

Сравнительная морфофизиологическая характеристика соматического и вегетативного отделов нервной системы
Соматический отдел нервной системы обеспечивает сенсорно-моторные функции тела, то есть иннервирует кожу, начальные отделы пищеварительного и дыхательного трактов, органы чувств, опорно-двигательный аппарат, в частности поперечнополосатую, произвольную мускулатуру, сокращение которой зависит от нашего сознания.
Вегетативный отдел нервной системы иннервирует гладкую, непроизвольную мышечную ткань, входящую в состав стенки внутренних органов, железы внешней и внутренней секреции, сосуды, сердце. Работа всех этих органов происходит постоянно, ритмично и не подчиняется нашей воле. Всё это и послужило поводом для английского физиолога Ленгли (1903) назвать вегетативную нервную систему «автономной», функционирующей независимо от нашего сознания.
В 1955 году термин «автономная нервная система» вошел в состав Международной Парижской анатомической номенклатуры (PNA). В нашей отечественной анатомической литературе эквивалентом «автономной» оставлен термин «вегетативная нервная система». Тем самым подчеркивается, во-первых, условность деления нервной системы на соматический и вегетативный отделы, во-вторых, что деятельность всех внутренних органов находится под постоянным влиянием «сомы» и контролем высших нервных центров. Примерами справедливости этих методологических подходов служат все закономерности биологических, в том числе физиологических, процессов в организме. Так, в ответ на «соматические» мышечные сокращения изменяются частота сердечных сокращений, сосудистый тонус, питание тканей, теплообмен. Поэтому считается, что мышечная система является главным биологическим стимулятором развития и совершенствования всех других систем организма.
«Автономия» вегетативной регуляции внутренних органов, вне зависимости от «сомы» и высших нервных центров, не укладывается в такие понятия, как психотерапия, самовнушение, физиотерапия, иглоукалывание, аутотренинг и т.п. Всё это возможные пути коррекции деятельности внутренних органов через сомато-висцеральные рефлексы, замыкание которых происходит на уровне центральной нервной системы.
Отличительной особенностью строения вегетативной нервной системы от соматической является повсеместное распространение ее в теле человека. Если соматическая нервная система в своей эфферентной (двигательной) части ведает специальной функцией сокращения скелетных мышц, то вегетативная, обладая универсальной адаптационно-трофической ролью, имеет обширную, повсеместную область эфферентации, в том числе и в элементах «сомы».
Другое принципиальное различие между соматической и вегетативной частями нервной системы состоит в строении рефлекторных дуг (колец). Для той и другой системы характерна простая трехнейронная цепь, представленная афферентным, вставочным и эфферентным нейронами. Основная разница между ними заключается в положении отдельных звеньев рефлекторной дуги (рис. 5).
Афферентное звено соматической и вегетативной рефлекторной дуги устроено однотипно. Оно представлено псевдоуниполярными клетками чувствительных узлов спинномозговых и некоторых черепных нервов, содержащих чувствительный компонент. Цитоплазматический вырост таких клеток делится Т-образно на периферический отросток - дендрит, связанный с рецепторами тела или внутренних органов, и центральный - аксон (нейрит). Последний вступает в спинной или головной мозг и передает нервный импульс на вставочный (ассоциативный, кондукторный) нейрон. Вставочное звено как в соматическом, так и вегетативном отделах многонейронно, отличается мультиполярностью и имеет сложные связи с различными отделами центральной нервной системы. При этом, ассоциативные нейроны соматического отдела не выходят за пределы спинного или головного мозга, передавая нервный импульс на расположенные здесь эфферентные нейроны двигательных ядер. В вегетативной же нервной системе нейроны эфферентного звена «вынесены» за пределы центральной нервной системы и образуют на периферии вегетативные, соответственно симпатические и парасимпатические, узлы.
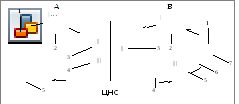
Рис. 5. Схема рефлекторной дуги соматического (А) и вегетативного (Б)
отделов нервной системы:
I – афферентный нейрон; II – вставочный, ассоциативный нейрон;
III – эфферентный нейрон;
А: 1 – рецептор кожи; 2 – чувствительный узел спинномозгового или черепного нерва;
3 – вставочный нейрон; 4 – двигательное ядро; 5 – скелетная мышца;
Б: 1 – рецептор внутреннего органа; 2 – чувствительный узел спинномозгового или черепного нерва;
3 – сегментарный вегетативный центр; 4 – преганглионарное волокно; 5 – вегетативный узел;
6 – постганглионарное волокно; 7 – рабочий орган.
Все вегетативные узлы представляют собой скопления тел эфферентных нейронов периферического отдела вегетативной нервной системы. Кроме вегетативных узлов вне ЦНС располагаются чувствительные узлы спинномозговых и черепных нервов. По строению и физиологии они принципиально отличаются от вегетативных узлов (рис. 6). Псевдоуниполярные клетки чувствительных узлов спинномозговых и черепных нервов межнейронных синапсов на периферии не имеют. Нервный импульс, возникший в рецепторном аппарате тела или внутренних органов, по дендритам достигает тела нейрона чувствительного узла и транзитом проходит через нервную клетку на аксон. В вегетативных же узлах происходит передача нервного импульса с ассоциативного нейрона на эфферентный посредством синапсов.
Весьма важной особенностью строения вегетативных узлов является принцип мультипликации: преганглионарное волокно, входя в узел, многократно ветвится на свои концевые отделы, которые заканчиваются на десятках нервных клетках. Эти соотношения выражаются как 1:32 (Тонков В.Н.. 1962). Таким образом, при минимальном количестве преганглионарных волокон в нервный процесс вовлекается большое количество периферических нейронов, постганглионарные волокна которых непосредственно связаны с рабочими структурами.

А Б
Рис. 6. Схема строения чувствительного (А) и вегетативного (Б) узла:
а – аксон; d – дендрит; R – рецептор; 1 – псевдоуниполярная клетка чувствительного узла;
2 – эфферентный нейрон вегетативного узла; 3 – синапс; 4 – вставочный нейрон.
Как отмечалось выше, передача нервного импульса с нейрона на нейрон или с нейрона на орган - это сложный электронно-химический процесс, в реализации которого особая роль принадлежит биологически активным веществам - медиаторам. Межнейронная передача в вегетативной нервной системе осуществляется с помощью ацетилхолина, катехоламинов (адреналин, норадреналин), серотонина и других веществ химической медиации. При этом главным медиатором для парасимпатического отдела нервной системы является ацетилхолин: передача импульса с аксона вставочного нейрона (преганглионарного волокна) на дендрит или тело эфферентного нейрона, а также с аксона последнего на орган - исполнитель (постганглионарное волокно) происходит с помощью ацетилхолина (холинэргические волокна). В симпатическом отделе вегетативной нервной системы передача импульса с аксона вставочного нейрона (преганглионарного волокна) на эфферентный нейрон симпатических узлов так же осуществляется посредством ацетилхолина, а с постганглионарного звена на рабочий орган - с помощью адреналина и норадреналина (адренэнергические волокна). (Рис. 7).
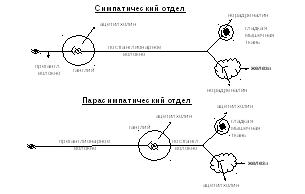
Рис. 7. Схема медиации симпатического и парасимпатического отделов
вегетативной нервной системы.
Различия в химической медиации симпатического и парасимпатического отделов вегетативной нервной системы лежат в основе избирательного (специфического или неспецифического) действия лекарственных препаратов. Установлено, что многие наркотические вещества (например, никотин) оказывают парализующее влияние на синапсы. Поэтому при воздействии раствором никотина на вегетативный узел передача нервного импульса блокируется, тогда как при таком же воздействии на чувствительный узел передача нервного импульса сохраняется.
С морфологических позиций расшифровка механизмов синаптической передачи нервного импульса позволяет считать вегетативный узел не просто скоплением нервных клеток на периферии, а специфическим звеном нейро-синаптических взаимодействий, периферическим центром вегетативных регуляций. Поэтому нервный импульс справедливо сравнивают с «дымом из трубы», а вегетативный узел - это «печь» со своим клеточным и синаптическим аппаратом.
Локализация вегетативных узлов в теле человека, на первый взгляд, характеризуется определённой хаотичностью, но с филогенетических позиций и морфо-функционально она строго обусловлена. В целом вегетативные узлы могут располагаться вблизи ЦНС, в отдалении от мозга, вблизи от внутренних органов и внутриорганно (интрамурально). Разделение их на узлы I, II и III порядка в зависимости от отдаленности от мозга следует считать ошибочным, так как порядковая классификация (ранжирование) предполагает зависимость одних образований от других. В морфофизиологии вегетативных узлов такой зависимости не наблюдается.
Симпатические узлы располагаются по бокам позвоночного столба и носят название паравертебральных (околопозвоночных). Соединяясь между собой межузловыми ветвями, они образуют парные симпатические стволы, truncus sympathicus.
Другая группа симпатических узлов получила название превертебральных. Наиболее постоянными из них являются: парные чревные узлы, ganglia coeliaca, они находятся у места отхождения от аорты чревного ствола; парные аортопочечные узлы, ganglia aortorenalis, – у начала почечных артерий; верхний, ganglion mesentericum superior, и нижние (1-2) брыжеечные узлы, ganglion mesentericum inferior, – у места отхождения соответствующих артерий.
Парасимпатические узлы располагаются около– или внутриорганно. К околоорганным относятся: ресничный узел, ganglion ciliare, расположенный в глазнице; ушной, ganglion oticum, лежит на основании черепа у овального отверстия; крылонебный, ganglion pterygopalatinum, - в крыловидно-небной ямке; поднижнечелюстной, ganglion submandibulare, и подъязычный, ganglion sublinguale, узлы – в области соименных слюнных желез.
В соответствии с особенностями расположения (удаления) вегетативных узлов по отношению к мозгу и рабочим органам длина пре- и постганглионарных волокон будет различной: в симпатическом отделе преганглионарные волокна короткие, а постганглионарные длинные; напротив, парасимпатические преганглионарные волокна длинные, а постганглионарные – короткие.
Все преганглионарные волокна по гистофизиологии являются мякотными (миелиновыми): на осевой цилиндр их концентрически намотаны оболочки леммоцитов (без нейроплазмы), состоящими из липопротеидных комплексов (78% липидов и 22% белка). Такие волокна имеют перехваты Раньве, импульс по ним проходит скачкообразно, но с большой скоростью - от 2 - 4 до 10 - 20 м/сек.
Постганглионарные волокна в большинстве случаев безмякотные, осевой цилиндр их погружен в шванновские клетки, скорость проведения импульса по ним небольшая – 1 - 2 м/сек. Учитывая особенности длины и строения пре- и постганглионарных волокон, различную скорость проведения импульса, можно предположить роль того или иного вегетативного звена в режиме деятельности органа в нормальных и патологических условиях.
Отростки ассоциативных нейронов вегетативной нервной системы в составе спинномозговых или черепных нервов выходят из мозга под названием преганглионарных (предузловых) волокон и синаптически заканчиваются на клетках вегетативных узлов. Аксоны нервных клеток вегетативных ганглиев достигают рабочих органов в виде постганглионарных волокон.
Кроме «классической» рефлекторной дуги центрального направления, то есть связанной с головным или спинным мозгом, в стенках внутренних органов (интрамурально) находятся вегетативные сетчато-ганглиозные структуры. Нервные клетки их образуют местные двух- или трехчленные рефлекторные дуги с присущими им афферентным, ассоциативным и эфферентным звеньями. Это собственный нервный аппарат внутренних органов. Он не является принадлежностью ни симпатической, ни парасимпатической нервной системы, поэтому выделяется в самостоятельный отдел – метасимпатический, pars metasympathica (рис. 8).
Афферентное звено местной висцеральной рефлекторной дуги представлено крупными нервными клетками овальной или грушевидной формы. При импрегнации азотнокислым серебром они интенсивно окрашиваются, тогда как ядро остается светлым (II тип клеток Догеля). Дендриты таких клеток посредством рецепторного аппарата регистрируют все изменения внутреннего состояния органа, а аксоны их передают информацию на эфферентные нейроны (I тип клеток Догеля). Отростки эфферентных нейронов оканчиваются на гладкомышечных и железистых элементах внутренних органов.
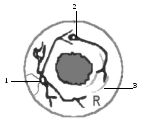
Рис. 8. Схема местной (метасимпатической) рефлекторной дуги:
1 – афферентный нейрон (клетки II типа Догеля); 2 – эфферентный нейрон (клетки I типа Догеля);
3 – стенка внутреннего органа.
Основное назначение интрамуральных ганглиозных образований метасимпатического отдела нервной системы заключается в обеспечении автоматизма и, таким образом, относительной функциональной автономии внутренних органов. Известно, например, что выделенные из организма некоторые органы при определённых условиях могут продолжать свою деятельность: изолированное сердце лягушки при орошении физиологическим раствором продолжает сокращаться; сохраняют свои движения кишечник, отрезки кровеносных сосудов, матка, желчный пузырь. При повреждении спинного мозга или после удаления симпатических стволов (в эксперименте на животных) сохраняется работа сердечно-сосудистой, пищеварительной систем, остаются возможными оплодотворение половых клеток и даже наступление беременности.
Однако, все эти процессы протекают однотипно, монотонно. Организм лишается самого главного - защитных и адаптивных реакций в ответ на изменения внешней среды, которые осуществляются при посредстве симпатической и парасимпатической иннервации. Животные, подвергшиеся симпатэктомии, лишены чувства страха, защитных и агрессивных реакций. Они плохо переносят холод, чрезмерные или даже умеренные физические нагрузки, не адекватно реагирует на эмоциональные стрессы. Это свидетельствует о том, что местные ганглиозные скопления метасимпатического отдела находятся под постоянным нервным контролем со стороны центральной нервной системы.
После децентрализации вегетативных регуляций организм и его органы лишаются адаптационно-трофической функции и способности к нормальному существованию в постоянно меняющихся условиях окружающей среды. Ганглиозные образования метасимпатической нервной системы в таких случаях работают в режиме лишь пусковых механизмов (тормозящих или возбуждающих) под влиянием местных механических, химических, биологических и других раздражителей (рН ткани, давления при наполнении органов, гуморальных факторов и т.п.).