

Особенности проведения возбуждения по миелинизированным и немиелинизированным нервным волокнам. Скорость проведения возбуждения по нервным и мышечным волокнам. Классификация нервных волокон по скорости проведения возбуждения. Законы проведения возбуждения по нервным волокнам.
Мембрана
нейрона и его отростков (нервных волокон)
обладает способностью не только
генерировать потенциал действия, но и
передавать го на относительно большие
расстояния без уменьшения его амплитуды
и расходования энергии, поэтому нервные
импульсы распространяются без
затухания (бездекрементно). Способность
к распространению возбуждения связана
с тем, что во время потенциала действия
происходит изменение знака заряда в
возбужденном участке мембраны (инверсия
заряда). Между ним и невозбужденными
соседними участками мембраны возникают
локальные электрические токи, под
действием которых происходит деполяризация
новых соседних участков, что приводит
к формированию в них потенциала действия.
За счет последовательного охвата
возбуждением все новых участков
нервного волокна возбуждение перемещается
вдоль дендритов и аксонов. Чем больше
диаметр волокна, тем больше скорость
распространения возбуждения. Особенности
проведения возбуждения в нервных
волокнах определяются наличием или
отсутствием у них оболочки. Нервные
волокна делятся на миелиновые
и безмиелиновые,
то есть покрытые особой миелиновой
оболочкой или не покрытые. Эта оболочка
образуется на периферии шванновскими
клетками, а в головном мозге – клетками
олигодендроглии.
Механизм проведения импульса по нервным волокнам объясняется теорией “местных токов” Тасаки. В безмиелиновом нервном волокне ПД генерируется каждой точкой поверхности мембраны – так называемое непрерывное проведение возбуждения. Во время пикового ПД на наружной поверхности волокна происходит перезарядка, в результате чего возникает местный ток от соседнего положительно заряженного участка к возбужденному, а внутри – наоборот. Таким образом образуется кольцевой ток. В миелиновых волокнах слой миелина не целиком охватывает волокно, а имеет промежутки – перехваты Ранвье. Сам миелин обладает высоким сопротивлением электрическому току, поэтому в участках, покрытых миелином, потенциал действия генерироваться не может. Перехваты Ранвье располагаются примерно через каждые 0,5-1,5 мм (в зависимости от толщины оболочки), в них обнаружено очень много (около 12 тыс/мкм2) натриевых каналов, поэтому в миелиновых волокнах проведение возбуждения происходит скачкообразно (сальтаторно). Между соседними участками возникает большой по величине ток (ПД = 100-120 мВ), который в 3-5 раз превышает порог раздражения и способен возбудить соседние участки, что составляет фактор надежности. Скорость проведения возбуждения пропорциональна корню квадратному от диаметра волокна. Самые тонкие волокна – безмиелиновые, а в миелиновых волокнах чем толще слой миелина, тем больше расстояние между перехватами Ранвье. В безмиелиновых волокнах местные кольцевые токи последовательно распространяются “маленькими шажочками” непрерывно, скорость проведения возбуждения в безмиелиновых волокнах гораздо ниже, чем в миелиновых. Такая передача импульса является медленной и неэкономичной в плане энергозатрат. Поэтому безмиелиновыми являются только самые тонкие волокна (менее 1 мкм в диаметре), проводящие возбуждение к медленно работающим органам – кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре.
Виды мышечных волокон. Иннервация скелетной мышцы. Нейромоторная единица.
Типы нервных волокон:
1) медленные, неутомляемые (красные, статические, тонические) – это тонкие, богатые кровеносными сосудами и миоглобином мышцы, во время работы проявляют большую силу, долго не утомляются, но скорость их сокращений небольшая. Например, они сохраняют вертикальную статику, удерживают в определенном положении отдельные части тела, т.е. осуществляют опорную функцию. К ним также относятся наружные мышцы глазного яблока. Медленные фазические сокращения обеспечивают тонус мышц, и поэтому такие сокращения называются тоническими. Они необходимы для поддержания равновесия в статике и динамике. Медленные мышечные клетки составляют основную массу двигательных единиц. В них много миоглобина и миозина, где происходит окисление. Такие мышцы имеют красный цвет и мало утомляются.
2) быстрые, легко утомляемые (белые, динамические, фазические): они имеют толстые мышечные пучки, меньше кровеносных сосудов и миоглобина, скорость сокращений их велика так же, как и утомляемость. Уступая в силе, они способны производить разнообразные мелкие быстрые движения. Быстрые фазические аэробные мышцы немного бледнее, поскольку в них меньше миоглобина, но сохраняется еще достаточно большое количество миозина, а следовательно, интенсивно протекают процессы окисления. В таких мышцах утомление развивается быстрее, чем в выше описанных. По количеству мышечных клеток в моторной единице быстрые фазические мышцы занимают второе место после медленных. Анаэробные мышцы обеспечивают самые быстрые сокращения. В них мало миоглобина и миозина. Клетки, входящие в состав быстрых анаэробных мышц имеют белый цвет. В таких мышцах протекает анаэробный гликолиз, поэтому, в результате накопления недоокисленных продуктов (молочной кислоты), развивается кислородный долг, и как следствие, самое быстрое утомление. Примером таких мышц могут служить мышцы пальцев рук и глаза.
3) быстрые, устойчивые к утомлению (промежуточные).
Все
три типа волокон могут содержаться в
одной и той же мышце, и соотношение их
числа определяется в значительной
степени наследственностью. Например,
в четырехглавой мышце бедра человека
процент медленных волокон может
составлять от 40 до 98 %. Чем больше медленных
волокон, тем больше мышца приспособлена
к работе на выносливость. И наоборот,
люди с высоким процентом быстрых сильных
волокон более способны к работе, требующей
большой силы и скорости сокращения
мышц.
Сила сокращения мышц определяется числом активных мышечных волокон, участвующих в сокращении, частотой нервных импульсов и наличием синхронизации активности отдельных мышечных волокон во времени. Даже в покое скелетные мышцы редко бывают полностью расслабленными. Обычно в них сохраняется некоторое напряжение – тонус. Тонус мышц увеличивается после тяжелых физических упражнений и во время психоэмоционального напряжения.
К каждой мышце подходит много двигательных нейронов , которые ветвятся и иннервируют группы мышечных волокон . Каждая такая группа вместе с иннервирующим ее аксоном называется двигательной единицей . При надлежащей стимуляции все мышечные волокна в ней сокращаются одновременно. Число волокон в двигательной единице варьирует и зависит от того, насколько сложным должно быть управление данной мышцей. Например, в глазодвигательной мышце это число составляет около 10, а бицепсе - более 1000.
В месте контакта двигательного аксона с мышечным волокном формируется нервно-мышечное соединение , или двигательная концевая пластинка . Здесь аксон не имеет миелиновой оболочки и его ветвящиеся окончания погружены в разветвленные бороздки концевой пластинки.
Скелетная мышца и иннервирующий ее нерв состоит из множества нейромоторных единиц. Мышца иннервируется несколькими мотонейронами, аксоны в составе двигательного нерва подходят к мышце. Каждый аксон, войдя в мышцу ветвится на множество веточек, конечная веточка в свою очередь, может иннервировать одно или несколько мышечных волокон. Двигательный нейрон и иннервируемая им группа мышечных волокон образует нейромоторную единицу. Нейромоторная единица работает как единое целое. В нормальных условиях импульсы, посылаемые мотонейроном приводят в действие все мышечные волокна, входящие в состав данной моторной единицы. Т.е. нейромоторная единица является элементарной, структурной и функциональной единицей нервно – мышечного аппарата. Благодаря тому, что каждая скелетная мышца состоит из множества функциональных единиц (их может насчитываться в крупных мышцах несколько сотен), она сокращается при ра боте не вся, а по частям. Это – то и используется при регуляции скорости и силы мышечного сокращения. Но сила сокращения может также зависеть и регулироваться в пределах и самой отдельной нейромоторной единицы, не за счет изменения величины нервных импульсов, а в результате изменения частоты их следования, чем больше частота, тем больше интенсивность сокращения. Но все - таки сила сокращения прежде всего зависит от количества функционирующих нейромоторных единиц. Сила сокращения будет меньше, чем меньшее число нейромоторных единиц вовлечено в сокращение, то есть в этом случае в мышце сокращаются небольшие ее участки, увеличение силы сокращения обусловлено вовлечением все большего и большего количества нейромоторных единиц, входящих в состав данной мышцы.
Таким образом, скорость сокращения и расслабления может регулироваться и скоростью вовлечения в сокращение определенного количества или выключением из процесса сокращения определенного количества нейромоторных единиц. Нейромоторные единицы не являются однородными, а в процессе эволюции они приспособились к определенной функции, произошла их специализация. Одни из них приспособлены для быстрых, фазных движений – так называемые быстрые нейромоторные единицы, другие – для медленных тонических - так называемые медленные нейромоторные единицы. Скелетные мышцы как правило, являются смешанными и поэтому при включении в работу разных мышц одна и та же мышца может выполнять либо динамическую деятельность за счет разных движений, осуществляемых быстрыми нейромоторными единицами, либо статическую деятельность за счет тонических сокращений, выполняемые медленными нейромоторными единицами.
3.Особенности проведения возбуждения в нервно-мышечном синапсе. Квантовая секреция медиатора. Механизм возникновения потенциала концевой пластинки.
Структура нервно-мышечного синапса представлена на рисунке. В процессе передачи возбуждения с нерва на мышечные волокна выделяют три последовательных процесса:
1. электрический, включащий достижение нервным импульсом концевой веточки аксона, деполяризацию и повышение проницаемости ее мембраны, выделение ацетилхолина (АХ) в синаптическую щель;
2. химический, основу которого составляет диффузия медиатора АХ к постсинаптической мембране и образование на ней его комплекса с холинорецептором; 3. электрический, включащий увеличение ионной проницаемости постсинаптической мембраны, возникновение локального электрического потенциала (потенциала концевой пластинки; ПКП), развитие потенциала действия мышечного волокна.
Запасов АХ в нервном окончании достаточно для проведения лишь примерно 10 000 импульсов. При длительной же импульсации мотонейрона, несмотря на постоянный синтез АХ, его содержание в концевых веточках может постепенно уменьшаться. В результате этого возможны нарушения передачи возбуждения в нервно-мышечных синапсах — пресинаптический нервно-мышечный блок.
Временно возникающий на постсинаптической мембране комплекс «АХ- рецептор» после прохождения каждого импульса разрушается ферментом ацетил-холинэстеразой. Однако при длительной высокочастотной импульсации мотонейрона (например при длительной и напряженной мышечной работе) АХ не успевает разрушаться и накапливается в синаптической щели. Способность постсинаптической мембраны к генерации ПКП при этом снижается и развивается частичный или полный постсинаптический нервно-мышечный блок, приводящий либо к частичному, либо даже полному прекращению развития потенциалов действия на мембране мышечного волокна.
1
— миелиновая оболочка
аксона;
2
— концевые веточки аксона;
3
— пузырьки, содержащие
ацетилхолин;
4
— митохондрия;
5
— пресинаптическая мембрана,
покрывающая концевую
веточку аксона в зоне нервно-мышечного
синапса;
6
— синаптическая щель;
7
— постсинаптическая мембрана,
покрывающая мышечное
волокно в зоне нервно-мышечного
синапса;
8
— ацетилхолинорецепторы на
постсинаптической мембране;
9
— митохондрия мышечного волокна;
10
— ядро мышечной клетки;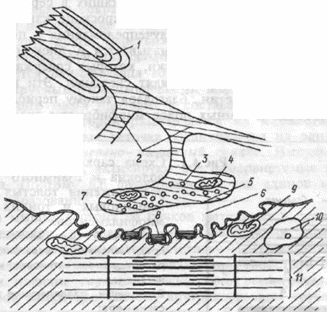
Высвобождение медиатора из синаптических пузырьков имеет квантовый характер. В состоянии покоя оно незначительно. При деполяризации пресинаптической мембраны под влиянием нервного импульса высвобождение медиатора резко усиливается. Взаимосвязь между деполяризацией пресинаптической мембраны и высвобождением медиатора выполняют ионы кальция.
В нервно-мышечном синапсе ацетилхолин синтезируется в окончанияхдвигательных нервов и накапливается в пузырьках. Когда в окончание приходит потенциал действия, ацетилхолин из 150-200 пузырьков высвобождается в синаптическую щель и связывается с холинорецепторами (холинорецепторы нервно-мышечных синапсов принадлежат к(N-холинорецепторам ), плотность которых особенно высока на гребнях складок постсинаптической мембраны. Каналы, сопряженные с холинорецепторами, открываются, в клетку входят катионы (в основном Na+), и происходит деполяризация постсинаптической мембраны, называемая потенциалом концевой пластинки. Поскольку этот потенциал в норме всегда сверхпороговый, он вызывает потенциал действия, распространяющийся по мышечному волокну и вызывающий сокращение. Потенциал концевой пластинки короткий, так как ацетилхолин, во-первых, быстро отсоединяется от рецепторов, во-вторых - гидролизуется АХЭ (ацетилхолинэстераза).