

Занятие 3 и 4
.pdfЗанятие 3.
Тема: Закономерности изменчивости. Хромосомная теория наследственности. Комбинативная и мутационная изменчивость. Генные, хромосомные и геномные мутации
Цель занятия: Сформировать представления о хромосомном и генном механизмах определения пола и закономерностях наследования признаков, сцепленных с полом. Изучить закономерности сцепленного наследования и особенности наследования при кроссинговере. Сформировать представление о формах и видах наследственной изменчивости, их причинах и роли в эволюции и индивидуальной жизни человека.
Задачи занятия:
1.Ознакомиться с теоретическим материалом.
2.Научиться решать задачи на сцепленное с полом наследование.
3.Научиться решать задачи на сцепленное наследование и кроссинговер.
Формируемые компетенции: ОК-1, ПК-9, ПК-31, ПК-32
Студент должен знать:
1.Понятие о поле и половых признаках.
2.Понятие о половом диморфизме.
3.Различия между аутосомами и гетеросомами.
4.Понятие о гомогаметном и гетерогаментом поле.
5.Хромосомный механизм определения пола при образовании зиготы.
6.Причины наблюдаемого в природе соотношения полов в разных возрастных группах.
7.Морфологию половых хромосом. Особенности строения и расположения генов Х-
иY-хромосом человека.
8.Классификацию разных видов сцепленного с полом наследования.
9.Особенности наследования и формирования признаков, контролируемых генами Х-
иY- хромосом человека.
10.Характер проявления в фенотипе доминантных и рецессивных генов, находящихся в Х- и Y- хромосоме человека.
11.Примеры признаков:
Сцепленных с Х-хромосомой
Сцепленных с Y-хромосомой
Частично сцепленных с полом
Аутосомных, зависимых от пола.
12.Х- и Y- половой хроматин. Причины инактивации одной из Х-хромосом у гомогаметного пола.
13.Особенности и цитологические основы наследования при сцеплении генов и кроссинговере.
14.Виды и причины наследственной изменчивости: комбинативной и мутационной.
15.Виды и причины мутаций: генные, хромосомные, геномные.
16.Основные свойства мутаций.
Студент должен уметь:
1.Давать цитологическое обоснование качественных и количественных закономерностей наследования признаков при разных видах сцепленного с полом наследования.
2.Давать цитологическое обоснование качественных и количественных закономерностей наследования признаков при разных видах взаимодействия генов.

3.Решать задачи на запись гамет организма с известным генотипом, включающим аутосомные и сцепленные с половыми хромосомами генами.
4.Находить генотип организма, анализируемого по аутосомным и сцепленным с полом признакам, по образующимся у него гаметам.
5.Решать задачи на наследование признаков, сцепленных с полом:
-с Х-хромосомой
-с Y-хромосомой
-частично сцепленных с полом.
6.Решать задачи на наследование нескольких признаков, одни из которых сцеплены с полом, а другие обусловлены аутосомными генами.
7.Решать задачи на сцепленное наследование при полном сцеплении генов и с учетом кроссинговера.
Студент должен владеть:
1.Медико-функциональным понятийным аппаратом и специальными терминами.
2.Навыками записи и решения генетических задач.
Оснащение занятия:
1.Таблицы:
-сцепленное наследование
-схема кроссинговера
-виды изменчивости
-мутационная изменчивость
-мутации у дрозофилы Микроскоп световой Микропрепараты:
Дрозофила – норма Дрозофила – мутация Изменение антенн в ножные структуры
Дрозофила – мутация Бескрылая форма
Хронологическая карта занятия:
1.Организационная часть
2.Тестовый контроль базового уровня знаний
3.Объяснение практического задания
4.Самостоятельная работа
5.Проверка выполненных работ в альбомах
6.Контроль конечного уровня знаний
7.Установка задания для подготовки к следующей теме
Теоретический обзор Изучите материал, используя следующую литературу:
1.«Биология», учебник в 2 т/ под ред. В.Н. Ярыгина. - М.: ГЭОТАР-Медиа, 2011.
2.«Биология», учебник в 2 т/ под ред. В.Н. Ярыгина, М.: Медицина, 2006 г
3.Биология/ А.А. Слюсарев, С.В. Жукова, - К: Высшая школа, 1987.
4.Электронная версия учебно-методического пособия (каф. мед. биологии и генетики)
Дополнительная литература:
5. Гены./ Льюин Б. – М.: Мир. 1987.
Базы данных по геному и генетическим заболеваниям человека
1.Genes and Diseases (гены и болезни — издание Национального Центра Биотехнологической Информации, США) — http://www.ncbi.nlm.nih.gov/books/NBK22183/
— элементарный справочник по генетическим болезням. Рекомендуется для начинающих.
2.OMIM (Online Mendelian Inheritance in Man, издание Национального Центра Биотехнологической Информации, США) — http://www.ncbi.nlm.nih.gov/omim — de facto
применяется многими научными журналами для идентификации заболевания, имеющего генетическую причину развития (как монотак и полигенного характера) или предрасположенность к развитию заболевания. Прекрасный и существенно полный источник информации по генам человека и по генным заболеваниям.
3.List of Genetic Disorders (список генетических заболеваний) —
http://en.wikipedia.org/wiki/List_of_genetic_disorders — не только список, но и отсылки на статьи по конкретным заболеваниям. Для начинающих.
4. Список наследственных заболеваний — http://ru.wikipedia.org/wiki/%D0%A1%D0%BF%D0%B8%D1%81%D0%BE%D0%BA_%D0 %BD%D0%B0%D1%81%D0%BB%D0%B5%D0%B4%D1%81%D1%82%D0%B2%D0%B5 %D0%BD%D0%BD%D1%8B%D1%85_%D0%B7%D0%B0%D0%B1%D0%BE%D0%BB% D0%B5%D0%B2%D0%B0%D0%BD%D0%B8%D0%B9 — облегчённый вариант англоязычной версии (3), частично имеются отсылки на статьи по конкретным заболеваниям. Для начинающих и не владеющих английским.
СОДЕРЖАНИЕ ЗАНЯТИЯ
1.Хромосомная теория наследственности. Сцепленное наследование.
1.1.Явление сцепления генов.
Закон Менделя о независимом наследовании признаков справедлив лишь в том случае, если аллели разных генов находятся в разных, негомологичных друг другу хромосомах. Гены, расположенные в одной хромосоме, будут наследоваться вместе.
Основы хромосомной теории заложили работы немецкого биолога Т. Бовери (1902–1907) и американского цитолога У. Сеттона (1902–1903), которые независимо друг от друга предположили, что гены расположены в хромосомах, и связали закономерности Менделя, описывавшие поведение наследственных факторов, с поведением хромосом во время мейоза и при оплодотворении. Детальная разработка хромосомной теории была произведена Т.Х. Морганом и его учениками (начиная с 1910 г.).
Совместное наследование разных генов, расположенных в одной и той же паре гомологичных хромосом, называется сцеплением генов (Закон Т. Моргана).
Совокупность генов, расположенных в одной паре гомологичных хромосом, образуют группу сцепления. Количество групп сцепления равно гаплоидному набору хромосом. Если гены, находящиеся в одной хромосоме, передаются вместе – сцепление называется полным. Иногда полное сцепление нарушается по причине кроссинговера. Кроссинговер – это обмен одинаковыми участками гомологичных хромосом, когда гомологичные хромосомы коньюгируют (сближаются) друг с другом. Кроссинговер происходит при мейозе в профазу первого деления. Кроссоверных гамет образуется намного меньше, чем некроссоверных. Причем частота кроссинговера и, следовательно, частота образования кроссоверных гамет зависят от расстояния, на котором аллели данных генов находятся друг от друга в хромосоме. Чем больше расстояние между генами, тем выше частота кроссинговера и частота образования кроссоверных гамет.
Рассмотрим один из первых экспериментов Т. Моргана по изучению сцепленного наследования. При скрещивании дрозофил, различающихся по двум парам сцепленных
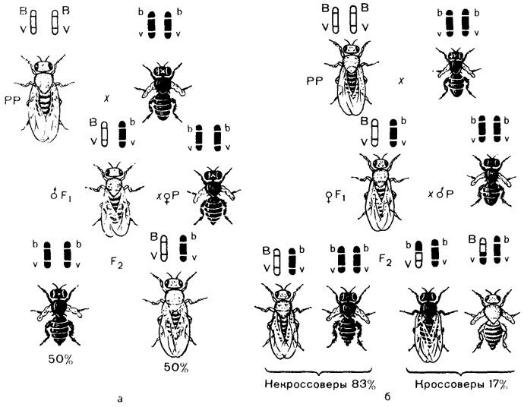
признаков — серых с нормальными крыльями (BV||BV) и черных с зачаточными крыльями (bv||bv)— гибриды F1 были серыми с нормальными крыльями (BV||bv) (рис. 1).
Далее были проведены два типа анализирующих скрещиваний. В первом из них брали дигетерозиготных (BV||bv) самцов F1, и скрещивали с гомозиготными по рецессивным аллелям самками (bv||bv), а во втором — дигетерозиготную самку (BV||bv) скрещивали с самцами, рецессивными по обоим признакам (bv||bv) (черное тело и зачаточные крылья). Результаты этих скрещиваний оказались разными (см. рис. 1).
Рис. 1. Сцепленное наследование генов окраски тела и состояния крыльев у плодовой мухи.
Ген серого цвета (В) доминирует над геном черного цвета тела (b), ген длинных крыльев (V) - над геном коротких крыльев (v). В и V находятся в одной хромосоме.
а - полное сцепление генов вследствие отсутствия перекреста хромосом у самцов дрозофилы б - неполное (частичное) сцепление признаков вследствие перекреста хромосом у самок дрозофилы
В первом случае были получены потомки с исходными для этого опыта родительскими (Р) фенотипами, т. е. серые мухи с нормальными крыльями (BV||bv) и черные мухи с зачаточными крыльями (bv||bv) в отношении 1:1. Следовательно, данная дигетерозигота образует только два типа гамет (BV и bv) вместо четырех. Исходя из указанного расщепления, можно предположить, что у самца наблюдается полное сцепление генов.
Во втором случае в F2 наблюдалось иное расщепление. Кроме родительских комбинаций признаков, появились новые — мухи с черным телом и нормальными крыльями (bV||bv) , а также с серым телом и зачаточными крыльями (Bv||bv). Правда, количество рекомбинантных потомков невелико и составляет 17%, а родительских — 83%. Причиной появления небольшого количества мух с новыми сочетаниями признаков является кроссинговер, который приводит к новому рекомбинантному сочетанию аллелей генов b и v в гомологичных хромосомах. Эти обмены происходят с вероятностью 17% и в итоге дают два класса рекомбинантов с равной вероятностью — по 8,5%.
Зная частоту образования кроссоверных гамет или частоту кроссинговера, можно определить расстояние между генами. Расстояние между генами в хромосоме оценивают в условных единицах, называемых морганидами. Одна морганида – это единица расстояния между генами, при котором у организма образуется 1 % кроссоверных гамет. То есть, если гены в одной хромосоме находятся на расстоянии 5 морганид, то это означает, что у организма образуется 5% кроссоверных гамет.
1.2. Генетические карты хромосом.
Кроссинговер позволяет построить генетическую карту хромосомы. Генетическая карта содержит информацию о порядке расположения генов в этой хромосоме и расстоянии между ними. Возможность подобного картирования основана на постоянстве процента кроссинговера между определенными генами.
Информация, которая содержится в генетических картах хромосом имеет большое значение в исследовательской деятельности – например, с помощью генно-инженерных методов, возможно создание штаммов микроорганизмов, способных синтезировать необходимые для фармакологии и сельского хозяйства белки, гормоны и другие сложные органические вещества.
Знания о локализации гена в определенной хромосоме используются в медицине при диагностике ряда тяжелых наследственных заболеваний человека. Уже теперь появилась возможность использовать эту информацию для генной терапии, т. е. для исправления структуры или функции генов.
Сравнение генетических карт разных видов живых организмов способствует также пониманию эволюционного процесса.
1.3. Основные положения хромосомной теории наследственности.
Анализ явлений сцепленного наследования, кроссинговера, сравнение генетической и цитологической карт позволяют сформулировать основные положения
хромосомной теории наследственности:
1.Гены локализованы в хромосомах. При этом различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов каждой из негомологичных хромосом уникален.
2.Аллельные гены занимают одинаковые локусы в гомологичных хромосомах.
3.Гены расположены в хромосоме в линейной последовательности.
4.Гены одной хромосомы образуют группу сцепления. Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и n+1 у гетерогаметных особей. При этом сила сцепления находится в обратной зависимости от расстояния между генами.
5.Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссинговера возникают гаметы, хромосомы которых содержат новые комбинации генов.
6.Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера. За единицу расстояния между генами принимают 1 морганиду (1% кроссинговера) или процент появления кроссоверных особей.
7.Для выяснения характера расположения генов в хромосомах и определения частоты кроссинговера между ними строят генетические карты. Карта отражает порядок расположения генов в хромосоме и расстояние между генами одной хромосомы.
8.Каждый биологический вид характеризуется определенным набором хромосом — кариотипом.
2. Генетика пола
Генетика пола - раздел генетики, изучающий роль механизмов наследственности и наследственной изменчивости в процессе определения и дифференциации пола.
Пол – это совокупность морфологических и физиологических особенностей организма, обеспечивающих продолжение рода. Формирование пола начинается с момента зачатия. При зачатии закладывается генетический (хромосомный ХХ или ХУ) пол. В процессе эмбриогенеза появляется гонадный (семенники или яичники) и соответственно гаметный (сперматозоиды или яйцеклетка) пол.
Различают две группы связанных с полом отличительных признаков: первичные и вторичные половые признаки. Первичные половые признаки – это наличие определенных половых желез и половых протоков. Первичные половые признаки непосредственно связаны с полом, они непосредственно принимают участие в размножении и определяют формирование вторичных половых признаков. Вторичные половые признаки – это те признаки, по которым мужские и женские особи отличаются друг от друга, но которые непосредственного участия в размножении не принимают.
После рождения, в процессе созревания повышается уровень дифференциации полов и происходит увеличение проявления полового диморфизма.
Половой димор изм – анатомические различия между самцами и самками одного и того же биологического вида, не считая половых органов. Половой диморфизм может проявляться в различных физических признаках, например: размер (у млекопитающих и многих видов птиц самцы более крупные и тяжёлые, чем самки. У земноводных и членистоногих самки, как правило, крупнее самцов); волосяной покров (борода у мужчин, грива у львов или бабуинов); окраска (цвет оперения у птиц, особенно у утиных); кожа (характерные наросты или дополнительные образования, такие как рога у оленевых, гребешок у петухов); зубы (бивни у самцов индийского слона, более крупные клыки у самцов моржей и кабанов). У некоторых животных, половой диморфизм проявляется только во время спаривания. Считается, чем различнее являются вклады обоих полов в уход за потомством, тем больше выражен половой диморфизм.
2.1. Половые хромосомы. Соотношение полов при оплодотворении.
Пол, как и любой другой признак организма, наследственно детерминирован. Важнейшая роль в генетической детерминации пола и в поддержании закономерного соотношения полов принадлежит хромосомному аппарату.
При изучении кариотипов (хромосомных наборов) самцов и самок ряда животных между ними были обнаружены некоторые различия. В кариотипах всех клеток мужских и женских особей имеются пары гомологичных друг другу хромосом – аутосомы. Каждая такая хромосома имеет парную себе хромосому, и они по всем признакам полностью гомологичны друг другу: содержат одинаковые наборы генов, одинаковы по размеру, форме, положению первичной перетяжки и соотношению плеч. Отличия наблюдаются только в одной паре. Такие хромосомы, по которым самцы и самки отличаются друг от друга, называют половыми хромосомами, гоносомами или гетеросомами. Те из них,
которые являются парными у одного из полов, называют X-хромосомами (например, у дрозофилы и млекопитающих) или Z-хромосомами (например, у птиц). Непарная половая хромосома, имеющаяся у особей только одного пола, была названа У-хромосомой (у дрозофилы и млекопитающих) или W-хромосомой (у птиц). Впервые половые хромосомы были обнаружены в 1891 немецким исследователем Г. Хеннингом, а в начале 20 в. К. Мак-Кланг и Э. Уилсон обнаружили различия в хромосомных наборах у самцов и самок насекомых.
Пол, у представителей которого имеются различные половые хромосомы (X и У), образует гаметы двух типов (половина с X-хромосомой и половина с У-хромосомой), то
есть, является гетерогаметным, а пол, у представителей которого в каждой клетке содержатся одинаковые половые хромосомы (X-хромосомы), — гомогаметным. Организмы, у которых в кариотипе имеется только одна половая хромосома – моногаметными (у самцов прямокрылых, например, кузнечиков). У человека, млекопитающих, большинства насекомых – гомогаментым является женский пол, а гетерогаментым – мужской; у птиц, рептилий и бабочек, наоборот, гомогаметный пол – мужской, а гетерогаментый – женский.
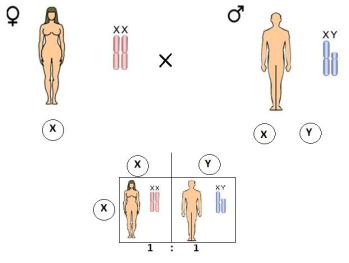
Согласно хромосомной теории К.Корренса (1907), пол будущего потомка определяется сочетанием половых хромосом в момент определения. Так, например, у человека в кариотипе имеется 46 хромосом (44 – аутосомы и 2 половые хромосомы). При созревании половых клеток в результате мейоза гаметы получают гаплоидный набор хромосом. В каждой яйцеклетке есть 22 аутосомы + Х-хромосома. Сперматозоиды дают гаметы двух видов: половина содержит 22 аутосомы + Х – половую хромосому, и половина содержит 22 аутосомы + Y – половую хромосому. Пол будущего ребенка определяется в момент оплодотворения и зависит от того, каким сперматозоидом будет оплодотворена данная яйцеклетка. Если яйцеклетка оплодотворена сперматозоидом, имеющим Х – хромосому, развивается женский организм, если Y – хромосому – мужской.
Теоретически вероятность рождения мальчика и девочки будет одинакова, т.е. пол будет наследоваться как менделирующий признак, в соотношении 1:1 (рис.1).
Рис.1. Соотношение полов при оплодотворении.
Различают первичное соотношение полов, точнее, соотношение эмбрионов и вторичное - соотношение мальчиков и девочек среди новорожденных.
Теоретически соотношение полов в момент оплодотворения должно быть близким 1:1, так как встреча яйцеклетки со сперматозоидом, содержащим Х- или Y- половую хромосому, равновероятна.
При обследовании у человека обнаружено, что на 100 женских зигот образуется 140-
160мужских (первичное соотношение полов).
Кмоменту рождения на 100 девочек приходится 103-105 мальчиков 9 (вторичное соотношение полов).
Третичное соотношение полов (постнатальный период) к 20-ти годам на 100 девушек приходится 100 юношей; к 50-ти годам на 100 женщин - 85 мужчин, а к 85-ти годам на 100 женщин - 50 мужчин.
Отсюда напрашивается вывод о большей жизнестойкости женского организма, что может быть объяснено, наряду с другими причинами, мозаицизмом женского организма
по половым хромосомам, а также то, что мужской организм имеет всего одну Х – хромосому, и все гены (доминантные и рецессивные) проявляют свое действие, это делает мужской организм менее жизнеспособным.
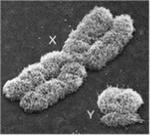
2.2.Мор ология половых хромосом.
Учеловека Х-хромосома крупная, имеет два почти одинаковых по размеру плеча и центромеру, лежащую почти в центре хромосомы – называется метацентрической. Y- хромосома человека маленькая, неравноплечая, центромера сильно смещена к концу плеча (рис. ) – акроцентрическая. Y-хромосома содержит небольшое число генов, в том числе и ген CRY, наличие которого обуславливает развитие зародыша по мужскому типу.
рис. -- . Х- и Y- хромосомы человека.
В Х- и Y-хромосоме человека различают три группы участков:
1)негомологичные участки Х-хромосомы, которые отсутствуют в Y-хромосоме.
Вэтих участках располагаются гены, которые наследуются только с Х-хромосомой и поэтому называются сцепленными с Х-хромосомой. Частота встречаемости таких генов среди мужчин и женщин не одинакова. Так, рецессивные сцепленные с Х-хромосомой гены проявляются чаще у мужчин, чем у женщин, например, гемофилия, дальтонизм.
Доминантные сцепленные с Х-хромосомой гены проявляются чаще у женщин, например, гипоплазия эмали (эмаль коричневого цвета). Примеры записи: ХАХА, ХАY, XaY
2)негомологичные участки Y-хромосомы, которые отсутствуют в Х-хромосоме. В
этих участках расположены гены, которые передаются только по мужской линии,
называются сцепленными с Y-хромосомой, например, гипертрихоз (повышенное оволосение ушной раковины). Примеры записи: XYa, XYA.
3)гомологичные участки Х- и Y-хромосомы: в этих участках расположены гены,
которые с одинаковой вероятность проявляются у мужчин и женщин, называются
частично сцепленные с полом, например, пигментная ксеродерма, пигментный ретинит. Примеры записи XAYa.
По генам, расположенным в негомологичных участках Х и Y-хромосом, мужской организм является гемизиготным, так как в мужском организме каждый такой ген представлен не двумя аллелями, а одним аллелем гена.
X-хромосома человека содержит около 150 миллионов пар оснований, что составляет примерно 5 % ДНК в клетках женщин, 2,5 % в клетках мужчин. Несет более 1400 генов (в то время как Y-хромосома несет всего 86 белок-кодирующих генов, несущих информацию о 23 индивидуальных белках.) (рис. 2). У женщин две X- хромосомы; у мужчин одна X-хромосома и одна Y-хромосома. Одна X-хромосома наследуется от матери, а вторая (только у женщин) от бабушки по отцовской линии.
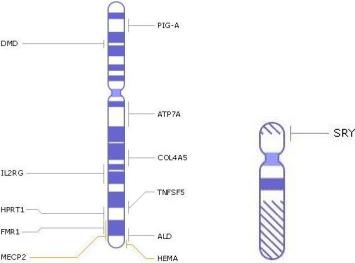
Рис. 2 . Некоторые гены, сцепленные с Х- и Y-хромосомами.
Так, например, в Х-хромосоме находятся гены, отвечающие за следующие генные болезни (рис. 2):
-альменолейкодистрофия (ALD);
-гемофилия А (HEMA);
-гипер IgM синдром (TNFSF5);
-мышечная дистро ия Дюшенна (DMD);
-пароксизмальная ночная гемоглобинурия (PIGA);
-синдром Альпорта (COL4A5);
-синдром Леша-Наяна (HPRT1);
-синдром Мартина-Белла (FMR1);
-синдром Менкеса (ATP7A);
-синдром Рета (MECP2);
-Х-сцепленный тяжелый комбинированный иммунодефицит (IL2RG)
Y-хромосома содержит ген SRY, который определяет мужской пол, а также гены, необходимые для нормального формирования сперматозоидов.
Для нормальной жизнедеятельности организму достаточно функционирования одной Х-хромосомы. Одновременное функционирование генов обеих Х-хромосом в клетках женского организма привело бы к нежелательному увеличению количества продуктов их активности – белков – по сравнению с клетками мужского организма. Поэтому, на ранних этапах эмбрионального развития у гомогаметного пола (женского пола) одна из Х хромосом в соматических клетках инактивируется. Процесс называется лайонизация. Инактивированная Х-хромосома называется Х-половым хроматином или тельцем Барра. Тельце располагается рядом с ядерной оболочкой и виден в световой микроскоп в виде крупной глыбки гетерохроматина. Количество телец Барра в клетке равняется общему количеству Х хромосом в кариотипе минус один. Инактивация Х- хромосом является сложным и многоэтапным процессом, детальные механизмы которого до конца выяснены. Основные особенности, связанные с инактивацией Х-хромосомы в женском организме:
a.инактивация сопровождается метилированием цитозина,
b.инактивация сопровождается глобальным гипоацетилированием гистонов, повышающим прочность взаимодействия гистоновых белков с молекулой ДНК,
c.инактивация сопровождается выраженной конденсацией хроматина,
d.инактивация начинается в определенном участке Х-хромосомы, называемом центром инактивации, и затем распространяется на всю Х-хромосому, основным геном центра инактивации является XIST,
e.инактивированное состояние Х-хромосомы передается от соматической клетки ее потомкам, образующимся в результате митоза,
f.инактивация не затрагивает клетки зародышевого пути, из которых образуются половые клетки, или происходит восстановление активности Х-хромосомы.
g.инактивация начинается на ранних этапах эмбрионального развития и носит случайный характер и инактивирована может быть любая из Х-хромосом.
Таким образом, у женского организма, гетерозиготного по какому-либо признаку, определяемому геном X-хромосомы, в разных клетках работают разные аллели этого гена (генетический мозаицизм). Случайная инактивация Х-хромосомы приводит к мозаичному проявлению у женщины тех генов Х-хромосомы, которые находятся у нее в гетерозиготном состоянии, в том числе по генам, вызывающим наследственные заболевания. Так, например, при гемофилии у гетерозиготной женщины часть клеток печени выделяют антигемофилический фактор, а часть клеток нет. Подобный мозаицизм наблюдается при цветовой слепоте красно-зеленого типа, недостаточности глюкозо-6- фосфатдигидрогеназы. Однако следует отметить, что при нормальной инактивации Х- хромосомы, мозаицизм обычно не приводит к патологическим изменениям.
2.3. Генное определение пола.
Установлено, что, кроме гена SRY, несколько аутосомных генов играют определенную роль в мужской дифференциации пола. Один из них — ген супрессор опухоли Вильмса, кодирует фактор транскрипции, связывающийся со специфическими ДНК-последовательностями. Точковые мутации в этом гене иногда приводят к формированию гонадно противоположного пола. Второй ген такого же типа — SOX9. Белок, кодируемый этим геном, также содержит HMG-бокс. Мутации в гене SOX9 вызывают кампомелическую диспла-зию, при которой у индивидуумов 46, XYфенотипически развивается женский пол.
Для нормального развития гонад необходимы также X-хромосомные локусы. У некоторых лиц с гонадным дисгенезом 46,XY наблюдалась дупликация области Хр21. В этой области находится ген DAX1, белок которого функционирует как транскрипционный фактор. Еще один ген в хромосоме X — ХН2, играет не вполне выясненную роль в дифференцировке яичек. Он кодирует фермент геликазу, которая расплетает ДНК, делая ее доступной для транскрипционных факторов.
Нет сомнений в том, что современные представления о генетическом контроле становления пола далеко не полные. Это относится не только к тому, какие гены вовлечены в данный процесс, но и к тому, на каких стадиях развития и в каких тканях эти гены проявляются. На примере генетического контроля определения пола мы сталкиваемся с некоторыми кардинальными проблемами генетики развития — ткане- и времяспецифичности действия генов. При образовании полового гребешка из мезонефроса и целомического эпителия в соответствующих тканях обнаружена активность гена WT1, а в зачатке надпочечника, который также имеет отношение к образованию полового гребешка, — генов SF1 и DAX1. Половой гребешок формирует недифференцированные гонады при участии тех же генов.
Хорошо изученный пример генного определения пола — определение пола у дрожжей. У большинства штаммов дрожжей вегетативные почкующиеся клетки гаплоидны, а зигота, которая образуется при половом процессе, делится мейозом. У дрожжей есть гены а и альфа, которые находятся в разных локусах одной хромосомы. Оба они неактивны. Один из этих генов удваивается и посылает свою копию в третий локус — МАТ-локус. Там этот ген включается и определяет пол (спариваться могут только клетки