

Автореферат Демидова А.Т
..pdfТаблица 1 Видовой состав шмелей в зональных и азональных экосистемах
Среднеобской низменности
|
|
|
Северная |
Средняя |
Пойма |
Относительное обилие |
|
|
|
Виды шмелей |
шмелей в Среднеобской |
||||
|
|
тайга |
тайга |
р. Обь |
низменности в целом, |
||
|
|
|
|||||
|
|
|
|
|
|
B, баллы (Песенко, 1982) |
|
1. |
Bombus distinguendus |
+ |
+ |
+ |
4 |
(много) |
|
2. |
B.hortorum |
|
+ |
+ |
2 |
(мало) |
|
3. |
B.consobrinus |
+ |
+ |
+ |
2 |
(мало) |
|
4. |
B.muscorum |
|
|
+ |
3 |
(средне) |
|
5. |
B.pascuorum |
+ |
+ |
+ |
4 |
(много) |
|
6. |
B.schrencki |
|
+ |
+ |
3 |
(средне) |
|
7. |
B.rupestris |
|
|
+ |
2 |
(мало) |
|
8. |
B.campestris |
|
+ |
+ |
2 |
(мало) |
|
9. |
B.bohemicus |
+ |
+ |
+ |
4 |
(много) |
|
10. |
B.barbutellus |
|
|
+ |
1 |
(единично) |
|
11. |
B.flavidus |
+ |
+ |
+ |
4 |
(много) |
|
12. |
B.norvegicus |
+ |
+ |
+ |
2 |
(мало) |
|
13. |
B.quadricolor |
|
+ |
|
1 |
(единично) |
|
14. |
B.sylvestris |
+ |
+ |
+ |
3 |
(средне) |
|
15. |
B.lapponicus |
+ |
+ |
|
1 |
(мало) |
|
16. |
B.hypnorum |
+ |
+ |
+ |
4 |
(много) |
|
17. |
B.modestus |
|
+ |
|
1 |
(единично) |
|
18. |
B.pratorum |
|
+ |
+ |
3 |
(средне) |
|
19. |
B.jonellus |
+ |
+ |
+ |
4 |
(много) |
|
20. |
B.cingulatus |
+ |
|
|
1 |
(единично) |
|
21. |
B.balteatus |
+ |
|
|
1 |
(единично) |
|
22. |
B.sporadicus |
+ |
+ |
+ |
3 |
(средне) |
|
23. |
B.lucorum |
+ |
+ |
+ |
5 |
(очень много) |
|
24. |
B.sichelii |
|
+ |
+ |
3 |
(средне) |
|
25. |
B.cullumanus |
|
|
+ |
1 |
(единично) |
|
26. |
B.semenoviellus |
+ |
+ |
+ |
3 |
(средне) |
|
|
|
Итого |
15 |
20 |
21 |
|
|
Примечание: + – наличие шмелей в указанной экосистеме.
4.3. Зоогеографический анализ шмелей Среднеобской низменности
Для описания распространения видов шмелей Среднеобской низменности нами была использована классификация К. Б. Городкова (1984).
Зоогеографический анализ показал, что фауна шмелей изучаемой территории сформирована видами, имеющими ареалы пяти типов: трансголарктические, циркумполярные, транспалеарктические, субтранспалеарктические, европейско-сибирские.
Большинство видов шмелей имеет широкие зональные ареалы – температные и бореальные. По долготной составляющей преобладают транспалеарктические виды.
В северной тайге, в пределах исследуемого участка, особый вклад вносят арктобореальные и арктотемператные виды шмелей. Пойма р. Обь отличается присутствием суббореального B.cullumanus, отсутствием арктобореальных видов шмелей. В средней тайге доминируют бореальные и температные виды.
11
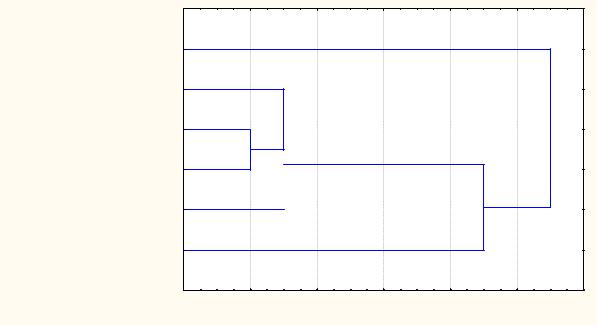
На основании собственных сборов и анализа литературных данных нами проведено сравнение фауны шмелей Среднеобской низменности с некоторыми районами (рис. 2).
Среднеобская низменность
Томское Приобье
Лесостепь, степь Западной Сибири
Южный Урал
Северо-запад гор Южной Сибири
Северная Европа
0,10 |
0,12 |
0,14 |
0,16 |
0,18 |
0,20 |
0,22 |
1-Ics
Рис. 2. Сходство фаун шмелей Среднеобской низменности с сопредельными территориями.
Примечание: Ics – коэффициент Чекановского-Съеренсена.
Среднеобская низменность – собственные исследования (Демидова 2008, 2010, 2011); Томское Приобье – Томская область (Конусова, 2000, 2005); Северная Европа – Архангельская область, республика Коми (Купчикова, 1960; Болотов, Подболоцкая, 2003; Колосова, 2005; Болотов, Колосова, 2007 и др.), Южный Урал – Челябинская область (Тюмасева, 2004; Красная книга Челябинской области, 2005; Резчикова, 2007), лесостепь, степь Западной Сибири – Курганская, Тюменская, Омская, Новосибирская области, Алтайский край (Лавров, 1927; Гребенников, 1972; Богатырев, 1985; Афиногенов, Золотарева, 1988; Бывальцев, 2008), северо-запад гор Южной Сибири – Кемеровская область (Еремеева, Лу-
зянин, 2008).
Фаунистический комплекс шмелей Среднеобской низменности сходен с Северной Европой за счет арктобореальных и арктотемператных видов с кластером, объединяющим условно южные районы (Южный Урал, лесостепь, степь Западной Сибири, Томское Приобье, северо-запада гор Южной Сибири) за счет бореальных и температных видов шмелей. Особенностью фауны шмелей Среднеобской низменности, в отличие от сравниваемых регионов, является наличие циркумполярного арктобореального вида
B.balteatus.
Глава 5. ЭКОЛОГО-БИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ШМЕЛЕЙ СРЕДНЕОБСКОЙ НИЗМЕННОСТИ
5.1. Биотопическое распределение шмелей
Для изучения биотопического распределения шмелей на территории Среднеобской низменности были обследованы четырнадцать групп биотопов. В зональной экосистеме средней тайги Западной Сибири в пределах Среднеобской низменности: смешанные и темнохвойные леса. В зональной
12
экосистеме северной тайги: светлохвойные и смешанные леса. В азональных экосистемах – четыре группы биотопов в пределах поймы реки Оби (светлохвойные, смешанные леса, ивняки и заливные луга) и два болотных биотопа (низинные и верховые болота). В антропогенных и антропогеннотрансформированных естественных экосистемах: вырубки смешанного леса, вторичные суходольные разнотравные луга, просеки среди темнохвойного леса, обочины дорог.
Наибольшим видовым разнообразием характеризуются смешанные леса средней тайги и поймы р.Обь, заливные луга (65% от общей фауны Среднеобской низменности). Самыми обедненными в видовом отношении являются низинные болота (27% фауны) и светлохвойные леса северной тайги
(35% фауны).
Для оценки структуры сообщества в исследуемых биотопах были вычислены индексы разнообразия Шеннона-Уивера Н', выравненности E и доминирования Симпсона DSm (рис. 3).
Небольшие показатели индексов разнообразия Н' (1,07) и выравненности E (0,47), высокие значения индекса доминирования DSm (0,56) шмелей в темнохвойных лесах средней тайги говорят о меньшем разнообразии. В данной группе биотопов доминирует, в основном, эвритопный вид B. lucorum, остальные виды шмелей встречались единично. Самые высокие показатели индекса разнообразия Н' наблюдаются в светлохвойных и смешанных лесах поймы р. Обь (2,22 и 2,10 соответственно) за счет относительно равномерного распределения эвритопных видов шмелей и увеличения лесных представителей. Вырубки светлохвойных лесов отличаются богатым разнообразием по этим же причинам. Кроме того, в данной группе биотопов появляются луговые шмели, предпочитающие открытые места обитания. Ивняки в пойме р.Обь имеют самый высокий показатель выравненности E (0,89), что, в первую очередь, говорит об устойчивости сообщества шмелей в данном биотопе. Вторичные суходольные луга и пустыри, обочины дорог отличаются высоким обилием шмелей при одинаковой методике сбора исследуемых объектов для всех биотопов. Однако из рисунка 3 наглядно видно, что луга имеют выше значения индексов разнообразия Н' (2,14) и выравненности E (0,84), ниже значения индекса доминирования DSm (0,15) по сравнению с обочинами (1,64; 0,64; 0,26 соответственно). Данный факт можно объяснить степенью антропогенной нагрузки двух сравниваемых биотопов: луга представлены более разнообразной растительностью, расположены среди леса, отличаются меньшим вытаптыванием почвенно-растительного покрова и т.д., напротив, пустыри и обочины дорог представлены, в основном, рудеральными растениями, обычно с нарушением структуры почвы, отличаются сильной антропогенной нагрузкой в виде шумов, химического загрязнения от выхлопов автотранспорта и т.п.
13
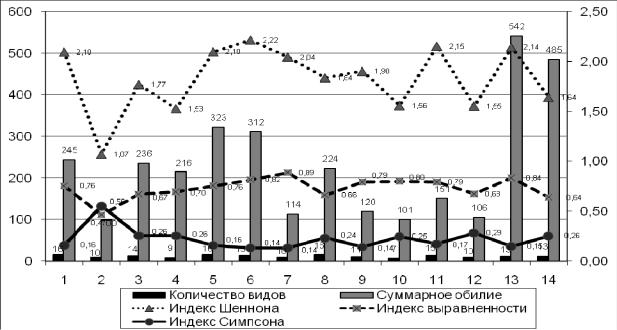
Рис. 3. Соотношение числа видов шмелей, обилия и индексов разнообразия Шеннона, выравненности и доминирования Симпсона в исследуемых биотопах.
Примечание: Средняя тайга: 1.Смешанные леса, 2.Темнохвойные леса; Северная тайга: 3.Смешанные леса, 4.Светлохвойные леса; Пойма р. Обь: 5.Смешанные леса, 6.Светлохвойные леса, 7.Ивняки, 8.Заливные луга; Болота: 9.Верховые, 10.Низинные; Антропогенные комплексы: 11.Вырубки светлохвойных лесов, 12.Просеки в темнохвойном лесу, 13.Вторичные суходольные луга, 14.Пустыри и обочины дорог.
Проанализировав данные с использованием индекса сходства Чеканов- ского-Съеренсена, пришли к выводу, что наиболее схож видовой состав шмелей светлохвойных лесов поймы р. Обь и вырубки светлохвойных лесов (преимущественно этот тип биотопов исследован в подзоне средней тайги). Сходство обусловлено наличием, в основном, видов-эврибионтов. Сходство фаун шмелей ивняков, заливных лугов и смешанных лесов поймы р. Обь объясняется месторасположением данных биотопов (близость друг другу, все три биотопа расположены в пойме р. Обь), схожими климатическими условиями (особенно увлажнение и температура), а также благоприятствующими условиями для распространения шмелей. Сходство фаун шмелей данных биотопов со вторичными суходольными лугами можно объяснить этими же причинами, так как данный тип биотопов, в основном, был изучен в пойменной части р. Обь (окрестности д. Сайгатина). Низинные болота имеют незначительное сходство с другими биотопами в связи с особенностью растительности, а также, потому что в данной группе биотопов встречается небольшое количество видов шмелей.
Главными фактами сходства видового разнообразия шмелей являются наличие тех же кормовых растений в биотопах и эврибионтность видов шмелей. Биотопическое распределение шмелей также обусловливается ланд- шафтно-климатическими условиями и степенью антропогенной нагрузки.
5.2. Основные экологические группы имаго
При выявлении экологических групп по степени приуроченности к местообитанию мы использовали традиционную классификацию Д.В. Панфи-
14
лова (1956, 1957, 1978), которой придерживаются исследователи пчелиных до настоящего времени. Согласно этой классификации выделяют три экологические группы: лесные, луговые и эвритопные. Анализ биотопического распределения шмелей в Среднеобской низменности позволил нам выявить некоторые отличия принадлежности изучаемых видов шмелей к той или иной экологической группе.
Установлено, что в северной тайге на водоразделах крупных рек отсутствуют луговые виды шмелей; в средней тайге и пойме р. Обь обнаружены как лесные и луговые виды шмелей, так и эвритопные; в средней тайге основу фауны шмелей составляют эвритопные виды, увеличивается доля лесных видов; в пойме р. Обь возрастает доля эвритопных и луговых видов шмелей, уменьшается доля лесных представителей.
Сравнивая фауны шмелей других регионов Западной Сибири, можно проследить следующие тенденции (Конусова, 2005; Бывальцев, 2008; Еремеева, Лузянин, 2008 и др.): в направлении с севера на юг возрастает роль луговых видов шмелей, уменьшается роль лесных; эвритопные виды встречаются повсеместно; в пойменных частях крупных рек увеличивается доля луговых видов шмелей.
5.3. Трофические связи шмелей
Нами исследованы трофические связи шмелей 14 видов Среднеобской низменности с растениями 83 видов из 68 родов, 25 семейств. При анализе пищевой специализации выяснено, что учтенные шмели отдавали предпочтение, в наибольшей степени, растениям семейств Бобовые
(Fabaceae) – 100% изученных шмелей, Розоцветные (Rosaceae) – 100%, и
Сложноцветные (Asteraceae) – 93%. Сходные тенденции отмечены в литературе (Тюмасева, 1988; Воробьева, Седин, 2005; Еремеева, Лузянин, 2008 и др.). Наименьшее видовое разнообразие шмелей наблюдали на растениях се-
мейств Маревые (Chenopodiaceae) – 7%, Гречишные (Polygonaceae) – 7%, Брусничные (Vacciniaceae) – 7%, и Крестоцветные (Brassicaceae) – 7%.
Анализ спектра посещаемых шмелями растений показал, что наибольшая ширина спектра (22–74 опыляемых видов растений) характерна для шмелей пяти видов (B.distinguendus, B.pascuorum, B.hypnorum, B.jonellus, B.lucorum), составляющих 80,4% от общего количества особей шмелей, зарегистрированных на территории Среднеобской низменности.
Шмели B.lucorum и B.pascuorum отличаются наибольшими показателями ширины трофической составляющей экологической ниши (H') – 3,98 и 3,12 соответственно, что является логичным, в связи с большим обилием этих насекомых. Широкая трофическая составляющая ниши говорит о пластичности данных видов шмелей по сравнению с другими видами. На основании сходства посещаемых растений шмелями разных видов нами оценена (с помощью индекса Жаккара – Ij) степень перекрывания трофических составляющих экологических ниш шмелей. Исследования показали, что перекрывание трофических составляющих ниш шмелей составляет от 28% до
61%.
Для шмелей нашего региона характерны два типа перекрывания трофических составляющих экологических ниш: ниша одного вида шмелей целиком включена в более обширную нишу другого вида; ниши частично перекрываются.
15
5.4. Особенности суточной и сезонной активности шмелей
5.4.1. Сезонная активность шмелей
Динамику сезонной активности шмелей изучали на протяжении трех лет – с апреля по октябрь 2008–2010 гг. на суходольном разнотравнозлаковом лугу в окрестностях деревни Сайгатина, расположенной на отрезке Сургутской поймы р. Оби.
Установлено, что в разные годы срок вылета имаго шмелей варьирует. В 2008 и 2009 году вылет приходился на первую декаду мая (7 и 8 мая соответственно) при температурах 11–14°С, единично шмели регистрировались в апреле (при температуре выше 10°С). В 2010 году шмели впервые были отмечены 24 апреля при температуре 13°С и до конца мая встречались спорадично, и только в первой декаде июня встречи стали постоянными.
Самки B.lucorum первыми появлялись после зимней диапаузы. Вылет приходился на третью декаду апреля – первую декаду мая, в зависимости от года. Стабильное их появление обычно наблюдалось в мае. После B.lucorum в начале мая вылетали B.pascuorum, а позже и шмели других видов. Наиболее продолжительный лёт наблюдался у B.lucorum, B.pascuorum, B.sporadicus.
Нами выявлено, что обилие шмелей в течение лётного периода зависит от суммы эффективных температур в весенние месяцы – времени, когда вылетают самки после зимней диапаузы (коэффициент корреляции 0,99 (p=0,05)). Суммарное обилие шмелей от суммы эффективных температур в весенне-летне-осенние месяцы зависит в меньшей степени (коэффициент корреляции 0,50 (p=0,05)).
Подробно фенологические процессы и динамика сезонной активности нами были изучены для массового вида B. lucorum. Закладка гнезда B. lucorum предположительно приходилась на вторую, третью декады мая, именно в это время появлялись самки с обножками. Первые рабочие особи отмечались в третьей декаде мая – первой декаде июня. Массовое их фуражирование приходилось на третью декаду июня – первую декаду июля. Молодые самки и самцы появлялись в августе. К концу сентября наблюдался спад обилия шмелей. Изучив динамику сезонной активности B.lucorum на исследуемой территории, выявили задержку в фенологических процессах по сравнению с южными районами на две недели (лесостепь Западной Сибири, Южный Урал).
5.4.2. Суточная активность шмелей
Характер суточной активности шмелей B.distinguendus, B.lucorum, изучали в двух локалитетах – проселочная дорога среди смешанного леса в окрестностях поселка Салым Нефтеюганского района (2007–2008 гг.), разнотравный луг с антропогенной нагрузкой (вытаптывание, выпас скота) в окрестностях деревни Сайгатина (2008–2009 гг.). Изменение активности шмелей в течение суток является видоспецифичным показателем.
Наибольшая активность шмелей вида B.distinguendus приходится в утреннее время с 10.00 до 11.00 и в вечернее с 16.00 до 19.00. Ранним утром в 6.00 активность низкая, также и в обеденное время. Шмели-чесальщики B.distinguendus относятся к насекомым с утренне-вечерним ритмом активности. При исследовании зависимости активности шмелей от температуры воздуха нами подтверждено, что интенсивность работы опылителей зависит от
16
температуры. Лимитирующими факторами активности фуражирования шмелей являются как невысокие температуры воздуха – меньше 15°С, так и слишком высокие – свыше 25°С. Оптимальная температура для B.distinguendus в районе исследования 19–24 ºС.
Шмели B.lucorum почти равномерно активны в течение светового дня, небольшой пик активности наблюдается в утреннее время в 9.00-10.00 часов. Равномерность активности B.lucorum в течение дня хорошо видна в зависимости от показателей температуры. Наибольшая активность его наблюдается при температурах от 19ºC до 23ºC. Однако при температуре выше +23ºС происходит некоторый спад активности этих насекомых.
5.5.Влияние экологических факторов на жизнедеятельность шмелей
Внастоящей главе показано влияние экологических факторов на видо-
вое разнообразие и обилие шмелей, обитающих в окрестностях археологического памятника «Барсова гора», на границе санитарно-защитной зоны полигона твердых и бытовых отходов (ЗАО «Полигон ЛТД»), около факела сжигания газа (ЗСК) на расстоянии 10–40 высот факела (предполагаемая зона распыления), в окрестностях парка отдыха «Орбита» г. Сургута в придорожной зоне. Памятник «Барсова гора» приняли как «условно чистый», в виде контроля.
Окрестности памятника «Барсова гора» и придорожная зона около парка отдыха «Орбита» характеризуются высоким разнообразием шмелей (индекс H' 1,68 и 1,90 соответственно) и устойчивостью сообщества (индекс E 0,73 и 0,74 соответственно, индекс DSm 0,29 и 0,21 соответственно).
Для объяснения структуры сообщества этих насекомых в исследуемых биотопах нами были рассмотрены основные параметры, способные повлиять на деятельность шмелей (загрязнение окружающей среды, климатические условия, наличие кормовых ресурсов и др.).
Содержание тяжелых металлов (мышьяка As, свинца Pb, кадмия Cd) в почве, соцветиях и шмелях неодинаково. Проба почвы с территории «Барсова гора» характеризуется меньшим количеством Pb, а в придорожной зоне около парка отдыха «Орбита» – меньшим количеством As. Почва всех четырех исследуемых биотопов содержит незначительное количество Cd – менее 0,5мг/кг. В соцветиях растений количество Pb и As больше в районе факела, а содержание Cd немного превышает в районе полигона. Большее превышение As и Pb наблюдается у шмелей, собранных около дороги в районе Орбиты. В соцветиях растений исследуемых биотопов содержание микроэлементов в сравнении с почвой значительно уменьшается: Cd в 125–500 раз, As в 147– 350 раз, Pb в 3–13 раз. Количество Cd и Pb в теле шмелей по сравнению с соцветиями растений возрастает: Cd в 2–12 раз, Pb в 2–4 раза. Количество As варьирует: в шмелях меньше, чем в соцветиях растений около факела и на Барсовой горе; незначительно увеличивается около полигона и в 4 раза превышает около автодороги.
Анализ проведенных исследований, на предмет соотношения видового разнообразия шмелей и накопления в их теле тяжелых металлов в условиях экологически неблагоприятных территорий, показал прямо пропорциональную зависимость. Возможно, этот факт объясняется высокими адаптационными способностями этих организмов, которые сформировались в процессе сопряженной эволюции. Так, коэффициент корреляции индекса разнообразия
17
H' и содержания Cd, Pb и As в шмелях высокий: 0,73 (p=0,05), 0,74 (p=0,05) и 0,71 (p=0,05) соответственно. При изучении влияния тяжелых металлов – Pb и As, аккумулированных в шмелях, на обилие этих насекомых не выявлена закономерность (коэффициент детерминации R2сумм.обилие=0,125, 0,010 соответственно). При увеличении Cd в шмелях наблюдается незначительное повышение обилия их (R2сумм.обилие=0,328).
Однако потенциальную угрозу на живые организмы несет в себе загрязнение атмосферного воздуха. Нами изучено влияние концентрации в воздухе диоксида серы (SO2) и оксида углерода (CO). Загрязненность атмосферы SO2 выше на полигоне, чем около факела и придорожной зоне, но на этих территориях наблюдается высокая концентрация CO. Нами установлено, что видовое разнообразие шмелей резко снижается в биотопах, где загрязнение воздуха CO превышает фоновый уровень (коэффициент корреляции равен -0,93 (p=0,05)), а загрязнение SO2 также оказывает отрицательное воздействие, но умеренное (коэффициент корреляции равен -0,31 (p=0,05)). Кроме того, исходя из показателей коэффициента детерминации, видно, что концентрация загрязняющих веществ в атмосферном воздухе в большей степени влияет на обилие шмелей, чем на видовое разнообразие их
(R2индекс Шеннона=0,022 (CO), 0,082 (SO2); R2сумм.обилие=0,593 (CO), 0,424 (SO2)).
Актуальным вопросом является изучение влияния радиактивности окружающей среды на организм. Нами проведены измерения гаммаизлучения в исследуемых биотопах. Выявленные уровни гамма-излучения не превышают допустимых норм (0,06–0,13 мЗв). Установлена обратная заметная связь между разнообразием шмелей в исследуемых биотопах и мощностью дозы гамма-излучения – коэффициент корреляции равен -0,66 (p=0,05). Исходя из показателей коэффициента детерминации, видно, что: при увеличении мощности гамма-излучения уменьшается разнообразие шмелей (R2индекс Шеннона=0,364); колебания показателей мощности гамма-излучения в диапазонах 0,06–0,13 мЗв не влияют на обилие шмелей (R2сумм.обилие=0,084).
Среди абиотических факторов также были исследованы температура, давление, влажность, скорость ветра и освещение. Кроме того, изучены некоторые биотические факторы – проективное покрытие биотопов цветковыми растениями и наличие клещей (семейство Parasitidae и др.) на шмелях. В результате факторного анализа выявили: видовое разнообразие шмелей в различных биотопах зависит, в первую очередь, от температуры, силы ветра, освещения; при увеличении площади покрытия территории энтомофильными растениями, возрастает видовое разнообразие шмелей (корреляция заметная, составляет 0,64); паразитические клещи оказывают отрицательное воздействие на видовое разнообразие шмелей (коэффициент корреляции умеренный, составляет -0,37); при увеличении обилия шмелей возрастает пораженность их клещами (коэффициент корреляции заметный, составляет 0,63); обилие зависит, в большей степени, от силы ветра – чем больше сила ветра, тем обилие на данном участке в данное время уменьшается (корреляция высокая, составляет -0,88).
Глава 6. ПРАКТИЧЕСКАЯ ЗНАЧИМОСТЬ ШМЕЛЕЙ
В настоящей главе дан анализ хозяйственного значения шмелей как эффективных опылителей многих растений. Приведены рекомендации по охране шмелей Ханты-Мансийского автономного округа – Югры.
18
ВЫВОДЫ
1. Впервые на территории Среднеобской низменности обнаружены 26 видов шмелей рода Bombus Latr. трибы Bombini. Наибольшим количеством видов исследуемого региона представлены подроды шмелей-кукушек Psithyrus Lepeletier (30,8% от фауны Среднеобской низменности) и шмелей Pyrobombus Dalla Torre (23,1%). Массовыми видами являются: B.lucorum,
B.pascuorum, B.hypnorum, B.jonellus, B.distinguendus, B.bohemicus, B.flavidus.
2. Фауна шмелей Среднеобской низменности сформирована видами, имеющими ареалы пяти типов: трансголарктические, циркумполярные, транспалеарктические, субтранспалеарктические, европейско-сибирские. Большинство видов шмелей имеет широкие зональные ареалы – температные
ибореальные. По долготной составляющей преобладают транспалеарктические виды.
3.Из четырнадцати изученных групп биотопов зональных, азональных
иантропогенных экосистем Среднеобской низменности наибольшим видовым разнообразием отличаются смешанные леса средней тайги и поймы
р.Обь, заливные луга (65% от фауны шмелей изучаемого региона). Самыми обедненными в видовом отношении являются низинные болота (27% фауны)
исветлохвойные леса северной тайги (35% фауны).
4.Особенности экологических групп шмелей по местообитанию терри-
тории Среднеобской низменности заключаются в следующем: в северной тайге на водоразделах крупных рек отсутствуют луговые виды шмелей; в средней тайге и пойме р. Обь обнаружены как лесные и луговые виды шмелей, так и эвритопные; в средней тайге основу фауны шмелей составляют эвритопные виды, увеличивается доля лесных видов; в пойме р. Обь возрастает доля эвритопных и луговых видов шмелей, уменьшается доля лесных представителей.
5. Шмели четырнадцати видов в условиях Среднеобской низменности трофически связаны с покрытосеменными растениями 83 видов, относящимися к 68 родам из 25 семейств. Шмели отдают предпочтение растениям семейств Бобовые (Fabaceae) – 100% изученных шмелей, Розоцветные (Rosaceae) – 100% и Сложноцветные (Asteraceae) – 93%. Наибольшая ширина спектра питания (22–74 опыляемых видов растений) характерна для шмелей
B.distinguendus, B.pascuorum, B.hypnorum, B.jonellus, B.lucorum, составляю-
щих 80,4% от общего количества особей шмелей, зарегистрированных на территории Среднеобской низменности. Шмели B.lucorum и B.pascuorum отличаются наибольшими показателями ширины трофической составляющей экологической ниши.
6. Сезонная активность шмелей составляет 157–170 дней. Массовое появление шмелей наблюдается в третьей декаде мая. Последние шмели встречаются в третьей декаде сентября, а у B.lucorum лёт продолжается до первой декады октября. Задержка в фенологических процессах, по сравнению с южными районами, составляет две недели. Суммарное обилие шмелей в течение лётного периода зависит от суммы эффективных температур в весенние месяцы (коэффициент корреляции 0,99 (p=0,05)). Суммарное обилие шмелей от суммы эффективных температур в весенне-летне-осенние месяцы зависит в меньшей степени (коэффициент корреляции 0,50 (p=0,05)).
7. Общая продолжительность суточной активности шмелей в начале лёта составляет 11–12 часов. В период массового лёта шмелей продолжи-
19
тельность увеличивается до 18–19 часов. Суточный ритм активности B.distinguendus имеет вид двухвершинной кривой, а для B.lucorum характерна одновершинная кривая. Максимальная активность шмелей отмечена нами при температуре +19°С – +25°С. Нижним порогом активности шмелей в ис-
следованном регионе является температура +15°С.
8. Наибольшее воздействие на видовое разнообразие шмелей оказывают: климатические условия (температура, сила ветра, освещение), кормовая база, мощность гамма-излучения окружающей среды и тяжелые металлы (As, Сd, Pb) в соцветиях растений и почве; на обилие шмелей: сила ветра, оксид углерода (CO) и диоксид серы (SO2) в атмосферном воздухе, зараженность шмелей клещами и тяжелые металлы (Сd) в соцветиях растений и почве.
Список работ, опубликованных по теме диссертации Работы, опубликованные по теме диссертации в издании, рекомен-
дованном ВАК
1. Демидова А.Т. Сравнительный анализ биотопического распределения и относительного обилия шмелей (Hymenoptera, Apidae, Bombus Latr.) зональных, азональных и антропогенных экосистем Среднеобской низменности // Вестник Алтайского государственного аграрного университета. Барна-
ул. 2011. №4 (78). С. 48–54.
2. Демидова А.Т., Тюмасева З.И. Фаунистический и зоогеографический анализ шмелей (Hymenoptera, Apidae, Bombus Latr.) Среднеобской низменности // Вестник Саратовского госагроуниверситета им. Н.И. Вавилова.
Саратов. 2011. №10. С. 14–17.
Список работ, опубликованных по теме диссертации в других изданиях
3. Хамматова А.Т., Тюмасева З.И. Биотопическое распределение шмелей (Hymenoptera, Bombus) средней тайги Западной Сибири (в пределах ХМАО) // Труды Института биоресурсов и прикладной экологии: материалы IV международной конференции «Биоразнообразие и биоресурсы Урала и сопредельных территории»; отв. ред. З.Н. Рябинина; Мин-во образования и науки РФ, Федер. агентство по образованию, Оренб. гос. пед. ун-т. Оренбург. 2008. С. 230–231.
4. Хамматова А.Т., Тюмасева З.И. Эколого-фаунистический обзор шмелей (Hymenoptera, Bombus) Шапшинского Урочища (природный парк Самаровский Чугас) // Биоразнообразие: проблемы и перспективы сохранения: материалы международной научной конференции, посвященной 135летию со дня рождения И.И. Спрыгина; ПГПУ им. В.Г. Белинского. Пенза. 2008. Часть II. С. 297–298.
5. Тюмасева З.И., Хамматова А.Т. Эколого-фаунистический обзор шмелей (Hymenoptera, Bombus) средней тайги Западной Сибири // Биологическая наука и образование в педагогических вузах: материалы пятой Всероссийской научно-практической конференции «Проблемы биологической науки и образования в педагогических вузах». Новосибирск. 2008. Вып. 5.
С.79–83.
6. Хамматова А.Т., Тюмасева З.И. Фауна и экологические аспекты шмелей (Bombus) Шапшинского урочища (природный парк «Самаровский чугас») // Природный парк «Самаровский чугас»: итоги и перспективы исследований: сб. научных трудов. Екатеринбург: ПП «Самаровский чугас», Урал. гос. лесотехн.ун-т, 2008. Вып. 1. С. 85–92.
7. Демидова А.Т., Тюмасева З.И. Видовой состав и биотопическое распределение шмелей ХМАО // Сборник научных трудов биологического фа-
20