

2 курс / Нормальная физиология / ВСЯ ФИЗИОЛОГИЯ МЕДВУЗА
.pdfОБЩИЕ ПРИНЦИПЫ РЕГУЛЯЦИИ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
1. Понятие о гомеостазе и гомеокинезе. Саморегуляторные принципы поддержания постоянства внутренней среды организма
Гомеостаз — саморегуляция, способность открытой системы сохранять постоянство своего внутреннего состояния посредством скоординированных реакций, направленных на поддержание динамического равновесия, если говорить проще —> стремление системы воспроизводить себя, восстанавливать утраченное равновесие, преодолевать сопротивление внешней среды.
Гомеокинез – это процесс изменения работы организма, направленный на установление гомеостаза.
Для эффективного функционирования биологических процессов необходимы определенные условия, многие из которых должны быть постоянными. И чем эти условия стабильнее, тем надежнее функционирует биологическая система. К этим условиям относятся те, которые способствуют сохранению стабильного уровня обмена веществ. Для этого необходимо поступление исходных ингредиентов обмена и удаление конечных метаболитов, поступление кислорода. Эффективность протекания обменных процессов обеспечивается определенной интенсивностью внутриклеточных процессов, обусловленной в первую очередь активностью ферментов. В то же время ферментативная активность зависит не только от поступления ингредиентов и удаления метаболитов, но и от температуры. Стабильность большинства условий необходима на любом структурно-функциональном уровне — начиная от отдельной биохимической реакции, клетки, вплоть до сложных функциональных систем организма. В реальной жизни эти условия часто могут нарушаться.
Возникшие изменения отражаются на состоянии биологических объектов, на течении в них процессов обмена. Чем сложнее устроена биологическая система, тем большие отклонения от стандартных условий она выдерживает без существенных нарушений жизнедеятельности. Это связано с наличием в организме соответствующих механизмов, направленных на восстановление возникших изменений. Так, например, в клетке активность ферментативных процессов при понижении температуры на каждые 10 °С снижается в 2-3 раза.
В результате сохраняется стабильность этого условия для протекания ферментативных реакций на неизменном уровне. В процессе эволюции происходило формирование приспособительных реакций, направленных на поддержание постоянных условий внешней среды организма. Они имеются как на уровне отдельных биологических процессов, так и всего организма. Каждое из этих условий характеризуют соответствующие параметры. Поэтому системы

регуляции постоянства условий контролируют постоянство этих параметров. А если указанные параметры почему-либо отклоняются от нормы, то механизмы регуляции обеспечивают возврат их к исходному уровню.
2. Основные формы регуляции физиологических функций. Взаимоотношение нервных и гуморальных механизмов регуляции.
Физиологическая регуляция – это активное управление функциями организма и его поведением для поддержания оптимального уровня жизнедеятельности, постоянства внутренней среды и обменных процессов с целью приспособления организма к меняющимся условиям среды.
Механизмы физиологической регуляции:  нервный
нервный  гуморальный.
гуморальный.
Гуморальная физиологическая регуляция для передачи информации использует жидкие среды организма (кровь, лимфу). Сигналы передаются посредством химических веществ: гормонов, медиаторов, биологически активных веществ, электролитов и т.д.
Особенности гуморальной регуляции:
 не имеет точного адресата – с током биологических жидкостей вещества могут доставляться к любым клеткам организма;
не имеет точного адресата – с током биологических жидкостей вещества могут доставляться к любым клеткам организма;
 скорость доставки информации небольшая – определяется скоростью тока биологических жидкостей – 0,5-5 м/с;
скорость доставки информации небольшая – определяется скоростью тока биологических жидкостей – 0,5-5 м/с;
 продолжительность действия.
продолжительность действия.
Нервная физиологическая регуляция для переработки и передачи информации опосредуется через центральную и периферическую нервную систему. Сигналы передаются с помощью нервных импульсов.
Особенности нервной регуляции:
 имеет точный адресат – сигналы доставляются к строго определенным органам и тканям;
имеет точный адресат – сигналы доставляются к строго определенным органам и тканям;
 большая скорость доставки информации – скорость передачи нервного импульса – до 120 м/с;
большая скорость доставки информации – скорость передачи нервного импульса – до 120 м/с;
 кратковременность действия.
кратковременность действия.
Для нормальной регуляции функций организма необходимо взаимодействие нервной и гуморальной систем.
Нейрогуморальная регуляция объединяет все функции организма для достижения цели, при этом организм функционирует как единое целое.
Организм находится в неразрывном единстве с внешней средой благодаря активности нервной системы, деятельность которой осуществляется на основе рефлексов.

ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
1.Общие и частные свойства возбудимых тканей. Раздражители,
их классификация. Мера возбудимости.
Основным свойством живых клеток является раздражимость, т. е. их способность реагировать изменением обмена веществ в ответ на действие раздражителей.
Возбудимость — свойство клеток отвечать на раздражение возбуждением. К возбудимым относят нервные, мышечные и некоторые секреторные клетки.
Возбуждение — ответ ткани на ее раздражение, проявляющийся в специфической для нее функции (проведение возбуждения нервной тканью, сокращение мышцы, секреция железы) и неспецифических реакциях (генерация потенциала действия, метаболические изменения).
Cвойства возбудимых тканей
4 свойства: возбудимость, проводимость, рефрактерность, лабильность. Для мышечной ткани характерна также сократимость.
Возбудимость – способность ткани отвечать на раздражение изменением ряда своих свойств. Показатель возбудимости – порог раздражения. Это минимальное по силе раздражение, способное вызвать видимую ответную реакцию ткани.
Проводимость – способность ткани проводить возбуждение по всей своей длине. Показатель проводимости – скорость проведения возбуждения.
Рефрактерность – способность ткани терять или снижать возбудимость в процессе возбуждения. При этом в ходе ответной реакции ткань перестает воспринимать раздражитель.
Лабильность – способность ткани генерировать определенное число волн возбуждения в единицу времени в точном соответствии с ритмом наносимого раздражения. Лабильность определяется продолжительностью рефрактерного периода (чем короче рефрактерный период, тем больше лабильность).
Сократимость – способность мышцы отвечать сокращением на раздражение.
Раздражитель – фактор, способный вызвать ответную реакцию возбудимых тканей. В условиях физиологического эксперимента в качестве раздражителя чаще всего используют электрический ток.
Хронаксия – наименьший промежуток времени, в течение которого ток силой в 2 реобазы (пороговая сила раздражителя для электрического тока) вызывает в ткани возбуждение.
Различают два вида биологических реакций: специфические и неспецифические. Специфические реакции характерны для какой-то строго определенной ткани
Все многообразие раздражителей можно выделить в отдельные группы. Классификация раздражителей зависит от того, что берется за основу:
По своей природе раздражители бывают: 1) химические; 2) физические; 3) механические; 4) термические; 5) биологические.

Потенциал покоя — это разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны, когда клетка находится в состоянии физиологического покоя.
Потенциал действия - скачкообразное изменение постоянного мембранного потенциала с отрицательной поляризации на положительную и обратно

Биологические мембраны, их строение и функциональные особенности. Ионные каналы, их классификация и роль. Виды транспорта веществ через биологические мембраны.
Строение и функции ионных каналов.
Ионы Na+, K+, Са2+, Сlпроникают внутрь клетки и выходят наружу через специальные, заполненные жидкостью каналы. Размер каналов довольно мал (диаметр 0,5—0,7 нм). Расчеты показывают, что суммарная площадь каналов занимает незначительную часть поверхности клеточной мембраны.
Именно ионные каналы обеспечивают два важных свойства мембраны: селективность и проводимость.
Селективность, или избирательность, канала обеспечивается его особой белковой структурой. Большинство каналов являются электроуправляемыми, т. е. их способность проводить ионы зависит от величины мембранного потенциала. Канал неоднороден по своим функциональным характеристикам, особенно это касается белковых структур, находящихся у входа в канал и у его выхода (так называемые воротные механизмы).
Пассивный транспорт  Простая диффузия
Простая диффузия  Осмос
Осмос  Диффузия ионов
Диффузия ионов
 Облегченная диффузия Активный транспорт
Облегченная диффузия Активный транспорт  Первично-активный транспорт
Первично-активный транспорт
 Вторично-активный транспорт
Вторично-активный транспорт

Структура биологических мембран. Фосфолипиды образуют двойной прерывистый слой. В этот слой включены белки, полярные группы которых сохраняют контакт с водной фазой. Некоторые белки пронизывают мембрану насквозь, другие погружены в липидный бислой наполовину. Часть белков связана друг с другом; другие в большей или меньшей степени окружены липидами. Одни из них являются ионными каналами, другие содержат боковые цепи гликозаминогликанов.
Гликокаликс. Поверхность мембраны покрыта гликокаликсом — трехмерной сетью нитей гликозаминогликанов, соединенных между собой при помощи кальциевых мостиков. Гликокаликс обеспечивает механическую прочность мембраны, участвует в межклеточных взаимодействиях, рецепции, иммунологическом дифференцировании, разделяет молекулы веществ, контактирующих с клеткой, по величине и заряду.
Липиды. Молекулы липидов, образующих бислой, амфотерны. Своими гидрофильными головками они обращены в сторону водных фаз (межклеточная жидкость и цитоплазма) и формируют внешнюю и внутреннюю поверхности мембраны. Важнейшей особенностью мембранных липидов является способность к перекисному окислению (ПОЛ) с образованием свободных радикалов.
Белки. Функциональное отличие мембраны одной клетки от мембраны другой определяется наличием в ней специфических мембранных белков.
Белки, погруженные в фосфолипидный слой и пронизывающие его насквозь, называются внутренними мембранными белками, или белковыми каналами.
Другие белки — периферические — прикреплены к поверхности клетки.
С учетом выполняемых функций мембранные белки всех клеток делят на 5 классов: белки-насосы, белки-каналы, белкирецепторы, ферменты и структурные белки.
Функции мембран. Важнейшими функциями клеточных мембран являются барьерная, биотрансформирующая, транспортная, рецепторная, генерация электрических потенциалов и образование межклеточных контактов.
Белки-каналы представляют собой пути избирательного переноса ионов и заряженных молекул. Механизм переноса связан с конформацией белкаканала, в результате которой он открывается или закрывается. Взаимодействие рецептора с соответствующим ему лигандом инициирует закрытие или открытие связанного с рецептором канала.

Ионоселективные каналы делят на химические и электрозависимые. В первом случае раздражителем является вещество (медиатор, гормон, метаболит, лекарственное средство), во втором — возникающее в непосредственной близости от электрозависимого канала возбуждение, т.е. потенциал действия.
Ионоселективные каналы в зависимости от скорости их активации и переноса ионов делят на быстрые (например, натриевые) и медленные (например, калиевые, кальциевые).
Для каждого из переносимых через мембрану вида ионов существуют самостоятельные транспортные системы — ионные каналы (натриевые, калиевые, кальциевые, каналы для хлора), основные свойства и механизмы действия которых сходны. Ионный канал состоит из поры, воротного механизма, сенсора (индикатора), напряжения ионов в самой мембране и селективного фильтра.
Мембранно-ионные механизмы происхождения потенциала покоя. Статическая поляризация характеризуется наличием постоянной разности потенциалов между наружной и внутренней поверхностями клеточной мембраны и цитоплазмой, равной —60—90 мВ и называемой мембранным потенциалом (МП), или потенциалом покоя.
Функции клеточных мембран
1.Барьерная функция выражается в том, что мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. При этом мембрана принимает участие в механизмах электрогенеза. К ним относятся механизмы создания потенциала покоя, генерация потенциала действия, механизмы распространения биоэлектрических импульсов по однородной и неоднородной возбудимым структурам.
2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»).
3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах).
4.Высвобождение нейромедиаторов в синаптических окончаниях.
ФИЗИОЛОГИЯ МЫШЦ И НЕРВОВ
Структура миофибрилл и ее изменения при сокращении. Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни участки анизотропны, т. е. обладают двойным лучепреломлением. В обыкновенном свете они выглядят темными, а. в поляризованном— прозрачными в продольном направлении й непрозрачными в поперечном. Другие участки в Обыкновенной свете выглядят светлыми — они изотропны, т. е. не обладают двойным лучепреломлением. Анизотропные участки обозначают буквой А, изотропные — буквой I. В середине диска А различается светлая полоска Н, посередине диска / — темная полоска -представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы. Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрилл внутри одного волокна во время сокращения не смещаются по отношению друг к другу.
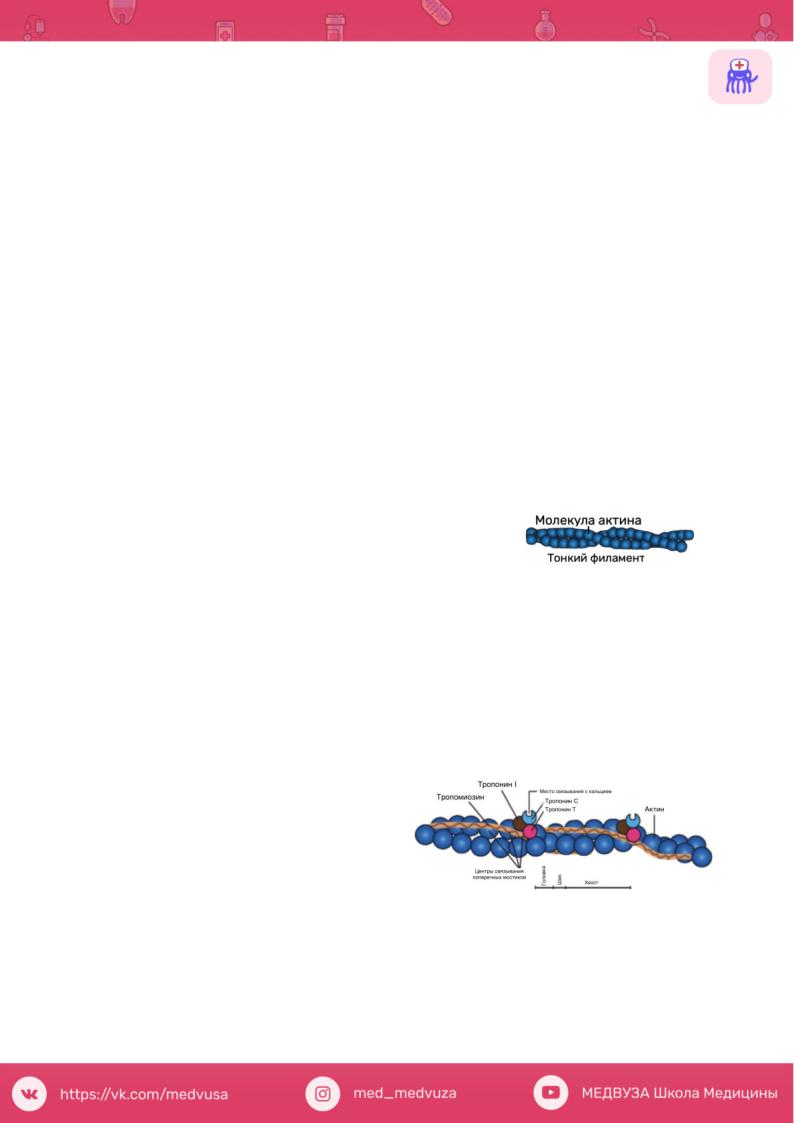
Молекула актина Это глобулярный белок, состоящий из одного полипептида, который
полимеризуется с другими молекулами актина и образует две цепи, обвивающие друг друга. Такая двойная спираль представляет собой остов тонкого филамента. На каждой молекуле актина есть участок связывания миозина. В покоящемся
мышечном волокне взаимодействие между актином и миозином предотвращают два белка - тропонин и тропомиозин.
Тропомиозин - стержневидная молекула из двух полипептидов, обвивающихся друг около друга; молекула соответствует в длину примерно семи мономерам актина. Цепи из молекул тропомиозина, уложенные конец в конец, располагаются вдоль всего тонкого филамента. Молекулы тропомиозина частично прикрывают участки связывания каждой молекулы актина, мешая контакту миозина с актином. В таком блокирующем положении молекула тропомиозина удерживается тропонином.
Тропонин - гетеротримерный белок. Он состоит из тропонина Т (отвечает з а с в я з ы в а н и е с о д и н о ч н о й
молекулой тропомиозина), тропонина С (связывает ион Са2+) и тропонина I (связывает актин и ингибирует сокращение). Каждая молекула тропомиозина связана с одной гетеротримерной молекулой тропонина, которая регулирует доступ к участкам связывания миозина на семи мономерах актина, прилегающих к молекуле тропомиозина.
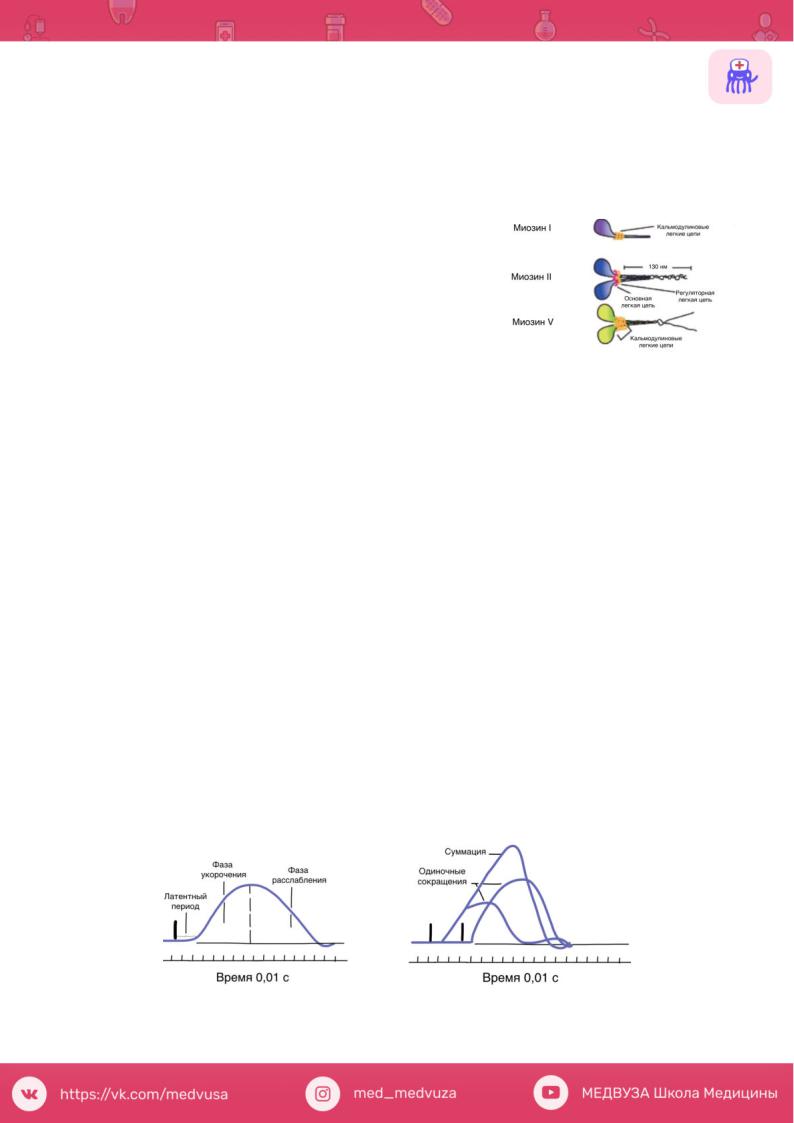
Миозин Это единое название большой семьи протеинов, имеющих определенные
отличия в клетках разных тканей. Миозин присутствует у всех эукариотов. В последнее время показано, что миозин II участвует в мышечном сокращении, тогда как миозин I и миозин V вовлечены в работу подмембранного (кортикального) цитоскелета. В настоящее время идентифицировано более 10 классов миозина.
На рисунке показаны схемы строения миозина, который состоит из головки, шейки и хвоста. Молекула миозина состоит из двух больших полипептидов (тяжелых цепей) и четырех меньших (легких цепей). Эти полипептиды составляют молекулу с двумя глобулярными «головками», которые содержат оба вида цепей, и длинным стержнем («хвостом») из двух
переплетенных тяжелых цепей. Хвост каждой молекулы миозина располагается вдоль оси толстого филамента, а две глобулярные головки выступают по бокам, их иначе называют поперечными мостиками. На каждой глобулярной головке находятся по два участка связывания: для актина и для АТФ. Участки связывания АТФ обладают также свойствами фермента АТФазы, гидролизующей связанную молекулу АТФ.
Суммация сокращений и ее виды. Тетанус и его виды. Оптимум и пессимум.
Мышечное сокращение является жизненно важной функцией организма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений осуществляются за счет скелетных мышц . Непроизвольные движения (кроме сокращения сердца) обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Одиночное мышечное сокращение. При раздражении мышцы одиночным импульсом тока возникает одиночное мышечное сокращение.
Амплитуда
Амплитуда одиночного сокращения мышцы зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных групп волокон различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается, несмотря на дальнейшее нарастание силы раздражающего тока.
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения При раздражении скелетной мышцы одиночным импульсом электрического тока
сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы:
•латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения;
•возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
•фаза укорочения (около 50 мс);
•фаза расслабления (около 50 мс).
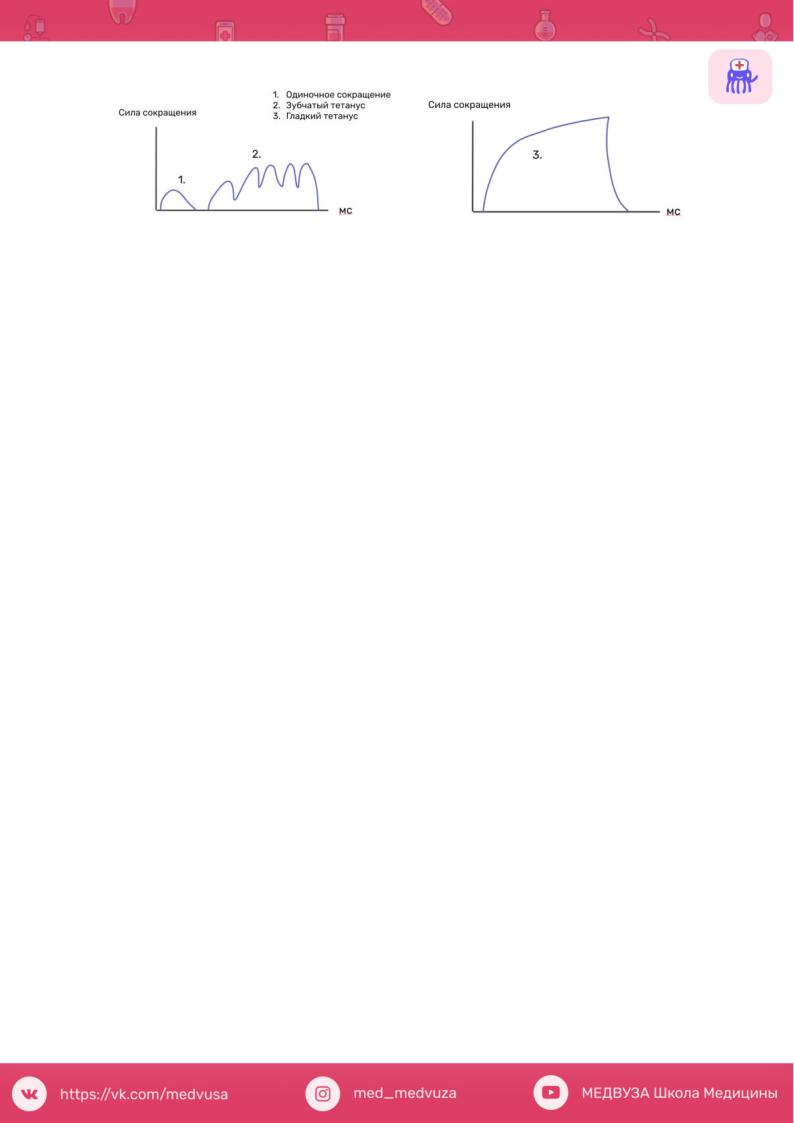
Тетанус, тетаническое мышечное сокращение — состояние длительного сокращения, непрерывного напряжения мышцы, возникающее при поступлении к ней через мотонейрон нервных импульсов с высокой частотой. При этом расслабления между последовательными одиночными сокращениями не происходит и возникает их суммация, приводящая к стойкому максимальному сокращению мышцы.
Виды тетануса, механизм возникновения различных видов тетануса:
Если последующее раздражение наносится, когда мышца уже начала расслабляться после предыдущего укорочения (то есть приходится на фазу
расслабления предыдущего цикла) -наблюдается зубчатый тетанус (неполный).