в клюве палочкой в панцирь черепахи, а также убивают скорпионов камнем, зажатым в когтях (Lawick-Goodall, Lawick, 1966). Вороны бросают камни и пучки травы в своих врагов, а также в сидящих на гнездах чаек, чтобы согнать их с гнезда и съесть яйца (Janes, 1976; Montevecchi, 1978). Известно, что вороны, грифы-бородачи и стервятники бросают с высоты на камни яйца, моллюсков и кости, чтобы расколоть их и достать содержимое. Певчий дрозд берет в клюв улитку и колотит ею по камню с той же целью (Morris, 1954). В этих случаях птицы используют камень как наковальню, и он не считается орудием, поскольку не является продолжением тела животного. Однако такое поведение, возможно, у некоторых видов эволюционно связано с использованием орудий. Египетские грифы (Neophron pernopterus) могут поднимать камень в воздух и бросать его на гнездо страуса, поднявшись в воздух, либо брать камень в клюв и бросать его на яйцо с высоты своего роста (Thouless et al.,1987).
Цапли некоторых видов бросают в воду разные предметы, привлекая тем самым внимание рыб, которых затем ловят. Впервые такое поведение было описано на примере зеленой цапли (Butorides virescens): птица использовала куски хлеба, бросая их на поверхность воды в том месте, где ею были замечены рыбы. Она приносила хлеб к месту, удобному для охоты, прогоняла других птиц, пытающихся схватить хлеб, и заменяла приманку на другую, если кусок отплывал от удобного места или тонул (Lovell, 1958). Позднее цапля того же вида в Морском аквариуме Майами была замечена за приманиванием рыб с помощью гранулированного рыбьего корма, который она воровала у сотрудников. Тем же занимались ее мать и брат, и птицы делали это достаточно часто; такое поведение можно рассматривать как обычное для их индивидуального репертуара (Sisson, 1974). В книге Бека (Beck, 1980) приводятся также единичные наблюдения разных авторов за другими видами (Ardeola ralloides, Eurypya helias), которые с той же целью использовали перья, личинки мясных мух и других насекомых. Детально была исследована орудийная деятельность в популяциях B. striatus :цапли использовали для ловли рыбы веточки, ягоды, насекомых, фантики, оброненные людьми крекеры. Было замечено, как цапля укорачивает веточку, прежде чем бросить в воду. Особи, прибегающие к орудийной технике рыбалки, редки в популяции, и среди них молодые цапли менее успешны, в частности, они используют крупные предметы, скорее пугающие, чем привлекающие, рыбу (Walsh et al., 1985). В том же ряду “приманивания добычи с помощью орудий” исследователи рассматривают поведение кроличьего сыча ( Athene cunicularia), которого, сравнивая с цаплями, они назвали “сухопутным рыболовом” (Levey et al., 2004). Сычи раскладывают вокруг своих вырытых в земле гнезд собранные ими экскременты млекопитающих и затем поедают привлеченных таким образом жуков-навозников. Опыты, в которых наблюдатели лишали хозяев гнезд их “орудий лова”, показали, что количество жуков, которые достаются сычам, уменьшается в среднем в 10 раз, если они лишены возможности использовать приманку для привлечения добычи.
Гнезда птиц не рассматриваются как “орудия для выращивания птенцов”. Однако при постройке славки-портнихи (род Orthotomus) используют орудия, помогающие скрепить элементы гнезда. Птички используют в качестве основы большой лист, свертывая лапками и клювом и затем закрепляя в форме кулька. Для того чтобы края листа не расходились, птица прокалывает клювом дырочки и протаскивает в них скрученные волокна паутины или стебельки трав. Пушистые кончики “нитей” сами по себе хорошо удерживаются маленькими отверстиями, но хозяйка гнезда к тому же сворачивает что-то вроде узелков; этот процесс ближе не к сшиванию, а к клепанию (Фройде, 1986, Attenborough, 1998).
Использование орудий беспозвоночными. Примеров орудийной деятельности беспозвоночных животных немного, что соответствует слабой изученности их поведения. Разрозненные примеры, относящиеся к ракообразным и головоногим моллюскам, представляются весьма спорными. Это связано не столько с “неспособностью” беспозвоночных, сколько с обширными лакунами в существующих данных. Систематические сведения об использовании предметов касаются парализующих ос, принадлежащих к родам Ammophila и Sphex. Самки выкапывают норки в сыпучем грунте, приносят туда парализованных насекомых как корм будущим личинкам, откладывают непосредственно на эти “живые консервы” яйца, запечатывают норку и улетают. Для уплотнения грунта при закрывании норки осы используют камешки, кусочки коры и веточки (Armbruster, 1921; Fricsh, 1940; Evans, Eberhard, 1970).
Пристальное внимание исследователей к поведению муравьев привело к тому, что и орудийная деятельность этих насекомых была замечена и оказалась предметом дискуссий. Первый вопрос, прозвучавший после доклада автора на Европейском экологическом конгрессе (Reznikova, 1995) о способности муравьев к сложным формам коммуникации, был следующим: “Если исследуемые вами виды муравьев способны к деятельности столь высокого уровня, не употребляют ли они орудия?” В обсуждении выяснилось, что председатель заседания энтомолог М. Р. Пайва наблюдала орудийное поведение муравьев Iridomyrmex purpireus , аналогичное тому, что открыли Феллерсы (Fellers I., Fellers G., 1976) в своем эксперименте на нескольких видах рода Aphaenogaster. Цитируемый опыт выглядел следующим образом. Возле гнезд муравьев выкладывали приманку в виде желе. Муравьи, обнаружив пищу, возвращались с кусочками листьев, помещали их в желе и уходили за новыми. В течение 30-60 мин они перекладывали кусочки по кормушке, а когда листья покрывались желе, уносили их в гнездо и там слизывали или соскребали пищу. Поведение выглядело весьма гибким, так как муравьи применяли для своей цели не только листья, но и кусочки сухой грязи и щепки. Природная ситуация, которая соответствует эксперименту, заключается в том, что муравьи часто встречаются с необходимостью транспортировки в гнездо жидкой пищи. Жидкая или желеобразная белковая пища служит кормом личинкам. Это может быть, например, поврежденное насекомое, с которого муравьи слизывают гемолимфу, перенося ее в зобике. Использование предметов позволяло муравьям переносить за один рейс почти в 10 раз больше пищи, чем при обычной транспортировке. По мнению авторов, такая “рационализация труда” позволяет муравьям компенсировать давление межвидовой конкуренции. Так, A. rudis , которых доминирующие виды прогоняют с добычи, значительно чаще прибегают к транспортировке жидкой пищи с помощью вспомогательных средств. Полевые эксперименты были с тем же результатом продублированы в лаборатории. Еще более ранняя публикация посвящена мурвьям-жнецам Pogonomyrmex badius (Morill, 1972). По мнению автора, жнецы не только используют, но даже изготовляют орудия: они формируют из песчинок гранулы и помещают их в мед, а затем транспортируют к себе в гнездо. В книге Б. Холльдоблера и Э. Вильсона “Муравьи” (Hölldobler, Wilson, 1990) эти результаты трактуются как следствие обычного для муравьев поведения “забрасывания” мешающих насекомым объектов различными доступными им мелкими предметами (веточками, частицами почвы). В тех случаях, когда речь идет о лужицах меда или пади, закиданных щепочками и комочками почвы, муравьи иногда относят в гнездо предметы, покрывшиеся слоем сладкого вещества. Можно, однако, полагать, что экспериментальное моделирование подобных ситуаций поможет более детально представить составляющие этой интересной формы поведения.
В целом можно предполагать, что склонность к использованию орудий является частью видовой специфики животных. Те виды, которые используют орудия в естественной среде обитания, демонстрируют при этом сложные комплексы поведения, включающие элементы индивидуального опыта, а в некоторых случаях, возможно, и социального распространения навыков. Наблюдение за использованием орудий дает исследователям дополнительную возможность интегральной оценки интеллектуальных возможностей животных и способностей к принятию решений.
ЭКСПЕРИМЕНТАЛЬНЫЕ ИССЛЕДОВАНИЯ КОГНИТИВНЫХ АСПЕКТОВ ОРУДИЙНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ
По организации последовательности поведенческих актов орудийная деятельность животных нередко выглядит проще, чем конструктивная. Комплекс действий, совершаемых иволгой, сплетающей гнездо-корзинку, или паучихой, подвешивающей на травинке сплетенный кокон, сложнее, чем действия обезьяны, колотящей камнем по ореху. Животные, сооружающие конструкции, действуют настойчиво и целенаправленно. Так, в гнезде королька насчитывается до12 000 стебельков мха, что говорит о том, что птичка тысячи раз повторяла действия с элементами конструкции, прежде чем завершила ее. Еще более впечатляют результаты коллективной конструктивной деятельности, например, величественные сооружения термитов или плотины бобров. Почему же именно орудийное, а не конструктивное поведение животных находится в центре внимания когнитивной этологии? В стремлении упростить ситуацию, можно было бы сказать, что конструирование у животных основано на инстинктивных действиях, тогда как орудийная деятельность опирается на когнитивные процессы. На самом деле такой четкой границы нет. Это отчасти видно из приведенных выше примеров, и будет показано ниже, на ряде экспериментальных результатов. Орудийная деятельность дает возможность интегральной оценки интеллекта животных, так как эта форма активности часто является своеобразной “вершиной айсберга”: ей предшествует последовательность действий, связанных с оценкой обстоятельств, поиском подходящих предметов, “расчетом” возможных последствий, коммуникацией с сородичами.
“Популярная физика” для приматов. Известный американский этолог Д. Повинелли назвал одну из книг “Популярная физика для обезьян” (Povinelly, 2000), отразив в названии характер одного из направлений когнитивной этологии, связанного с исследованием пределов компетентности животных в связях и закономерностях предметного мира. По замечанию Визальберги (Visalberghi, 2002), обезьяны часто изобретательно выполняют действия, направленные на решение задач, не имеющих смысла, и столь же часто совершают попытки выполнить что-то по-настоящему полезное, действуя совершенно бессмысленно. В качестве примера исследовательница приводит случай, который наблюдала в зоопарке: капуцин пытался расколоть земляной орех (который легко открыть зубами), колотя по нему вареной картофелиной.
Более 100 лет назад Хобхауз использовал в опытах с обезьянами экспериментальную методику, которая стала универсальной и положила начало многим модификациям (Hobhouse, 1901). Речь идет о задаче “выталкивания приманки из трубы”. В первом же опыте шимпанзе вставил палочку в небольшую трубку и пытался коснуться цели, а во втором он сумел вытолкнуть ее наружу из противоположного отверстия трубы. Йеркс (Yerkes, 1916) предлагал подобную задачу шимпанзе и горилле. Самка шимпанзе решила эту задачу после 12 дней экспериментов. Задача, по сравнению с опытом Хобхауза, осложнялась тем, что предложенная обезьяне труба не помещалась в руках животного, а лежала на земле и была очень длинной по сравнению с ростом обезьяны (170 см). Она, однако, оказалась способной сообразить, что приманку надо толкать в направлении “от себя” для того, чтобы добиться успеха. Горилла освоила этот прием, но только тогда, когда ее научил этому экспериментатор.
Г.З. Рогинский (1948) и Н.Н. Ладыгина-Котс (1959) исследовали орудийную деятельность шимпанзе, работая с шимпанзе Парисом, который жил в Московском зоопарке. Рогинский предлагал Парису классическую кёллеровскую задачу: притянуть к себе с помощью палки лакомство, находящееся на столе, вне его досягаемости. Освоившись с заданием, Парис обрабатывал орудия, не подходящие для достижения цели. Он расщеплял слишком толстую палку, а короткую заменял более длинной. В других опытах экспериментатор предлагал Парису лакомство в чашке, которую можно было притянуть к себе тесемкой, пропущенной через ручку чашки, соединив в руке свободные концы тесемки. Парис в первый же раз, сблизив обе тесемки в руке, достиг требуемого результата. Это произошло, по-видимому, случайно, так как впоследствии только после 30 ошибочных опытов он научился сближать и притягивать тесемки. Все же подлинного понимания соотношения между тесемкой и чашкой у обезьяны, по-видимому, не образовалось, так как в слегка видоизмененных условиях опытов Парис снова делал ошибки. Ладыгина-Котс исследовала орудийную деятельность Париса, предлагая ему задачу выталкивания приманки из трубки. Ее опыты отличались от опытов Хобхауза и Йеркса тем, что исследовательница ставила обезьяну в ситуацию выбора необходимого орудия, пригодного по форме и величине, из группы непригодных, а также исследовала возможность обработки или изготовления орудия при разрешении проблемы. Ладыгина-Котс придумала множество модификаций задания, которые дали возможность проанализировать поведение шимпанзе в ситуациях различной сложности. Ниже приводится примерно треть из списка предметов, которые предлагались Парису “к рассмотрению” для того, чтобы с их помощью вытолкнуть из трубки сверток с лакомством: ветвистый прутик, палка, завернутая в бумагу, палка, обмотанная веревкой, загнутый прутик, проволока с петлей на конце, с загибом, с двумя загибами, виток проволоки, одинарная и двойная петли из проволоки, проволока в виде восьмерки и к тому же скрепленная в двух местах, крест из проволоки, проволока с поперечиной, связка из пяти палочек (одна из которых нужной длины, а остальные короткие), деревянная палочка с пятью проволочными поперечинами (которые нужно вынуть, чтобы протолкнуть палочку в трубку), дощечка с расширением на конце (в другом варианте посредине), а также с двумя и тремя расширениями, трубки с втулками, подвижно скрепленные палки и т.п. Шимпанзе справился с большинством заданий, проявив недюжинную сообразительность и энергию: разгибал проволоку, вытаскивал поперечины, расширения дощечек отгрызал зубами, веревку разматывал. Он осуществлял деконструирование, деформирование, расчленение путем обработки и на разных этапах своей деятельности сравнивал размеры предметов, проверяя готовность орудия к действию. При этом, как и все шимпанзе, Парис делал неточно все, что допускало неточность: например, разгибал проволоку лишь до таких пределов, которые едва позволяли с трудом просунуть ее в отверстие.
Можно согласиться с Ладыгиной-Котс в том, что “шимпанзе – раб прошлых навыков, трудно и медленно перестраиваемых на новые пути решения”. Это иллюстрируется и результатами опытов с шимпанзе Рафаэлем, в которых исследователи также пытались подвигнуть обезьяну на “открытие” некоторых законов физики (Штодин, 1947; Вацуро, 1948). Архимеда из Рафаэля не получилось, хотя он и использовал собственные случайно найденные удачные приемы для того, чтобы предотвратить вытекание воды из дырявой кружки. В одном из опытов Рафаэлю дали для закладывания отверстия кружки металлический шарик. В первый раз он случайно заткнул им отверстие благодаря тому, что, положив шарик в рот, набрал в рот воды, которую вместе с шариком выплюнул в кружку, причем шарик попал в отверстие и закрыл его. Замечательно, что зверь все же установил связь между “затычкой” и вытекающей водой. Но замечательно и то, что впоследствии Рафаэль воспроизводил это случайно удачное решение в том виде, в каком оно осуществлялось в первый раз, т.е. вопреки надобности помещал шарик, взятый рукой, в рот, а изо рта выплевывал его в кружку, в которую наливал воду. Получив целую кружку, Рафаэль опускал шарик и в нее, а когда ему дали две кружки, целую и продырявленную, он предпочел последнюю, повторяя ставшие для него ритуальными действия с шариком.
Л.А. Фирсов (1977) предложил членам колонии шимпанзе, живущих летом на острове, задачу, которую можно назвать “обратной задачей с трубкой”. Четырем обезьянам предлагалось достать приманку из выкопанной в земле узкой вертикальной ямки, глубиной 80 см, что было немного больше, чем дина руки шимпанзе. Первые действия всех обезьян были примитивны: они пытались достать приманку поочередно каждой из четырех конечностей. Отдышавшись, обезьяны все это повторяли еще раз, уже с меньшим энтузиазмом, а затем начинали оглядываться по сторонам. Покинув на некоторое время соблазнительную приманку, шимпанзе подбирали палки и доводили их до рабочего состояния с помощью зубов и рук. Интересно отметить, что после первого приступа бурной деятельности с орудием все обезьяны проходили стадию “спокойного обдумывания”, усаживаясь или укладываясь рядом с ямкой и затем переходя к спокойной “осаде” приманки.
Простые варианты обычной “задачи с трубкой” использовались в лабораторных условиях и применительно к другим видам обезьян: гамадрилам, мандрилам, макакам-резусам, макакам-лапундерам, мангобеям и капуцинам (Watson, 1908; Klhver, 1933; Harlow, 1951; Ладыгина-Котс, 1959). В ранних экспериментах лишь капуцины проявили себя как перспективные кандидаты, способные претендовать на решение задачи, однако полностью она ни разу не была ими решена. Визальберги с соавторами (Visalberghi, 1990; Visalberghi et al., 1995) использовали усложненные варианты задачи с трубкой, взятые почти без изменений из книги Ладыгиной-Котс. Обезьяны должны были вытолкнуть лакомство из трубки с помощью палочки, но в первом варианте палочки были связаны вместе, так что для достижения цели их надо было разъединить, во втором в палочку были вставлены поперечные штырьки, мешающие проталкиванию ее в трубку, в третьем требовалось вставить одну палочку в другую. Капуцины справились со всеми предложенными задачами, сочтя первую из них более легкой, чем остальные. Однако они перебирали и совсем неподходящие варианты, например засовывали одну короткую палку в один конец трубки с приманкой, а другую в другой. Более того, в повторных опытах число ошибок существенно не уменьшалось. Капуцины в отличие от шимпанзе, видимо, не слишком хорошо понимают связи между элементами задачи. В этой серии опытов участвовали и человекообразные. Оказалось, что юные шимпанзе (2-4 года) мало отличаются по “успеваемости” от капуцинов, особенно при решении второй и третьей задач. Взрослые обезьяны (4 бонобо, 5 шимпанзе и 1 орангутан) показали значительно лучшие результаты, но только при решении первого варианта. Все они сразу разъединяли палочки и не делали попыток достать приманку слишком коротким предметом. Второй и третий варианты задачи оказались для человекообразных так же трудны, как и для капуцинов, а существенной разницы в подходах к решению не наблюдалось. Те же задачи были предложены другим группам обезьян, и нашелся один капуцин, ни в чем не уступающий шимпанзе, а те в свою очередь не справились с заданием с поперечными штырьками (Anderson, 1996). Это говорит о существенном размахе индивидуальных вариаций в решении инструментальных задач в пределах каждого вида.
Остроумная модификации задачи с трубкой – “трубка с ловушкой” – оказалась сложной для всех исследованных видов обезьян (Visalberghi , Trinca, 1989). В этих опытах трубка имела отверстие, которое открывалось в прикрепленный снизу стаканчик. Если приманку толкать по направлению к стаканчику (“ловушке”), она упадет туда и не достанется обезьяне. Значит, надо толкать в противоположном направлении. Это умозаключение удалось только одному из четырех испытуемых капуцинов. Действия этой самки казались планируемыми, т. е. она строила свое поведение так, чтобы избежать “наказания” со стороны ловушки. В половине случаев она толкала приманку в неверном направлении, но, увидев, что лакомство вот-вот упадет в стакан, вытаскивала орудие и бежала в другому концу установки, меняя ошибочные действия на правильные. Однако число ошибок не уменьшалось день ото дня с ходом испытаний. Экспериментаторы “добили” животное тем, что перевернули установку так, что ловушка оказалась сверху (задача получила название “нефункциональная ловушка”). К этому времени обезьяна запомнила, что успешными являются действия, направляющие приманку в сторону, противоположную ловушке. Однако если ловушка сверху, то она уже не работает и не “отнимает” приманку, поэтому нет смысла следить за направлением движения орудия. Выучив “правильное” движение, обезьяна продолжала на нем настаивать. Исследователи объяснили это тем, что капуцины, по-видимому, не очень сильны в понимании причинных связей и им не удается планирование относительно сложных действий (Visalberghi et al., 1995). Задачу “трубка с ловушкой внизу” предложили пяти шимпанзе (Limongelli et al., 1995). В течение 70 первых опытов их действия были случайными. Два шимпанзе в течение последующих 70 опытов научились избегать “наказания” со стороны ловушки. Авторы предложили этим двум особям другие варианты положения ловушки: не в середине, как в первом варианте, а у самого конца трубки. Обезьяны успешно справились с этой задачей без всякого доучивания, что привело исследователей к предположению о том, что они неплохо понимают условие задачи. Тем не менее, когда одному шимпанзе из числа тех, что наиболее успешно решали задачу, была предложена “нефункциональная ловушка”, то, несмотря на многообещающее поведение на предварительной стадии экспериментов, этот шимпанзе действовал так же, как и капуцин, то есть настаивал на предварительно выученной последовательности движений (Reaux et al., 1999).
Степень проникновения в предметную ситуацию у разных видов обезьян была исследована и с помощью тестов на способность к абстрагированию в ходе решения инструментальной задачи. В контексте исследований речь шла о способности животных к выделению среди свойств предметов, изменяющихся в эксперименте, таких ключевых признаков, которые имели отношение к решению задачи. Обезьянки тамарины были предварительно тренированы доставать белую чашку с лакомством с помощью голубой загнутой на конце тросточки. В последующих опытах им предлагали на выбор предметы разных цветов и разной формы, часть из которых подходила для решения задачи, а часть нет (были слишком мягкими, слишком короткими или широкими и т.п.). Варьировали и цвета чашек с кормом. Тамарины прекрасно ориентировались в физических свойствах предметов, игнорируя меняющиеся второстепенные признаки. Они не обращали внимания на изменение текстуры предметов и их цвета, но сразу реагировали на изменение функциональных признаков, т. е. формы и размеров (Hauser et al., 1999). Это тем более удивительно, что речь идет о виде, который в естественных условиях не прибегает к использованию орудий.
В целом применение орудийных методик к исследованию интеллекта обезьян показало, что представители различных семейств приматов не только успешно манипулируют предметами для достижения цели, но и имеют некоторые, хотя и ограниченные, представления о связи форм и размеров предметов с их свойствами. Индивидуальные различия, проявляющиеся при решении инструментальных задач, очень велики. Становится понятным, почему, если 50 лет назад исследователи единодушно проводили границу между ментальными возможностями “высших” и “низших” (см.Miyamoto et al., 1988) обезьян на основе их достижений в при решении инструментальных задач, то сейчас к этому вопросу подходят более осторожно, стараясь сравнивать большое число особей и множество вариантов заданий. На данном этапе исследований разница в когнитивных возможностях между человекообразными обезьянами и остальными видами проявляется скорее на количественном, чем на качественном уровне (Call, 2000).
“Популярная физика” для птиц. Первые представления об интеллектуальных возможностях птиц, связанных с орудийным поведением, восходят к басне Эзопа о вороне, который поднимал уровень воды в кувшине, кидая туда камешки. Наблюдал ли кто-нибудь такую ситуацию на самом деле, неизвестно, однако если это и вымысел, то он, несомненно, основан на реальных случаях, в которых птицы проявляли способность к улавливанию свойств вещей и к их практическому использованию. В книге Бека (Beck, 1980) собраны примеры наблюдений за ситуациями, в которых птицы манипулировали жидкостями и емкостями, используя различные их свойства (в частности, вязкость жидкостей). Так, какаду использовал половинку от скорлупы грецкого ореха, чтобы добыть остатки воды со дна поилки; серого жако наблюдали всасывающим воду через соломинку, а попугая кеа – за игрой, которая заключалась в вычерпывании воды чашкой из больших емкостей; содержащийся в неволе ворон Corvus brachyrhyncos регулярно набирал пластиковым стаканчиком воду из поилки, относил к кормушке, наливал в еду и ждал, пока она размокнет и станет более аппетитной (Beck, 1980).
Первое детальное описание орудийной деятельности птиц, основанной на понимании ими свойств предметов, сделано на примере голубых соек Cyanocitta cristata (Jones, Kamil, 1973). Экспериментаторы обратили внимание на то, что, проголодавшись, одна из птиц отрывала кусок газеты, подстеленной под ее клетку, уносила на жердочку, затем, прижимая бумагу ногами, сворачивала в несколько раз с помощью клюва, возвращалась к решетке, просовывала изготовленный жгут сквозь отверстия и подгребала к себе кусочки пищи, находящиеся слишком далеко, чтобы достать их без помощи орудия. Инструментальное поведение было ответом на плановые ограничения в пище, которые соответствовали требованиям контроля над здоровьем птицы. Столь изобретательную форму протеста исследователи положили в основу специального эксперимента. Сойку стали снабжать различными материалами, подходящими для изготовления орудий, оставляя при этом гранулы корма на соответствующем расстоянии от клетки. Оказалось, что сойка манипулирует предметами тем чаще, чем дольше ее продержали без пищи. Ее орудийное поведение было весьма гибким: помимо газеты в ход шли пучки соломы, стебли чертополоха, куски пластиковых лент, которые используют для упаковки. Все эти предметы ловко преобразовывались для удобства манипулирования. Другие члены лабораторной колонии также были испытаны в сходных условиях. Из восьми птиц пять демонстрировали выраженное орудийное поведение, две - только фрагменты его, и лишь одна совсем не проявила соответствующей активности. Поскольку птицы содержались вместе, авторы предполагают существенную роль подражания в формировании орудийного поведения, дополненную самообучением. Это предположение подкреплено наблюдениями за членами локальной популяции соек, использующих орудия в естественных условиях (см. ниже).
Систематические исследования интеллектуальных аспектов орудийной деятельности птиц в настоящее время проводятся в лаборатории поведенческой экологии в Кембридже с новокаледонскими галками (Chappell, Kacelnik, 2002). Для того чтобы достать лакомство, птицы используют широкий набор предметов: собственные перья, выпавшие при линьке, полоски картона, обрезки проволоки. Предметы подвергаются преобразованию: птицы счищают с перьев бороздки, картон расчленяют на полоски требуемых размеров, проволочки сгибают так, что получаются крючки. Такой творческий подход галок к манипулированию предметами подсказал исследователям идею экспериментов, в которых птицы, подобно приматам, учитывали в своей деятельности закономерности механики. Прежде всего была исследована способность птиц выбирать среди разнообразных предметов те, которые в наибольшей степени подходили к решению предложенной задачи. В первой серии опытов птицам предлагали палочки разной длины, с помощью которых они могли вытолкнуть из прозрачной трубки кормушку с лакомством. Если палочки лежали рядом с установкой, обе птицы успешно соразмеряли длину выбираемого орудия и видимое расстояние до кормушки. Во второй серии палочки и экспериментальный аппарат находились в разных помещениях, так что нужно было, выбирая орудие, “мысленно” примерять его не к видимой, а к удерживаемой в памяти цели. С этим заданием достаточно успешно справлялся только самец Абель. В следующей серии опытов проверялась способность птиц модифицировать незнакомые им предметы для достижения цели, т.е. изготовлять орудие, адекватное ситуации (Weir et al., 2004). Здесь лидировала самка Бетти. Кусочки мяса помещали в корзинку, находящуюся внутри полой трубки, а для манипуляций галкам предлагались прямые проволочки. Раньше птицы с подобным материалом не встречались. Обе они пытались достать лакомство с помощью прямых проволочек. Абель достал приманку в одном случае из 10, но не пытался модифицировать незнакомый ему материал. Бетти пыталась достать приманку прямой проволочкой, но уже спустя полминуты приступала к изготовлению крючка. Она заклинивала один конец проволоки, помещая его под прочную опору, которая находилась на расстоянии до 3 м от аппарата. Другой конец галка загибала клювом. Изготовив крючок, она направлялась к экспериментальной установке, и доставала мясо. Полученные данные тем более удивительны, что в подобной ситуации шимпанзе показали гораздо худшие результаты.
Например, получая в виде заготовки согнутую трубку, они не догадывались разогнуть ее, протолкнуть в отверстие и достать яблоко (Povinelly, 2000). А ведь обезьяны в отличие от птиц имели предварительный опыт манипуляций с предлагаемыми заготовками.
Птицы участвовали также в сериях экспериментов “трубка с ловушкой”, с использованием метода, опробованного на приматах. Среди новокаледонских галок и дятловых вьюрков выделялись особи, которые столь же легко, как и обезьяны, выучивали последовательность действий, ведущую к успеху (Chappell, Kacelnik, 2004; Tebbich, Bshary, 2004). Наиболее успешному вьюрку был предложен вариант задачи “нефункциональная ловушка”. Птичка действовала точно так же, как и капуцин в опытах Визальберги, настаивая на выученной последовательности действий (Tebbich, Bshary, 2004).
В целом представители некоторых видов приматов и птиц при решении инструментальных задач демонстрируют значительную компетентность в закономерностях и связях физического мира. Представляет существенный интерес вопрос о том, реализуется ли столь высокий когнитивный потенциал у исследованных видов в ситуациях, не требующих использования орудий, или это узко направленная видовая “гениальность”. Мы проводим здесь аналогию с животными, запасающими пищу и запоминающими местоположение тысяч своих кладовых, таких специалисты называют “гениями запоминания” (Shettleworth,1990). Некоторые виды муравьев и медоносные пчелы могут быть названы “гениями коммуникации” (Резникова, Рябко, 1990). При этом (и это важно отметить), упомянутые животные демонстрируют вполне заурядные способности при решении целого ряда иных задач. Новокаледонских галок, вьюрков и шимпанзе в силу их видовой специфики можно было бы назвать “гениями орудийной деятельности”. В экспериментах животные демонстрируют накопление индивидуального опыта путем проб и ошибок, и при этом формирование оптимальных решений происходит быстро и целенаправленно. По-видимому, во всех описанных ситуациях речь идет о поведении, основанном на видотипической склонности к образованию специфических ассоциаций.
СООТНОШЕНИЕ ВРОЖДЕННЫХ И ПРИОБРЕТЕННЫХ КОМПОНЕНТ В ФОРМИРОВАНИИ ОРУДИЙНОГО ПОВЕДЕНИЯ
Вопрос о соотношении различных компонент в поведении животных – один из самых волнующих разделов этологии, а исследование соответствующих аспектов орудийной деятельности – один из наиболее интересных примеров в этом разделе. Участвуют ли когнитивные процессы в онтогенетическом формировании орудийной деятельности ?
Влияние раннего опыта и груза стереотипов на решение инструментальных задач. Существенное положительное значение для успеха в решении инструментальных задач имеет индивидуальный опыт животных, ранее приобретенный в играх с предметами и при добывании пищи с их помощью. Это фиксировали многие исследователи со времени ранних опытов до экспериментальных работ последних лет. В естественной ситуации детеныши человекообразных обезьян осваивают манипуляции с предметами во время игры в младенческом возрасте, во время тесной связи с матерью. Годовалые шимпанзе сгибают прутики, как бы строя гнездо, они также играют палочками, “добывая” муравьев из самых неподходящих мест, например, из шерсти матери. В трехлетнем возрасте они начинают живо интересоваться камнями и манипулировать ими, хотя пройдет еще немало времени, прежде чем обезьяны перейдут к настоящей орудийной деятельности. Игры с предметами особенно характерны для подростков (Гудолл, 1992). Социальная и зрительная депривация в течение первых 2 лет жизни вызывает нарушение когнитивных функций у шимпанзе, которые уже не восполняются в более зрелом возрасте. Это было показано в эксперименте, в котором сравнивали животных, достигших 14 лет и с двухлетнего возраста содержащихся в одинаковых лабораторных условиях, а до двух лет – в разных. Те из них, которые содержались в обедненной сенсорной среде, значительно хуже справлялись со всеми задачами, включая инструментальные, чем нормально развивающиеся животные (Rumbaugh, 1974). Эти результаты подчеркивают стимулирующую роль естественного раннего опыта животных в развитии у них сложного, в том числе орудийного, поведения. Напротив, явно тормозящую роль в проявлении когнитивной стороны орудийной активности у обезьян играет довлеющий над ними груз приобретенных стереотипов. У шимпанзе они легко превращаются в ритуалы, которые в меняющихся обстоятельствах становятся бессмысленными и даже мешающими. По свидетельству многих приматологов, обезьяны всех видов склонны настаивать на повторении собственных действий, которые однажды привели их к успеху. Они с гораздо большей легкостью изобретают новые формы поведения, применительно к возникающим в их жизни задачам и ситуациям, чем переучиваются или переносят навыки из одной ситуации в другую. Примером могут служить шимпанзе из лаборатории Павлова, Рафаэль и Роза, которые тушили огонь, перебираясь за водой по шесту на соседний плот и “не догадываясь “ зачерпнуть ее кружкой из озера. Полевые исследователи приводят немало подобных примеров, касающихся орудийной деятельности. Например, добывая плоды в кроне дерева, шимпанзе, не имея возможности вскарабкаться по гладкому стволу фигового дерева, забирались на соседнее, отламывали с него ветки, очищали и, действуя как крючьями, притягивали к себе ветви с плодами. Эта деятельность стала ритуальной для группы самцов. Им становилось все труднее добывать плоды, так как все подходящие ветки на доступном дереве оказались обломанными. Ни разу не было замечено, чтобы шимпанзе принес палку, взобрался на удобное дерево и использовал ее в качестве орудия, хотя, например, при добывании насекомых обезьяны поступают именно так: приносят подходящую палку со значительного расстояния (Sugiyama, Koman, 1979).
В новых для себя обстоятельствах обезьяны часто демонстрируют орудийную изобретательность, поражающую воображение. Примеры щедро рассыпаны по книгам, посвященным экспериментальному изучению обезьян и наблюдениям за ними. Например, в книге Л.А.Фирсова (1977), посвященной описанию жизни островной колонии шимпанзе, приводится много примеров подобной изобретательности, которой особенно отличался шимпанзе-подросток Тарас. Он заклинивал дверцу экспериментального аппарата сучком для того, чтобы без хлопот достать приманку, поднимал с помощью длинной палки затонувшую часть веревки, которой была привязана к берегу лодка, чтобы, не замочив ног, подтянуть ее к себе и полакомиться привезенными припасами. Не находясь в плену выученных стереотипов, шимпанзе могут эффективно и быстро достигать множества целей, манипулируя с предметами.
В описанных выше экспериментах с вьюрками был достигнут результат, который позволяет предположить, что в условиях, когда груз выученных стереотипов не довлеет над поведением, птицы потенциально способны к использованию представлений о действующих физических силах. Дело в том, что, когда вьюрку, настаивавшему на повторении стереотипных, но ставших бессмысленными действий при решении задачи с трубкой (вариант “нефункциональная ловушка”), предложили видоизмененную задачу, в которой стеклянную трубку заменили непрозрачной, характер деятельности птички резко изменился, и число успешных решений быстро достигло 90% (Tebbich, Bshary, 2004). Не исключено, что наследственная предрасположенность к оперативному формированию специфических связей препятствует проявлению инновационного поведения. Можно предположить, что, если бы вьюрки, шимпанзе и другие животные не обучались “слишком быстро”, они были бы способны к решению более широкого класса задач, требующих применения когнитивных способностей.
Развитие орудийного поведения в онтогенезе. Известно, что у разных видов животных наблюдается целая гамма проявлений сложных форм видотипического поведения – от полностью запрограммированной последовательности действий до полностью выученной. Хрестоматийным стал пример с видовыми песнями птиц (Marler, 1970). В группах близких в систематическом отношении видов можно найти разные варианты формирования песенного поведения: полностью запрограммированное и проявляющееся вне зависимости от действующих стимулов; врожденное, но зависящее от наличия образца, услышанного во время чувствительных периодов в онтогенезе; выученное на основе врожденной склонности к формированию определенной песни (“песенной болванки”); наконец- полностью сформированное подражанием образцу. Похоже, что такой же ряд можно построить и по отношению к орудийному поведению птиц. Голубые сойки Cyanocitta crystata обучаются использованию орудий почти исключительно социальным путем, подражая другим особям своего вида. Подобно вьюркам они используют острые предметы для извлечения насекомых из-под коры. Такое поведение известно лишь для одной популяции (Jones, Kamil, 1973). Этот пример, хотя и не столь яркий и известный, находится в одном ряду с культурным распространением навыков в популяции английских синиц (Fisher, Hinde,1949). Противоположным примером этого ряда является поведение египетского грифа Neophron pernopterus , который раскалывает для еды яйца страусов и других птиц, бросая на них камни. Элкок (Alcock, 1972) предположил, что такое поведение грифов эволюционировало из стереотипа раскалывания о камни яиц, бросаемых сверху, то есть в основе орудийного стереотипа лежало неорудийное манипуляторное поведение. Эта гипотеза получила экспериментальную поддержку (Thouless et al., 1989). Грифы, воспитанные в неволе и не знакомые ни с яйцами, ни с камнями, манипулировали камнями, раскалывая ими яйца, столь же успешно, как и их сородичи в дикой популяции. Интересно отметить, что если действия с камнями проявлялись у птицы спонтанно, то сведения о том, что яйца можно есть, добывались с помощью индивидуального опыта. Молодые грифы, как оказалось, не реагируют на них как на пищевые объекты до тех пор, пока не попробуют яйца, расколотые другими птицами или поврежденные случайно. Манипулирование камнями выглядит даже у взрослых грифов довольно неуклюже, и им удается расколотить не более половины тех яиц, к которым они подступаются с камнем (Lawick-Goodall, Lawick, 1966). Предположение Элкока о том, что предки Neophron pernopterus бросали яйца на камни, а не камни на яйца, подтверждается наблюдениями за выбором подходящих объектов у современных грифов. Птицы выбирают камни, похожие на яйца по форме и размеру, хотя это необязательно для удачного броска (скорее наоборот). Найдя яйца, птицы удаляются порой на 100 м, чтобы отыскать подходящий камень. Когда им предлагали предметы разной формы и разного цвета в качестве цели, оказалось, что стимулом к действиям является округлая форма в сочетании с размером. Грифы пытались расколоть зеленые и красные круглые предметы, но проходили мимо светлого куба, хотя он и был такого же цвета, как страусиное яйцо. Грифы проявляют строгую избирательность по отношению к размеру и весу орудия: среди предложенных камней они выбирали экземпляры, вес которых в узких пределах колебался около 46 г, хотя такая точность совсем необязательна для эффективного воздействия на яйцо. Столь же строгая избирательность и сходные параметры поведения были описаны позднее для обитающего в Австралии канюкового коршуна (Hamirostra melanosternon). Птицы этого вида выбирают для раскалывания яиц камни весом около 40 г (Aumann, 1990). Вероятным представляется предположение о видоспецифичных стереотипах орудийного поведения, которые носят врожденный характер.
В центре данного ряда вариантов соотношения запрограммированного и выученного поведения может быть помещено поведение дятлового вьюрка. В 60-е годы были проведены наблюдения, подчеркивающие роль приобретенного опыта (Eibl-Eibesfeldt, 1961; Millikan, Bowman,1967). Птенцы вначале действуют только клювом и лишь постепенно приходят к использованию орудий. Но каким путем они приходят к этому? Имеет ли место созревание нервных структур и соответственно проявление врожденных стереотипов, или молодые птицы подражают взрослым? Исследователи провели эксперимент, поместив в соседние вольеры дятлового вьюрка и большого кактусового земляного вьюрка (этот вид не использует орудия). Наблюдая за тем, как действуют соседи, земляные вьюрки стали использовать палочки, чтобы доставать насекомых. Это говорит о возможной роли подражания в орудийной деятельности вьюрков. Однако вьюрки других видов в подобных экспериментах не изменили свое поведение. Это означает, что подражание, если оно имеет место, базируется на наследственной предрасположенности к определенному роду деятельности.
В последние годы появилась серия экспериментов, основанных на вновь выявленных экологических особенностях фуражировочного поведения вьюрков (Tebbich et al., 2001). Выше упоминалось о том, что вьюрки, обитающие во влажных местах с обилием пищи, могут просто собирать насекомых с листьев и стволов. Среди них есть особи, изредка употребляющие орудия, но большинство птиц вообще ими не пользуется. Представители того же вида, обитающие в сухих и жарких местах, прибегают к использованию палочек и колючек. Авторы использовали природный эксперимент для ответа на следующие вопросы: могут ли вьюрки, которые ранее не использовали палочки, научиться этому путем подражания “умелым” сородичам ? как формируется орудийное поведение у вьюрков в условиях дефицита пищи: за счет подражания взрослым или путем накопления и совершенствования навыков? На первом этапе исследовались индивидуальные различия в поведении “неумелых” вьюрков, отловленных в местах с избытком пищи. 28 птичек тестировали дважды в день. Каждая особь находилась в помещении, в котором были созданы условия для добывания пищи: деревянный ствол с запрятанной в нем личинкой и палочки. В отдельных опытах птиц вынуждали доставать пищу из стеклянного “дупла”, так что можно было наблюдать и контролировать весь процесс проявления данной формы поведения. На втором этапе опытов 10 птичек, которые ни разу не употребляли орудия, помещали в одну вольеру с 10 “умелыми” демонстраторами. Спустя 2 нед. после совместного пребывания с демонстраторами, птичек вновь тестировали индивидуально, проверяя, научились ли они пользоваться орудиями. Наконец, в отдельных опытах исследовали выводки, взятые вместе с гнездом и родителями из тех и других местообитаний. Дятловые вьюрки строят закрытые гнезда, поэтому в природе птенцы не могли наблюдать за деятельностью родителей. Выводки заранее помещали в вольеры, а затем слетков из каждого семейства делили на две группы и воспитывали в разных условиях, так что получались следующие сочетания: птенцы умелых родителей в обществе птиц, не употребляющих и употребляющих орудия и наоборот (всего 4 группы). Оказалось, что птенцы следят за действиями неродственных птиц даже более охотно, чем за собственными родителями. В первой серии опытов в течение 5 нед испытаний орудийное поведение проявилось у 13 птиц, остальные ни разу не взяли в клюв палочку. Во второй серии из группы взрослых отобрали “неумелых” птиц и поместили вместе с “умелыми” демонстраторами, пойманными в засушливых местообитаниях, где орудийная деятельность была для них обычной. Такие птицы применяют палочки в среднем 6 раз за получасовое испытание. Группа “неумелых” наблюдателей была не совсем однородной: в ней было 7 птиц, которые ни разу не манипулировали палочками, и 3 особи, проявляющие интерес к орудийным заготовкам и манипулирующие ими хотя бы изредка. Из этих трех один вьюрок обучился путем наблюдений за демонстраторами употреблять орудие для доставания добычи. Остальные 9 из 10 вьюрков никак не изменили своего отношения к палочкам. Что же касается птенцов, то оказалось, что социальный опыт никак не влияет на формирование у них орудийного поведения. Будучи воспитанными в течение 5 нед в условиях, когда пища периодически была недоступна без вспомогательных предметов для голодных молодых птиц, все 13 вьюрков активно манипулировали палочками вне зависимости от того, могли ли они наблюдать за аналогичными действиями взрослых птиц или нет. Исследователи выделили 5 стадий в развитии игрового орудийного поведения вьюрков и при этом не выявили никаких существенных различий между группами. У всех птенцов одинаково развивалась достаточно высокая манипуляторная активность.
Итак, можно говорить о сложной форме поведения, которая формируется на наследственной основе путем индивидуального совершенствования. Значительную роль играет наличие чувствительного периода в онтогенезе птиц, т.е. возможность “оттачивать” соответствующую форму поведения в раннем возрасте. Если вьюрков лишить этой возможности, часть необходимых элементов орудийного поведения утрачивается. У птиц, обитающих в условиях избытка открытой и доступной добычи, происходит, по-видимому, утрата элементов юношеского игрового орудийного поведения, и они к этой форме манипуляторной активности уже не возвращаются. Остается еще много нерешенных вопросов. Выше цитировались опыты, в которых было показано, что взрослые кактусовые земляные вьюрки обучаются путем подражания дятловым вьюркам, однако сами дятловые вьюрки практически не подражают конспецификам. Можно предположить, что некая “заготовка” данной формы поведения присутствует у одних видов и отсутствует у других и при этом для ее реализации в онтогенезе разным видам требуются разные условия.
Роль наследственно закрепленных “заготовок” поведения оказалась значительной и для новокаледонских галок, пожалуй, самых изобретательных пользователей орудий в животном мире. Это показано в недавних экспериментах, в которых птенцы галок воспитывались в лаборатории с самого раннего возраста (Kenward et al., 2005). Четырех птенцов разделили: двух воспитывали вместе друг с другом, им регулярно давали манипулировать палочками и люди-воспитатели показывали, как с помощью орудий добывать насекомых из щелей. Два других птенца воспитывались поодиночке и никогда не видели орудий. Хотя птенцы первой группы очень внимательно наблюдали за деятельностью демонстраторов и пытались им подражать, по мере созревания птиц не было отмечено никаких различий в уровне их манипуляторной активности. Все они одинаково эффективно использовали палочки и начинали это делать примерно в одинаковом возрасте (63-79 дней). В другой серии опытов всем птицам предъявили листья растения Pandanus, которые в естественных условиях служат галкам источником для изготовления орудий. Только одна галка, причем именно та, что была воспитана в изоляции, с первого испытания применила эффективную последовательность действий для изготовления “грабель” и сразу добыла личинку насекомого из щели. Впоследствии она неоднократно повторяла успешные действия, которые проявились у нее по принципу “все и сразу”. Остальные галки атаковали листья, однако им потребовалась длительная достройка врожденного стереотипа, а эффективного изготовления орудий из листьев они в ходе опытов так и не достигли.
Исследование орудийной деятельности птиц позволило пролить свет на одну из самых интересных проблем этологии, а именно, на проблему роли врожденных компонент в формировании сложных поведенческих стереотипов. Нами были получены весьма сходные результаты при исследовании охотничьего поведения муравьев - факультативных охотников на прыгающую добычу. Стереотип поимки жертвы у небольшой части семьи (менее 10%) формировался по принципу “все и сразу”, остальным особям требовалась многоэтапная достройка врожденного поведения (Фабри, 1976), включающая простейшие формы социального обучения (Резникова, Пантелеева, 2001). Это, хотя на первый взгляд далекое сравнение, как нам кажется, помогает раскрыть вопрос об относительном вкладе разных форм поведения в формировании сложных видотипических стереотипов. Как у муравьев, так и у новокаледонских галок проявление стереотипа по принципу “все и сразу” хотя бы у одной особи дает основание предполагать наличие генетически зафиксированного поведенческого стереотипа (и это говорит о строгой необходимости множественного сравнения индивидуальных онтогенетических сценариев). При этом распределение готовых фрагментов поведения среди муравьев оказалось близко к тому, что продемонстрировано в процитированном выше примере с сойками (Jones, Kamil, 1973), а не к тому, что позже наблюдалось у новокаледонских галок (Kenward et al., 2005). Характер распределения поведенческих моделей в экспериментальных группах можно объяснить тем, что как для муравьев, так и для соек выявленные сложные стереотипы поведения носят факультативный характер и в естественных условиях проявляются лишь в локальных популяциях. Как выяснилось в экспериментах (Jones, Kamil, 1973; Резникова, Пантелеева, 2003), в поведенческом репертуаре и тех и других присутствуют готовые “блоки”, которые высвобождаются при наличии соответствующих раздражителей и мотиваций. Новокаледонские галки - специализированный по отношению к орудийной деятельности вид. К настоящему времени это единственный известный среди птиц вид, у которого не часть особей в популяции (как, например, у вьюрков) и не локальные популяции (как у соек), а все члены всех популяций (которые удавалось наблюдать) используют орудия (Kacelnik et al., 2004). Можно полагать, что поэтому и наследственно обусловленные “заготовки” сложнейшего поведенческого стереотипа у них представлены более полно, чем у видов, факультативно использующих орудия. Тем не менее, у части особей в популяции сложный поведенческий стереотип требует многоэтапной достройки. Можно предсказать выявление в недалеком будущем существенной роли социального обучения для синхронизации данного поведения на популяционном уровне. Возможно, что обсуждаемые закономерности являются общими не только для орудийной деятельности, но и для многих сложно организованных видотипических форм поведения.
Весьма сложные вопросы, касающиеся формирования орудийной деятельности в онтогенезе, задают своим исследователям и приматы. Показательна серия опытов на тамаринах. Описанные выше эксперименты с выбором подходящих орудий, в которых от обезьян требовалось среди меняющихся свойств предметов (текстура, цвет, размер, форма) выбирать ключевые признаки, необходимые для решения задачи, были проведены не только на взрослых обезьянах, но и на детенышах, лишь недавно вышедших из-под материнской опеки (Kralik, Hauser, 2002). Как уже отмечалось, представители данного вида не употребляют орудий на воле. Оказалось, что детеныши, несмотря на свой юный возраст и полное отсутствие опыта, легко ориентировались в меняющихся свойствах предметов, выбирая признаки, необходимые для удачных манипуляций, практически не уступая в этом взрослым. Это позволило авторам предположить, что в мозгу общего предка приматов были заложены структуры, создающие предпосылки для интенсивной орудийной деятельности.
Роль импринтинга в формировании сложных форм поведения, в частности орудийной деятельности, представляется недостаточно исследованной в онтогенезе приматов. Между тем, на этом пути возможны интересные открытия. Так, весьма интересно свидетельство Л.А.Фирсова (1977) о том, что в его островной колонии шимпанзе только две самки прибегали к ужению муравьев и разорению гнезд земляных ос с помощью изготовляемых на месте орудий (заостренных прутиков). Эти же обезьяны делали себе добротные гнезда на деревьях, хотя употребляемая ими для этой цели ольха, конечно же, существенно отличалась от деревьев в их родных джунглях. Остальные строили довольно жалкие подобия гнезд, а к муравьям и осам не подступались, хотя и имели возможность часто наблюдать за успешной деятельностью своих сородичей. Автор называет наблюдаемые формы поведения “джунглевыми стереотипами”. Действительно, “умелые ” самки попали в лабораторию из джунглей, тогда как остальные члены колонии были взяты в лабораторию в младенческом возрасте. Однако “диким” самкам было всего 2.5 и 1.5 года, а в этом возрасте шимпанзе далеко до изготовления полноценных гнезд и результативного “ужения” муравьев (шимпанзе эффективно осуществляют эти формы поведения в возрасте около 4 лет). Можно предположить, что запечатленные в раннем детстве (после годовалого возраста) стереотипы успешно “разворачивались”, а разница в раннем опыте всего в полгода - год оказалась фатальной для развития стереотипов у остальных особей.
“Культурные” аспекты орудийной деятельности животных. Локальное распространение орудийного поведения в территориально ограниченных группировках животных дает основания предполагать значительную роль социального обучения в его формировании (обзор: Резникова, 2004). Роль подражания в формировании орудийных навыков была неоднократно продемонстрирована в лабораторных опытах. Так, Ванчатовой (Vančatová, 1984) в экспериментах с группой капуцинов впервые было показано, что обезьяны, которые первоначально отбрасывали в сторону предложенные им камни для раскалывания орехов и т-образные “грабли” для доставания лакомых кусочков, быстро перешли к орудийному поведению, подражая доминантным особям, специально обученным экспериментаторами. В естественных условиях вопрос о роли сигнальной наследственности (Лобашев, 1961) в формировании орудийного поведения животных требует для каждого случая специальных исследований, которые могут длиться годами. Ситуации, в которых животные используют орудия, не часты, а документированных свидетельств о культурной передаче навыков совсем немного. Для ряда видов, как это видно из приведенных выше примеров, этот вопрос находится в стадии исследования и накопления пока разрозненных сведений. Например, некоторыми авторами отмечалось, что детеныши каланов перенимают у своих матерей предпочтение орудий определенной формы и манеру использовать эти орудия. Однако систематических исследований на эту тему, по-видимому, не проводилось. Большую ценность представляют редкие случаи многолетнего мониторинга “очагов культуры” животных с момента их возникновения. Так, в одной из популяций японских макак в 1979 г. было впервые зафиксировано применение камня для раскалывания орехов молодой самкой. С тех пор распространение навыка тщательно фиксировалось. Выяснилось, что навыки распространяются от молодых особей к их матерям и товарищам по играм, а в следующем поколении - от матерей к детям. Спустя 20 лет этой техникой владело 80% популяции. Исследователи сравнили действия макак-“неофитов” и обезьян, принадлежащих другой, географически удаленной популяции, в которой обезьяны использовали камни “с незапамятных времен”. Стереотипы поведения оказались неразличимы. Это дало основания полагать, что двигательные комплексы, лежащие в основе орудийной деятельности, обладают видовой спецификой, а распространение навыка в популяции происходит на культурной основе (Huffman, Nishie, 2001).
“Культурные” аспекты орудийного поведения хорошо изучены лишь для шимпанзе. Формы орудийной деятельности шимпанзе в разных местообитаниях варьируют в широких пределах, например от малоэффективного раздавливания термитов в древесных гнездах до тщательного выбора, подготовки и умелого введения подходящего предмета внутрь термитника; от обмакивания в воду единственного листа в неизменном виде до извлечения влаги с помощью “губки” из скомканных листьев; от разбивания плода с твердой скорлупой о неподвижную поверхность (без применения орудия) до использования молота и наковальни. Предметы могут быть использованы в неизменном виде или модифицированы; подобраны в том месте, где они будут применяться, или принесены издалека, так что шимпанзе, выбирая орудие, оперирует “мысленными” представлениями о конечной цели (McGrew, 2004; Whiten et al., 1999). Различия в способах употребления орудий еще в 70-е годы пытались объяснять разнообразием культурных традиций (Goodall, 1970), однако понадобились многолетние тщательные наблюдения для того, чтобы подтвердить эту гипотезу. Действительно, лишь некоторые различия в орудийной деятельности шимпанзе можно объяснить экологическими особенностями: например, в разных местах обитания шимпанзе различные виды термитов и муравьев строят гнезда, требующие разной техники вскрытия (McGrew, 1992). Многие ситуации, однако, поддаются объяснению с привлечением разницы экологических условий. Так, шимпанзе, обитающие в национальных парках Таи и Боссу, разделенных расстоянием в 200 км, используют в камни в качестве молотков и наковален. Существуют некоторые различия в технике, кроме того, шимпанзе в Боссу разбивают орехи масличной пальмы, а в Таи – нет, хотя эти орехи там имеются (Boesch, 1991). Зато шимпанзе Махале и Гомбе не используют молотков, несмотря на обилие в этих местах как камней, так и твердых орехов. Гудолл (Goodall, 1970) отметила единственный для группировки Гомбе случай использования “молотка” и предположила, что эта техника распространится в группировке. Однако в течение последующих 28 лет этого не произошло (Whiten et al., 1999). В основе культурных традиций в группах шимпанзе, по мнению Дж. Гудолл (1992), лежит поведение особей-новаторов. Однако период передачи принципиально новых для популяции навыков в естественных условиях проследить очень трудно. После того, как в сообществе утвердился какой-либо технический прием, он, по предположению приматологов, сохраняется почти неизменным на протяжении бесчисленных поколений. В гранитных наковальнях, употребляемых шимпанзе в Таи и Боссу, обнаружены углубления, которые указывают на то, что их употребляли очень долгое время (Sugiyama, Koman, 1979). Молодые шимпанзе осваивают принятые в сообществе способы использования орудий в раннем детстве в результате подражания и собственной практики, включающей немало проб и ошибок (Whiten et al., 1999; McGrew, 2004). Можно также полагать, что существенную роль в этом процессе играет импринтинг. В пользу этого предположения свидетельствует попытка С. Брюер (1982) научить подростка шимпанзе Уильяма, с младенчества воспитывающегося в обществе людей, добывать термитов с помощью палочки. Самка, попавшая в группу адаптируемых животных в возрасте 6 лет, делала это с легкостью, и ее действия ни в чем не отличались от действий диких шимпанзе. Уильям внимательно наблюдал за ее действиями, но не подражал им. Желая преподать ему наглядный урок, С. Брюер не только медленно и многократно повторяла перед ним требуемые действия, но и съедала добываемых термитов. Это возымело краткий успех, и одного термита Уильям сумел поймать с помощью палочки, но, будучи укушен в палец, больше к этим насекомым не приближался. Огромные различия между Уильямом и старшей самкой наблюдались и в строительстве гнезд, и в разбивании твердых плодов о ветки деревьев, и эти различия напоминают таковые между шимпанзе Фирсова, также попавшими в его группу в разном возрасте. Видимо, по достижении определенного возраста шимпанзе утрачивают способность усваивать некоторые навыки, составляющие картину видотипичного поведения.
Томазелло с соавторами (Tomasello et al., 1993) обнаружили существенные различия в том, как шимпанзе и маленькие дети усваивают навыки обращения с орудиями, наблюдая за действиями демонстратора, т.е. в культурных аспектах их орудийной деятельности. Исследователи работали с двумя группами шимпанзе: “окультуренными”, которые были выращены в неволе и участвовали в экспериментах, требующих контакта с людьми и проявления интеллекта, и “наивными” животными того же возраста, не имеющими контактов с человеком. Использовали 16 разных предметов, которые давали испытуемым в руки - в одной серии опытов сразу же, а в другой - спустя 48 ч после демонстрации. Оказалось, что только при соблюдении двух условий - лабораторное прошлое и отсутствие отсрочки в опытах - орудийное подражательное поведение шимпанзе приближается к таковому у детей. Дети наблюдают за тем, как и с какой целью демонстратор использует предмет, и затем, получив доступ к этому предмету, по возможности точно копируют то, что видели. Обезьяны, воспитанные вне контакта с людьми, каждый раз “изобретают велосипед”: наблюдая за действиями демонстратора, они, получив его орудие, применяют собственный способ манипуляции с ним. Если вспомнить заключение Кёлера (K`hler, 1925) о том, что шимпанзе все делают крайне небрежно, можно не удивляться тому, что культурные традиции в популяциях этих животных не достигают высокого уровня манипуляций с предметами.
ЗАКЛЮЧЕНИЕ
Ранее неоднократно предпринимались попытки представить проявления орудийной деятельности разных видов животных в виде некой таблицы, в которой представители разных видов занимают определенное положение: у одних видов в основе орудийного поведения лежит сигнальная наследственность и подражательное обучение, а у других реализуются врожденные, более или менее жестко запрограммированные, стереотипы поведения. Однако теперь ясно, что пересматривать подобную таблицу пришлось бы слишком часто. Полученные в последние годы данные, касающиеся роли индивидуального опыта и имитационного обучения в шлифовке “наследственных” стереотипов орудийной деятельности животных, заставляют вновь задаваться вопросом об относительной гибкости орудийного поведения.
Вырисовывается следующая картина. Генетически предопределена некая склонность и потенциальная способность к использованию орудий. Некоторые виды обладают врожденной склонностью к манипулированию предметами определенной формы, и эта наследственная программа реализуется при наличии соответствующей мотивации. Так, птицы используют палочки и крючки для добывания пищи, но не пользуются ими, скажем, при постройке гнезд. Сравнительные исследования в этой области находятся пока в начальной стадии, и для многих видов роль сигнальной наследственности в формировании орудийного поведения остается пока практически неисследованной. На данном уровне накопленных знаний можно сказать, что социальное обучение играет в формировании орудийной деятельности видов превалирующую роль у одних видов (человекообразные обезьяны), существенную у других (примером могут служить голубые сойки), подчиненную у третьих (дарвиновы вьюрки). Это не исключает эффективного и быстрого накопления индивидуального опыта при совершенствовании сложных стереотипов (новокаледонские галки). Вероятно, существенную роль в формировании орудийного поведения животных играет встреча с соответствующими стимулами в критические периоды онтогенеза. Во всех случаях, когда в формировании орудийной деятельности участвует в качестве составляющей накопление и совершенствование опыта, можно, по-видимому, говорить о видотипической склонности к образованию специфических ассоциаций. Можно предположить, что слишком быстрое образование специфических ассоциаций может играть тормозящую роль, превращаясь в груз приобретенных стереотипов и препятствуя инновационным решениям. Это предположение нуждается в экспериментальной проверке.
Орудийная деятельность дает возможность интегрального исследования интеллекта животных. Она связана с оценкой обстоятельств, поиском подходящих предметов, планированием действий, коммуникацией с конспецификами. Эксперименты, основанные на решении инструментальных задач, позволяют представить пределы когнитивных возможностей животных и оценить размах индивидуальной вариабельности “интеллектуальных” способностей в группировках. Вопрос о том, реализуется ли наблюдаемый в некоторых экспериментах высокий когнитивный потенциал животных в ситуациях, не требующих использования орудий, или это узко направленная видовая “гениальность”, требует дальнейших исследований.
Работа поддержана грантами РФФИ № 05-04-48604, Совета по грантам Президента РФ (НШ-1038.2003.4) и Президиума РАН по программе "Происхождение и эволюция биосферы".
СПИСОК ЛИТЕРАТУРЫ
Брюер С., 1982. Шимпанзе горы Ассерик. М.: Мир. 279 c.
Вацуро Э.Г., 1948. Исследование высшей нервной деятельности антропоида (шимпанзе). М.: Изд-во АМН СССР. 180 с.
Гудолл Дж., 1992. Шимпанзе в природе: поведение. М.: Мир. 670 с.
Дежкин В.В., Мараков С.В.,1968. Каланы возвращаются на берег. М.:Мысль.199 c.
Зорина З.А. , Дерягина М.А. , Маркина Н.В., 1986. Структура и возрастные особенности манипуляционной активности серой вороны // Зоол. журн. Т. 65, № 10. С. 1552–1559.
Зорина З.А., Полетаева И.И., 2001. Элементарное мышление животных: Учебное пособие по высшей нервной деятельности и зоопсихологии. М.: Аспект Пресс. 320 с.
Ладыгина-Котс Н.Н., 1923. Исследование познавательных способностей шимпанзе. М.: Госиздат. 370 с.
Ладыгина-Котс Н.Н., 1959. Конструктивная и орудийная деятельность высших обезьян. М.: Изд-во АН СССР. 398 с.
Лобашев М.Е., 1961. Сигнальная наследственность // Исследования по генетике. Т. 1. Л.: Изд-во ЛГУ. С. 3–11.
Перри Р., 1974. Мир белого медведя. М. :Гидрометеоиздат. 185 с.
Резникова Ж.И., 2000. Интеллект и язык. Животные и человек в зеркале экспериментов. М.: Наука. 280 с.
Резникова Ж.И., 2004. Сравнительный анализ различных форм социального обучения у животных // Журн. общ. биологии. Т. 65, № 2. С. 136–152.
Резникова Ж.И., 2005. Интеллект и язык животных и человека. Введение в когнитивную этологию. М.: Академкнига. 560 с. [Введение и глава 8]
Резникова Ж.И., Пантелеева С.Н., 2001. Взаимодействие муравьев Myrmica rubra и ногохвосток (Collembola) как охотников и массовой добычи // Докл. Акад. наук. Т. 380, № 4. С. 567–569.
Резникова Ж.И., Пантелеева С.Н., 2003. Экспериментальное исследование этологических аспектов хищничества у муравьев // Успехи соврем. биологии. Т. 3, № 3. С. 234–242.
Резникова Ж.И., Рябко Б.Я., 1990. Теоретико-информационный анализ «языка» муравьев // Журн. общ. биологии. Т. 51, № 5. С. 601–609.
Рогинский Г.З., 1948. Навыки и зачатки интеллектуальных действий у антропоидов (шимпанзе). Л: Изд-во ЛГУ. 250 с.
Смирин В.М., Смирин Ю.М., 1991. Звери в природе. М.: Изд-во МГУ. 265 с.
Фабри К.Э., 1976. Основы зоопсихологии. М.: Изд-во МГУ. 287 с.
Фирсов Л.А., 1977. Поведение антропоидов в природных условиях. Л.: Наука. 160 с.
Фройде М., 1986. Животные строят. М.: Мир. 215 с.
Штодин М.П., 1947. Материалы к вопросу о высшей нервной деятельности человекообразных обезьян (шимпанзе) // Тр. Ин-та эволюционной физиологии и патологии ВНД им. И.П. Павлова. Т.2. Л: Изд-во АН СССР. С. 171–183.
Alcock J., 1972. The evolution of the use of tools by feeding animals // Evolution. V. 26. P. 464–473.
Allen C., Bekoff M., 1997. Species of Mind: The Philosophy and Biology of Cognitive Ethology. Cambridge, Mass.: MIT Press. 636 p.
Anderson J.R., 1996. Chimpanzees and capuchin monkeys: comparative cognition // Reaching into Thought. The Minds of the Great Apes / Eds. Russon A., Bard K., Parker S. Cambridge: Cambridge Univ. Press. P. 23–56.
Armbruster L., 1921. Űber Werkzeuggebrauch bei Tieren // Naturwissenschaften. V. 9. P. 303–305.
Attenborough D., 1998. The Life of Birds. Princeton, New Jersey: Princeton Univ. Press. 320 p.
Aumann T., 1990. Use of stones by the Black-breasted buzzard Hamirostra melanosternon to gain access to egg contents for food // Emu. V. 90. P. 141–144.
Beck B.B., 1980. Animal Tool Behavior: the Use and Manufacture of Tools by Animals. N.Y.: Garland STPM Press. 340 p.
Bekoff M., 2002. The Cognitive Animal. Cambridge, Mass.: MIT Press. 420 p.
Boesch C., 1991. Teaching among wild chimpanzees // Animal Behaviour. V. 41. P 530–532.
Bowman R., 1961. Morphological differentiation and adaptation in the Galapagos finches // Univ. Calif. Publ. Zool. V. 58. P. 1–326.
Call J., 2000. Representing space and objects in monkeys and apes // Cognitive Science. V. 24. P. 397–422.
Chappell J., Kacelnik A., 2002. Tool selectivity in a non-mammal, the New Caledonian crow (Corvus moneduloides) // Animal Cognition. N 5. P. 71–78.
Chappell J., Kacelnik A., 2004. Selection of tool diameter by New Caledonian crows (Corvus moneduloides) // Animal Cognition. N 7. P. 121–127.
Chevalier-Skolnikoff S., Liska J., 1993. Tool use by wild and captive elephants // Animal Behaviour. V. 46. P. 209–219.
Chiang M., 1967. Use of tools by wild macaque monkeys in Singapore // Nature. V. 214. P. 1258–1259.
Darwin C.R., 1871. The Descent of Man. V. 1. L: Murray. 320 p.
Douglas-Hamilton I., Douglas-Hamilton O., 1975. Among the Elephants. N.Y.: Viking. 340 p.
Eibl-Eibesfeldt I., 1961. Űber den Werkzeuggebrauch des Spechtfinken Camarhynchus pallidus (Slater und Slavin) // Zeitchrift fhr Tierpsychologie. H. 18. S. 343–346.
Evans H., Eberhard M., 1970. The Wasps. Ann Arbor: Univ. Michigan Press. 435 p.
Fellers J.H., Fellers G. M., 1976. Tool use in a social insect and its implications for competitive interactions // Science. V. 192. N 4234. P. 70–72.
Fisher E., 1939. Habits of the southern sea otter // J. Mammal. V. 20. P. 21–36.
Fisher J., Hinde R.A., 1949. The opening of milk bottles by birds // British Birds. V. 42. P. 347–357.
Frisch J., 1940. Did the Peckhams witness the invention of a tool by Ammophila urinaria? // Amer. Midl. Nat. V. 24. P. 345–350.
Gardner B.T., Gardner R.A., 1969. Teaching sign language to a chimpanzee // Science. V. 165. P. 664–672.
Gifford E., 1919. Field notes on the land birds of the Galapagos Islands and of Cocoas Islands, Costa Rica // Proc. Calif. Acad. Sci. V. 2. P. 189–258.
Goodall J., 1970. Tool using in primates and other vertebrates // Advances in the Study of Behavior. V. 3 / Eds. Lehraman D.S., Hinde R.A., Shaw E. N.Y.: Acad. Press. P. 195–249.
Griffin D.R., 1976. The Question of Animal Awareness: Evolutionary Continuity of Mental Experience. N.Y.: Rockefeller Univ. Press. 285 p.
Griffin D.R., 1992. Animal Minds. Chicago: Chicago Univ. Press. 320 p.
Hall K., Schaller G., 1964. Tool-using behavior of the California sea otter // J. Mammal. V. 45. P. 287–298.
Harlow H., 1951. Primate learning // Comparative Psychology / Ed. Stone C. N.Y.: Prentice Hall. P. 183–238.
Hart B.L., Hart L.A., McCoy M., Sarath C.R., 2001. Cognitive behaviour in Asian elephants: use and modification of branches for fly switching // Animal Behaviour. V. 62. P. 839–847.
Hauser M.D., 1988. How infant vervet monkeys learn to recognize starling alarm calls: The role of experience // Behaviour. V. 105. P. 187–201.
Hauser M.D., Kralik J., Botto-Mahan C., 1999. Problem solving and functional design features: Experiments with cotton-top tamarins (Saguinus oedipus) // Animal Behaviour. V. 57. P. 565–582.
Herman L.M., 1986. Cognition and language competencies in bottlenosed dolphins // Dolphin Cognition and Behavior. A Comparative Approach / Eds. Schusterman R.J., Thomas J.,Wood F.G. Hillside N.J.: Lawrence Erlbaum Associates. P. 221–251.
Hobbs J., 1971. Use of tools by the white-winged chough // Emu. V. 71. P. 84–85.
Hobhouse L.T., 1901 (re-edit. 1998). Mind in Evolution / Eds Wozniak R.H. Series: (T-CP) Thoemmes Press, Classics in Psychology. N.Y.: Thoemmes Press. 240 p.
Hölldobler B., Wilson E. O., 1990. The Ants. Berlin : Springer Verlag. 732 p.
Houk J., Geibel J., 1974. Observation of underwater tool use by the sea otter, Enhydra Lutris Linnaeus // Calif. Fish Game. V. 60. P. 207–208.
Huffman M. A., Nishie H., 2001. Stone handling, a two decade old behavioural tradition in a Japanese monkey troop // Advances in Ethology. V. 36. Contributions to the XXVII Internat. Ethological Conference. Berlin; Vienna: Blackwell. P. 35–35.
Hunt G.R., 1996. Manufacture and use of hook-tools by New Caledonian crows // Nature. V. 379. P. 249–251.
Hunt G.R., Gray R.D., 2004. Direct observations of pandanus-tool manufacture and use by a New Caledonian crow (Corvus moneduloides) // Animal Cognition. V. 7, N 2. P. 114–120.
Hunt, G.R. 2000. Human-like, population-level specialization in the manufacture of pandanus tools by New Caledonian crows Corvus moneduloides // Proc. Royal Society L. Series B. N 267. P. 403–413.
Janes S., 1976. The apparent use of rocks by a raven in nest defence // Condor. V. 78. P. 409.
Jones T., Kamil A.C., 1973. Tool-making and tool-using in the Northen blue jay // Science. V. 180, N 4090. P. 1076–1078.
Kacelnik A., Chappell J., Kenward B., Weir A.A.S., 2004.Cognitive adaptations for tool-related behaviour in New Caledonian Crows // Comparative Cognition: Experimental Explorations of Animal Intelligence / Eds. Wasserman E.A., Zentall T.R. N.Y.: Oxford Univ. Press. P. 200–226.
Katz S., 1975. Tool-using behaviour of the pocket gopher (Thomomys bottae) // Proc. of 55th Annual Meeting of the American Society of Mammalogists. Missoula, Montana: Montana Univ. Press. P. 240–242.
Kenward B., Weir A.A.S, Ritz C., Kacelnik A., 2005. Tool manufacture by naïve juvenile crows // Nature. V. 433. P. 121.
Kenyon K., 1969. The Sea Otter of the Eastern Pacific Ocean. Washington D.C.: US Bureau of Sport Fisheries and Wildlife. 190 p.
Klüver H.,1933. Behavior Mechanisms in Monkeys. Chicago: Chicago Univ. Press. 270 p.
Köhler W., 1925. The Mentality of Apes. L.: Routledge and Kegan Paul (Reprint ed. N.Y.: Liverigh. 1976). 580 p.
Kralik J.D., Hauser M.D., 2002. A nonhuman primate’s perception of object relations: experiments on cottontop tamarins, Saguinus oedipus // Animal Behaviour. V. 63. P. 419–435.
Lack D., 1947. Darwin’s Finches. Cambridge: Cambridge Univ. Press. 290 p.
Lawick-Goodall J., van, Lawick H., van, 1966. Use of tools by the Egyptian vulture Neophron pernopterus // Nature. V. 212. P. 1468–1469.
Levey D.J., Duncan R.S C.F., Levins C.F., 2004. Use of dung as a tool by burrowing owls // Nature. V. 431. P. 39–42.
Limongelli L., Boysen S., Visalberghi E., 1995. Comprehension of cause and effect relationships in a tool-using task by common chimpanzees (Pan troglodytes) // J. Compar. Psychol. V. 109. P. 18–26.
Lovell H., 1958. Baiting of fish by a green heron // Wilson Bull. V. 70. P. 280–281.
Marler P., 1970. A comparative approach to vocal learning: song development in white-crowned sparrows // J. Compar. Psychol. V. 71, N 2. P. 1–25.
Masson J.M., 1995. When Elephants Weep: the Emotional Lives of Animals. N.Y.: Delacorte Press. 240 p.
McGrew W.C., 1992. Chimpanzee Material Culture: Implications for Human Evolution. Cambridge M.A.: Cambridge Univ. Press. 240 p.
McGrew W.C., 2004. The Cultured Chimpanzee: Reflections on Cultural Primatology. Cambridge: Cambridge Univ. Press. 248 p.
Millikan G., Bownam R., 1967. Observations on Galapagos tool-using finches in captivity // Living Bird. N 6. P. 23–41.
Miyamoto M., Koop B.F., Slightom J.L., Goodman M., Tennant M.R., 1988. Molecular systematic of higher primates: genealogical relations and classification // Proc. Natl. Acad. Sci. USA. V. 85. P. 7627–7631.
Montevecchi W., 1978. Corvids using objects to display gulls from nests // Condor. V. 80. P. 349–349.
Morrill W., 1972. Tool using behaviour of Pogonomyrmex badius (Hymenoptera: Formicidae) // Florid Entomol. V. 55. P. 59–60.
Morris D., 1954. The snail-eating behaviour of thrushes and black-birds // Brit. Birds. V. 47. P. 33–49.
Morse D., 1968. The use of tool by brown-headed nuthatches // Wilson Bull. V. 80. P. 220–224.
Moss C., 1982. Portraits in the Wild: Behavior Studies of East African mammals. Chicago: Chicago Univ. Press. 560 p.
Nissen H.W., 1931. A field study of the chimpanzee // Comp. Psychol. Monogr. V. 8. P. 1–121.
Ottoni E.B., Mannu M., 2001 Semi-free-ranging tufted capuchins (Cebus apella) spontaneously use tools to crack open nuts // Int. J. Primatol. V. 22, N 3. P. 347–358.
Owing D., Coss R., 1977. Snake mobbing by California ground squirrels: Adaptive variation and ontogeny // Behaviour. V. 62. P. 50–69.
Pepperberg I.M., 1987. Acquisition of the same–different concept by an African grey parrot (Psittacus errithacus ): Learning with respect to categories of colour, shape and material // Anim. Learn. and Behav. V. 15. P. 423–432.
Povinelli D.J., 2000. Folk Physics for Apes: a Chimpanzee’s Theory of How the World Works. Oxford: Oxford Univ. Press. 280 p.
Reaux J.E., Theall L.A., Povinelli D.J., 1999. A longitudinal investigation of chimpanzees’ understanding of visual perception // Child Development. V. 70. P. 275–290.
Reznikova Zh., 2001. Interspecific and intraspecific social learning in ants // Advances in Ethology (XXVII Int. Ethological Conference). N 36 / Eds. Apfelbach R., Fendt M., Krämer S., Bjőrn M.S. Berlin–Vienna: Blackwell. P. 108.
Reznikova Zh.I., 1995. The role of individuals in an ant colony // Ecological Progress: Current Status and Perspectives. Abstracts of European Ecological Congress. Budapest. P. 87.
Reznikova Zh.I., Ryabko B.Ya., 1997. Distant homing and cognition in ants // Advances in Ethology (XXV Int. Ethological Conference). N 32. Eds. Taborsky M., Taborsky B. Berlin; Vienna: Blackwell. P. 192.
Rumbaugh D.M., 1974. Comparative Primate Learning and its Contributions to Understanding Development, Play, Intelligence, and Language // Perspectives in Primate Biology / Ed. Chiarelly B. N.Y.: Plenum. P. 253–281.
Savage-Rumbaugh E.S., 1986. Ape Language: From Conditioned Response to Symbol. N.Y.: Columbia Univ. Press. 398 p.
Schaik C.,van ,Dopyera C., 1997. The Red Ape's Surprise // Wildlife Conservation. January/February. P. 36–43.
Shettleworth S.J., 1990. Spatial memory in food-storing birds // Philos. Trans. R. Soc. L. Series B. N 329. P. 143–151.
Shettleworth S.J., 1998. Cognition, Evolution, and Behavior. N.Y.: Oxford Univ. Press. 460 p.
Sisson R., 1974. Aha! It really works! // Nat. Geogr. Mag. V. 145. P. 142–147.
Struhsaker T., Leland L., 1977. Palm-nut smashing by Cebus apella in Colombia // Biotropica. V. 9. P. 124–126.
Sugiyama Y., Koman J., 1979. Tool using and making behavior in wild chimpanzees at Bossou, Guinea // Primates. V. 20. P. 513–524.
Tebbich S., Bshary R., 2004. Cognitive abilities related to tool use in the woodpecker finch, Cactospiza pallida // Animal Behaviour. V. 57. P. 689–697.
Tebbich S., Taborsky M., Fessl B., 2002. The ecology of tool use in the woodpecker finch Cactospiza pallida // Ecology Letters. N 5. P. 656–664.
Tebbich S., Taborsky M., Fessl B., Blomqvist D., 2001. Do woodpecker finches acquire tools use by social learning? // Proc. Royal Soc. L. Series B. V. 268. P. 2189–2193.
Thouless C.R., Fanshawe J.H., Bertram B.C.R., 1987. Egyptian vultures Neophron pernopterus and Ostrich Struthio camelus eggs – the origins of stone-throwing behavior // Ibis. V. 131. P. 9–15.
Tomasello M., Savage-Rumbaugh S., Kruger A. C., 1993. Imitative learning of actions on objects by children, chimpanzees, and enculturated chimpanzees // Child Development. V. 64. P. 1688–1705.
Vančatová M., 1984. The influence of imitation on tool using in capouchine monkeys (Cebus apella) // Anthropologie. V. 22, N 1. P. 1–2.
Visalberghi E., 1990. Tool use in Cebus // Folia Primatologica. V. 54. P. 146–154.
Visalberghi E., 2002. Insights from capuchin monkeys' studies: ingredients of, recipes for, and flaws in capuchins' success // The Cognitive Animal: Empirical and Theoretical Perspectives on Animal Cognition / Eds. Bekoff M., Allen C., Burghardt G.. Cambridge: MIT Press. P. 1–11.
Visalberghi E., Fragaszy D. M., Savage-Rumbaugh S., 1995. Comprehension of causal relations in a tool-using task by chimpanzees (Pan troglodytes), bonobos (Pan paniscus), orang utans (Pongo pygmaeus), and capuchins (Cebus apella) // J. Compar. Psychol. V. 109. P. 52–60.
Visalberghi E., Trinca L., 1989. Tool use in the capuchin monkeys: distinguishing between performing and understanding // Primates. V. 30. P. 511–521.
Waal F.B.M., de, 1995. Bonobo sex and society. The behavior of a close relative challenges assumptions about male supremacy in human evolution // Scientific Amer. March. P. 82–88.
Walsh J.F., Grunewald J., Grunewald B., 1985. Green-backed heron (Butorides striatus) possibly using apparent bait // J. für Ornitologie. S. 126. S. 439–442.
Watson J., 1908. Imitation in monkeys // Psychol. Bull. N 5. P. 169–178.
Weir A.A.S., Kenward B., Chappell J., Kacelnik A., 2004. Lateralization of tool-use in New Caledonian crows Corvus moneduloides // Proc. Royal Soc. L. Ser. B. Biol. Letters. V. 271. P. 344–346.
Whitten A., Goodall J., McGrew W.C.,Nishida T., Reynolds V., Suglyama Y., Tutin C.E., Wrangham R. W., Boesch C., 1999. Culture in chimpanzees // Nature. V. 399. P. 682–685.
Yerkes R.M., 1916. The Mental Life of Monkeys and Apes: A Study of Ideational Behavior. Behavior Monographs. Baltimore. 430 p.