

Гш-
10. Провідні гуморальні і клітинні
фактори в розвитку противірусного
Імунітету: Тс - T-
супресор,
Тх- T-хелпер,
Тц- цитотоксичний Т-лімфоцит,
І
‘
унресорні фактори, 2,3,4 - хелперні фактори,
5 - презентація антигену, 6,7 - рі|н-м орні
медіатори, 8-антитіла; пунктирна лінія
- для розчинних факторів; лінія -
безпосередня дія клітин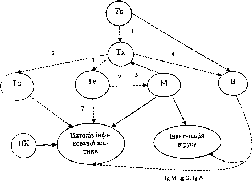
І «• і сфектор гст, м- макрофаг, в-в-клітина, пк-природний кілер,
\ 111 и і іла, які утворюються при вірусних інфекціях, діють безпосе- !«■ ни м на вірус або на клітини, інфіковані вірусом. У зв’язку з цим Ми.».н,і пнділити дві основні форми участі антитіл у розвитку про- ІИИІр и IIIII о імунітету.
Одна з них - нейтралізація вірусу антитілами. Така нейтралізація ' перешкоджає рецепції вірусу на клітині і проникненню його в | клітину. Ефект нейтралізації посилюється у присутності Ко-фактора або комплементу, а також антиідіотипових антитіл, що з’являються на пізніх стадіях інфекції і зв’язують імуноглобулінові епітопи комплексу, який складається з вірусних часток та антитіл. Опсонізація віріонів антитілами сприяє фагоцитозу. Комплекс, який складається з вірусних часток та імуноглобуліну, зв’язується з поверхнею макрофага за рахунок його Рс-рецепторів. Інтеріалізація комплексу у фагоцитарній вакуолі, як правило, призводить до загибелі збудника.
Інша форма участі антитіл - імунний лізис інфікованих клітин. Можливі два варіанти такої цитотоксичності. Комплементозалежна цитотоксичність виникає при дії антитіл на антигени, експресійовані на поверхні інфікованої клітини, з наступною активацією системи комплементу. В іншому випадку взаємодія інфікованої клітини з ан- І титілами класу \% в виявляється недостатньою для загибелі клітин- І мішеней. Цитотоксичність виникає, якщо клітини-мішені додатково І контактують з клітинами, які несуть рецептори до Бс-фрагментів ^ в. Такими клітинами є О-лімфоцити (це не Т- і не В-клітини), по- ліморфноядерні лейкоцити і макрофаги, які не володіють специфічністю щодо вірусного антигену.
Основну масу противірусних антитіл складають І§ в. Антитіла класу ^ М свідчать про нещодавно перенесену інфекцію, вони з’являються раніше від інших і раніше зникають порівняно з ^ 0« антитілами. За багатьох ентеровірусних і ротавірусних інфекцій важ* ливого значення набуває місцевий імунітет, пов’язаний з утворенням і секрецією антитіл класу 1% А. ^
Гуморальний імунітет бере участь у розвитку противірусної стіЙ» кості при інфекціях, спричинених арбо-, ентеро-, риновірусами. ЛНі титіла нейтралізують вірус після загибелі клітин-мішеней. ДоведсИІ зворотна залежність між захворюваністю деякими інфекціями і ТИ1* рами відповідних противірусних антитіл. Про значення циркулюючі^ антитіл у противірусному імунітеті свідчить великий практичний Дв* свід застосування імуноглобулінів для пасивної імунізації людей про* ти кору, краснухи, гепатиту А і В тощо.
При інших вірусних захворюваннях антитіла є лише свідками Іму< нної відповіді на вірус. Деякі віруси, наприклад, аденовіруси, 11^ песвіруси, можуть протягом тривалого часу персистувати в оргмиїїМ)
їа наявності антитіл. Навіть високий рівень антитіл не виключає тривалий перебіг деяких вірусних інфекцій. При недостатній концентрації антитіл може виникати феномен посилення вірусної репродукції. Антитіла можуть захищати вірус від впливу протеолітичних ферментів клітини, що (при умові збереження життєздатності вірусу) приводить до посилення реплікації вірусу.
Вірусонейтралізуючі антитіла діють безпосередньо на вірус лише і оді, коли вірус, зруйнувавши одну клітину, розповсюджується на ніші. Деякі віруси, наприклад віруси родини герпес переходять із кліпі ни в клітину по цитоплазматичних містках і уникають дії циркулюючих антитіл. У цьому випадку і у випадку інтегрованої форми вірусної інфекції, коли вірусний геном інтегрується з ДНК-чутливих клітин, провідну роль у становленні імунітету відіграють клітинні механізми, пов’язані передусім з дією специфічних цитотоксичних Т- имфоцитів, Т-ефекторів ГСТ і макрофагів. Першочергове значення у противірусному імунітеті має процес формування специфічних цито- юксичних Т-лімфоцитів. Попередник таких лімфоцитів активується з одного боку комплексом вірусного антигену з продуктами генів гі- і юсумісності класу І, а з іншого боку - розчинними медіаторами Т- м-11 перів. Цитотоксичні лімфоцити, які утворюються, здатні лізувати і« ні піни, інфіковані вірусом, реагуючи таким чином на вірусний анти- иті, асоційований з антигенами гістосумісності класу І.
Дня цитотоксичної дії Т-лімфоцитів на клітини-мішені необхідний і іг (посередній контакт клітин, після чого відбувається зміна мембранної проникності клітини-мішені, її осмотичне набрякання, розрив мем- гцшііи і вихід вмісту цитоплазми в навколишнє середовище. Ймовірно, мін механізм цитотоксичного ефекту пов’язаний з активацією мем- !і|шііпнх ферментних систем на ділянках прилипання клітин, утворен- пмм цитоплазматичних містків між клітинами і впливом лімфотоксину.
( пецифічні Т-кілери з’являються на ранніх стадіях (через 1 - 3 доїш теля зараження), їхня активність досягає максимуму через тижні ні. і повільно знижується протягом декількох тижнів, хоча клітини ним чи цитотоксичних лімфоцитів зберігаються протягом тривалого •♦ні \ І аким чином, утворення цитотоксичних Т-лімфоцитів є ранні"!" імунною відповіддю, яка нерідко передує утворенню циркулю- *ч'інч .ни тіл.
і -.її« ву роль у противірусному набутому імунітеті відіграють Т- і і "|ні ГСТ. Ці клітини розпізнають вірусний антиген в асоціації з
антигенами гістосумісності переважно класу II і виділяють медіатори клітинного імунітету. Серед медіаторів особливе значення має лімфо- токсин, який викликає загибель інфікованих клітин-мішеней, і медіатори, що активують функцію макрофагів. Роль специфічних клітинних факторів є особливо важливою при інфекціях, коли вірус недосяжний для її антитіл. Віруси, які надходять у міжклітинну рідину, нейтралізуються за допомогою антитіл, (3-інгібіторів і температурної денатурації. Макрофаги фагоцитують інфіковані вірусом живі і зруйновані клітини. Одним із факторів противірусного імунітету є інтерферон, що утворюється здебільшого у місцях розмноження вірусу. Інтерферон не впливає на вірус безпосередньо, він посилює експресію антигенів гістосумісності, викликає специфічне гальмування транскрипції вірусного геному і специфічне придушення трансляції вірусної мРНК, яка перешкоджає накопиченню вірусу в клітині. Що стосується властивостей інтерферону, то він, на відміну від відомих інгібіторів, які діють на віруси безпосередньо, впливає на них через чутливі клітини, надаючи їм стійкості до вірусної репродукції. Найбільш важливою ознакою інтерферону, яка відрізняє його від інших захис* них факторів, є внутрішньоклітинний характер дії. Тому, щоб вважа* ти інгібітор інтерфероном, слід переконатися в його здатності стиму* лювати резистентність чутливих клітин до різних вірусів. Щоб інтер» ферон міг проявити антивірусну дію, чутливі клітини повинні мати непорушений синтез клітинної РНК і клітинних білків.
У процесі непримиренної боротьби з імунною системою у вірусі!? постійно змінюється структура поверхневих антигенів. Ці процеси звано “антигенним дрейфом” і “антигенним шифтом”. Щоб зрозуміТЙ суть цього явища, доцільно розглянути його на прикладі різних штаміі вірусу грипу. На поверхні цього збудника розташований гемаглютені за допомогою якого вірус прикріплюється до клітини перед тим, проникнути всередину, а вивільнення новоутворених вірусних час’ від поверхневих сіалових кислот зараженої клітини забезпечує ней мінідаза. Для створення протективного імунітету гемаглютенін біл' важливий, ніж нейрамінідаза. Мінорні поступові зміни антигенних п( стивостей гемаглютеніну відбуваються в результаті точкових мугді вірусного геному (дрейф) тоді, коли значні зміни виникають впаслі обміну генетичним матеріалом з іншими вірусами іншого господ (шифт). Коли антигенна специфічність гемаглютеніну змінюється стільки, що набутий протягом минулої епідемії імунітет стає нсефв
вним, починається нова епідемія грипу. Протягом останніх 50 років епідемії грипу серед людей різних країн світу були пов’язані з процесами антигенного шифту. У 1933 р. епідемію грипу викликав штам ARP8, у 1947 р. його замінив штам А (ГМі), у 1957 р. - штам А (Синга- нур / Н2 N2), у 1969 р. - штам А (Гонконг / Н3 N2), а у 1997 р. - штам А (Сідней /5/97/Нз N2), причому кожна епідемія була зв’язана в основному з принциповими змінами гемаглютеніну.
При вірусних інфекціях відбувається інтенсивний розпад власних інфікованих клітин під впливом імунних факторів (антитіл, цитотоксичних Т-лімфоцитів, макрофагів). Це може призводити до розвитку лнтоімунних захворювань, а утворення імунних комплексів з вірусно- і о матеріалу і антитіл - до появи імунокомплексних уражень.
Імунітет при мікозах
Набутий імунітет при мікозах характеризується різноплановістю проявів, пов’язаних складністю антигенного складу гриба, його мін- нітістю залежно від умов існування, форми і стадії мікозу. Підґрунтя імунітету при мікозах становить клітинний імунітет. При зараженні патогенними грибами розвивається ГСТ, яку можна виявити за допомогою шкірних проб або реакції in vitro (реакції гальмування міграції макрофагів, бластотрансформації тощо). Імунні реакції клітинного шиу можуть зберігатись протягом тривалого часу, нерідко протягом \1 І.ОГО життя.
1 Іри глибоких мікозах можуть з’являтись антитіла, які належать до ж їх класів імуноглобулінів, однак вони не відіграють суттєвої ролі в механізмах захисту від грибів, а є лише свідками захворювання. Вна-
н і цок недосконалості імунітету досить часто мікози набувають хронічного, рецидивного перебігу. Антитіла, що утворюються, викликами 1.1іерехресні реакції між різними видами патогенних грибів.
Інтенсивність імунітету при мікозах залежить від форми захворю- нінпгт При поверхневих мікозах (трихофітії, мікроспорії) розвиваєть- м і паГжа імунна відповідь, більш сильний імунітет, який розвиваєть- t и ї ї клітинним або гуморальним типом, виникає у людей при під- міі’ 11 ч її їх (хромомікоз, споротрихоз) і глибоких мікозах (гістоплазмоз, (*111и тмікоз). Грибкові захворювання легко виникають на фоні вро- •И'ітшх і набутих імунодефіцитів, особливо на фоні дефіциту системи І чімі|іоцитІВ.