

29
римыми в воде, свободно всасываются в кишечнике и поступают в кровь воротной вены, оттуда в печень, минуя какие-либо превращения в кишечной стенке.
Всасывание жирных кислот с длинной цепью и моноглицеролов происходит более сложно. Этот процесс осуществляется при участии желчи и, главным образом, желчных кислот, фосфолипидов, холестерина, входящих в ее состав. Жирные кислоты с длинной цепью и моноглицеролы в просвете кишечника образуют с этими соединениями устойчивые в водной среде мицеллы. Структура мицелл такова, что их гидрофобное ядро (жирные кислоты, моноглицеролы и т.д.) оказывается окруженным снаружи гидрофильной оболочкой из желчных кислот и фосфолипидов.
В составе таких мицелл высшие жирные кислоты переносятся к всасывающей поверхности кишечного эпителия. В результате диффузии, мицеллы проникают в эпителиальные клетки ворсинок, где происходит распад жировых мицелл. При этом желчные кислоты поступают в ток крови и через систему воротной вены попадают сначала в печень, а оттуда вновь в желчь. Таким образом происходит постоянная циркуляция желчных кислот между печенью и кишечником. Этот процесс получил название печеночно-кишечной циркуляции.
После того как произошло всасывание глицерина жирных кислот, они могут использоваться тканями и органами тела в качестве энергетического материала.
2.2 Окисление жира
Образовавшиеся в результате гидролиза триацилглицерола (см. рис. 7) глицерин и жирные кислоты вовлекаются в дальнейший путь распада – окисление.
2.2.1 Окисление глицерина
Глицерин вовлекается в цитоплазме клетке в процесс гликолиза.
Сначала глицерин при участии фермента глицерофосфаткиназы (трансфераза) превращается в α-глицеролфосфат. Последний под действием НАД+ – зависи-
мой α-глицеролфосфатдегидрогеназы (оксидоредуктаза) превращается в фосфодиоксиацетон:
СН |
ОН АТФ |
АДФ |
СН2 |
ОН |
НАД+ |
НАДН + Н+ |
СН2 |
ОН |
2 |
|
|
|
|
|
|
|
|
ОН НС |
трансфераза |
ОН НС |
|
дегидрогеназа |
С |
O |
||
|
|
|
||||||
СН2 |
ОН |
|
α СН2 |
O |
(Р) |
|
СН2 |
O (Р) |
глицерол |
|
α -глицеролфосфат |
фосфодиоксиацетон |
|||||
Фосфодиоксиацетон, являясь обычным метаболитом гликолиза, изомеризуется в 3-фосфоглицериновый альдегид и включается в данный метаболический путь до ПВК. Пировиноградная кислота, превращаясь в ацетил-КоА (этап III), полностью окисляется в цикле Кребса (этап IV) до СО2, Н2О и выделением АТФ.
2.2.2 Окисление жирных кислот
Окисление жирных кислот протекает в печени, почках, скелетных и сердечных мышцах, в жировой ткани.

30
Ф.Кнооп высказал предположение, что окисление молекулы жирной кислоты в тканях организма происходит в β-окислении. В результате от молекулы жирной кислоты отщепляются двууглеродные фрагменты со стороны карбоксильной группы. Процесс β-окисления жирных кислот складывается из следующих этапов:
Активация жирных кислот. Подобно первой стадии гликолиза сахаров перед β-окислением жирные кислоты подвергаются активации. Эта реакция протекает на наружной поверхности мембраны митохондрий при участии АТФ, коэнзима А (НS-КоА) и ионов Mg2+. Реакция катализируется ацил-КоА-синтетазой:
|
|
|
|
|
|
|
β |
|
|
α |
|
COОН + |
АТФ + НS-КоА |
Mg2+ |
|||||||||
R |
|
(СН2)n |
|
|
CH2 |
|
CH2 |
|
|
|
|||||||||||||
|
|
|
|
|
|
ацетил-КоА-синтнтаза |
|||||||||||||||||
|
|
|
жирная кислота |
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
O |
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β |
|
α |
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C S |
|
КоА + АМФ + РР |
||||
|
|
|
|
|
R |
|
(СН |
)n |
|
|
CH |
|
CH |
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
2 |
|
|
|
2 |
2 |
|
|
|
|
|
||||
ацил-КоА
В результате реакции образуется ацил-КоА, являющийся активной формой жирной кислоты.
Транспорт жирных кислот внутрь митохондрий. Коэнзимная форма жирной кислоты, в равной мере как и свободные жирные кислоты, не обладает способностью проникать внутрь митохондрий, где, собственно, и протекает их окисление, переносчиком активированных жирных кислот через внутреннюю митохондриальную мембрану служит карнитин (γ-триметиламино-β-оксибути-рат):
|
|
|
|
|
β |
|
α |
|
O |
|
|
|
|
|
|
|
|
|
|
CH3 |
|
|
|
|
|
|
|
|
|
|
|
цитоплазматическая |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
+ H3C |
|
|
|
+ |
|
|
|
|
|
|
|
|
|
|
|
COOH |
|
|
||||||||||||
R |
|
(СН2)n |
|
CH2 |
|
CH2 |
|
|
|
|
C |
|
|
|
|
|
N |
|
СН2 |
|
СН |
|
СН2 |
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
S |
КоА |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
карнитин-ацилтранс- |
|||||
|
|
|
|
ацил-КоА |
|
|
|
|
|
|
CH3 |
|
|
|
ОН |
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
фераза |
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
карнитин |
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
CH3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
НS-КоА |
+ H3C |
|
N |
+ |
СН2 |
|
СН |
|
СН2 |
|
|
COOH |
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α |
|
β |
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
CH3 |
|
|
|
|
|
|
C |
|
|
|
|
|
|
(СН2)n |
|
|
R |
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
ОН |
|
|
|
CH2 |
|
CH2 |
|
|
|
|||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
O
ацил-карнитин (в цитоплазме)
После прохождения ацилкарнитина через мембрану митохондрий происходит обратная реакция – расщепления ацилкарнитина при участии НS-КоА и митохондриальной карнитин-ацилтрансферазы:
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
31 |
|
|
||
|
|
|
CH3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
H3C |
|
N |
+ |
СН2 |
|
СН |
|
СН2 |
|
|
COOH |
+ |
|
НS-КоА |
|||||||||||||||
|
|
|
|
|
|
||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
α |
|
β |
|
|
|
|
|
||||
|
|
CH3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
ОН |
|
C |
|
(СН2)n |
|
|
R |
||||||||||||||||||
|
|
|
|
|
|
|
|
|
CH2 |
|
|
CH2 |
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ацил-карнитин (в митохондрии) |
|
|
|||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
β |
α |
|
O |
+ |
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C |
||||||||||||
|
|
|
|
|
|
R |
|
|
(СН2)n |
|
|
CH2 |
|
CH2 |
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
ацил-КоА в митохондрии |
|
S |
КоА |
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||
митохондриальная
карнитин-ацилтранс- фераза
CH3
H3C N+ СН2 СН СН2 COOH CH3 ОН
карнитин
Ацил-КоА в митохондрии подвергается процессу β-окисления.
Этот путь окисления связан с присоединением атома кислорода к углеродному атому жирной кислоты, находящемуся в β-положении:
O
RCH2 CH2 C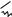


 SKoA
SKoA
При β-окислении происходит последовательное отщепление от карбоксильного конца углеродной цепи жирной кислоты двууглеродных фрагментов в форме ацетила-КоА и соответствующее укорачивание цепи жирной кислоты:
|
|
O |
С С С С |
С C |
SKoA |
|
|
В матриксе митохондрии ацил-КоА распадается в результате повторяющейся последовательности четырех реакций (рис.8).
1)окисление с участием ацил-КоА-дегидрогеназы (ФАД-зависимой дегидрогеназы);
2)гидратация, катализируемой еноил-КоА-гидратазой;
3)второго окисления под действием 3-гидроксиацетил-КоА-дегидрогеназы (НАД-зависимой дегидрогеназы);
4)тиолиза с участием ацетил-КоА-ацилтрансферазы.
Совокупность этих четырех последовательностей реакций составляет один оборот β-окисления жирной кислоты (см. рис. 8).
Образовавшийся ацетил-КоА подвергается окислению в цикле Кребса, а ацетил-КоА, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил-КоА (4-углеродное соединение), на последнем этапе β-окисления распадается на две молекулы ацетилКоА.
При окислении жирной кислоты, содержащей n углеродных атомов, происходит n/2-1 цикл β-окисления (т.е. на один цикл меньше, чем n/2, так как при окислении бутирил-КоА сразу происходит образование двух молекул ацетилКоА) и всего получится n/2 молекул ацетил-КоА.
Например при окислении пальмитиновой кислоты (С16) повторяется 16/2- 1=7 циклов β-окисления и образуется 16/2=8 молекул ацетил-КоА.

|
|
|
|
|
|
|
|
32 |
|
|
|
|
|
|
β |
α |
О |
||||
R |
|
(СН2)n |
|
СН2 |
СН2 |
|
С |
S |
|
КоА |
|
|
|
|
|||||||
ацил-КоА
ФАД
Окисление
(1)
ФАДН2
НН О
R(СН2)n С С С S КоА
С С S КоА
енолил-КоА (транс-форма)
Н2О |
Гидратация (2) |
|
|
|
ОН Н О |
R (СН2)n С С С


 S КоА
S КоА
НН
L-гидроксиацил-КоА НАД+
Второе окисление (3)
НАДН + Н+
ОО
R (СН2)n С СН2 С



 S КоА
S КоА
Оксоацетил-КоА НS - КоА
|
|
Тиолиз (4) |
|
|
|
|
О |
|
|
|
|
|
|
СН3 С |
S КоА |
О |
Ацетил-КоА |
|
|
|
R |
(СН2)n |
|
|
|||
С |
КоА |
О |
|
|
||
|
|
S |
|
|
||
|
|
|
СН3 |
С |
S КоА |
|
|
|
|
|
О |
|
ЦТК |
|
|
|
СН3 |
С |
S КоА |
|
|
|
|
|
|||
|
|
|
|
О |
|
|
|
β |
α |
СН3 |
С |
S КоА |
|
|
О |
|
|
|
||
|
H3C СН2 |
СН2 |
С |
|
|
|
|
бутирил-КоА S КоА |
|
|
|
||
|
|
β − |
окисление (последний этап) |
|||
О |
|
|
О |
|
|
|
СН3 С S КоА |
|
|
СН3 С |
S |
КоА |
ЦТК |
Ацетил-КоА
Рисунок 8 – Схема β-окисления жирной кислоты