

1. Первый этап ознаменовался открытием Г. Менделем (1865) дискретности (делимости) наследственных факторов и разработкой гибридологического метода, изучения наследственности, т. е. правил скрещивания организмов и учета признаков у их потомства. Дискретность наследственности состоит в том, что отдельные свойства и признаки организма развиваются под контролем наследственных факторов (генов), которые при слиянии гамет и образовании зиготы не смешиваются, не растворяются, а при формировании новых гамет наследуются независимо друг от друга.
Значение открытий Г. Менделя оценили после того, как его законы были вновь переоткрыты в 1900 г. тремя биологами независимо друг от друга: де Фризом в Голландии, К. Корренсом в Германии и Э. Чермаком в Австрии. Результаты гибридизации, полученные в первое-I десятилетие XX в. на различных растениях и животных, полностью подтвердили менделевские законы наследования признаков и показали их универсальный характер по отношению ко всем организмам, размножающимся половым путем. Закономерности наследования признаков в этот период изучались на уровне целостного организма (горох, кукуруза, мак, фасоль, кролик, мышь и др.).
В 1901 —1903 гг. де Фриз выдвинул мутационную теорию изменчивости, которая сыграла большую роль в дальнейшем развитии генетики.
Важное значение имели работы датского ботаника В. Иоганнсена, который изучал закономерности наследования на чистых линиях фасоли. Он сформулировал также понятие “популяциям (группа организмов одного вида, обитающих и размножающихся на ограниченной территории), предложил называть менделевские “наследственные факторы” словом ген, дал определения понятий “генотип” и “фенотип”.
Второй этап характеризуется переходом к изучению явлений наследственности на клеточном уровне (питоге-нетика). Т. Бовери (1902—1907), У. Сэттон и Э. Вильсон (1902—1907) установили взаимосвязь между менделевскими законами наследования и распределением хромосом в процессе клеточного деления (митоз) и созревания половых клеток (мейоз). Развитие учения о клетке привело к уточнению строения, формы и количества хромосом и помогло установить, что гены, контролирующие те или иные признаки, не что иное, как участки хромосом. Это послужило важной предпосылкой утверждения хромосомной теории наследственности. Решающее значение в ее обосновании имели исследования, проведенные на мушках дрозофилах американским генетиком Т. Г. Морганом и его сотрудниками (1910—1911). Ими установлено, что гены расположены в хромосомах в линейном порядке, образуя группы сцепления. Число групп сцепления генов соответствует числу пар гомологичных хромосом, и гены одной группы сцепления могут перекомбинироваться в процессе мейоза благодаря явлению кроссинго-вера, что лежит в основе одной из форм наследственной комбинативной изменчивости организмов. Морган установил также закономерности наследования признаков, сцепленных с полом. 1911 Т.Морган сформулировал хромосомную теорию (гены в хромосомах)
Третий этап в развитии генетики отражает достижения молекулярной биологии и связан с использованием методов и принципов точных наук — физики, химии, математики, биофизики и др.—в изучении явлений жизни на уровне молекул. Объектами генетических исследований стали грибы, бактерии, вирусы. На этом этапе были изучены взаимоотношения между генами и ферментами и сформулирована теория “один ген — один фермент” (Дж. Бидл и Э. Татум, 1940): каждый ген контролирует синтез одного фермента; фермент в свою очередь контролирует одну реакцию из целого ряда биохимических превращений, лежащих в основе проявления внешнего или внутреннего признака организма. Эта теория сыграла важную роль в выяснении физической природы гена как элемента наследственной информации.
В 1953 г. Ф. Крик и Дж. Уотсон, опираясь на результаты опытов генетиков и биохимиков и на данные рентгеноструктурного анализа, создали структурную модель ДНК в форме двойной спирали. Предложенная ими модель ДНК хорошо согласуется с биологической функцией этого соединения: способностью к самоудвоению генетического материала и устойчивому сохранению его в поколениях — от клетки к клетке. Эти свойства молекул ДНК объяснили и молекулярный механизм изменчивости: любые отклонения от исходной структуры гена, ошибки самоудвоения генетического материала ДНК, однажды возникнув, в дальнейшем точно и устойчиво воспроизводятся в дочерних нитях ДНК. В последующее десятилетие эти положения были экспериментально подтверждены: уточнилось понятие гена, был расшифрован генетический код и механизм его действия в процессе синтеза белка в клетке. Кроме того, были найдены методы искусственного получения мутаций и с их помощью созданы ценные сорта растений и штаммы микроорганизмов — продуцентов антибиотиков, аминокислот.
В последнее десятилетие возникло новое направление в молекулярной генетике —генная инженерия — система приемов, позволяющих биологу конструировать искусственные генетические системы. Генная инженерия основывается на универсальности генетического кода: триплеты нуклеотидов ДНК программируют включение аминокислот в белковые молекулы всех организмов — человека, животных, растений, бактерий, вирусов. Благодаря этому можно синтезировать новый ген или выделить его из одной бактерии и ввести его в генетический аппарат другой бактерии, лишенной такого гена.
Таким образом, третий, современный этап развития генетики открыл огромные перспективы направленного вмешательства в явления наследственности и селекции растительных и животных организмов, выявил важную роль генетики в медицине, в частности, в изучении закономерностей наследственных болезней и физических аномалий человека.
2. Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком в 1958 году и приведено в соответствие с накопившимися к тому времени данными в 1970 году. Переход генетической информации последовательно от ДНК к РНК и затем от РНК к белку является универсальным для всех без исключения клеточных организмов, лежит в основе биосинтеза макромолекул. Репликации генома соответствует информационный переход ДНК → ДНК. В природе встречаются также переходы РНК → РНК и РНК → ДНК (например у некоторых вирусов), а также изменение конформации белков, передаваемое от молекулы к молекуле.
Биополимеры — это синтезируемые живыми существами биологические полимеры. ДНК, РНК и белки относятся к линейным полимерам, которые собираются путём последовательного присоединения друг к другу отдельных элементов — мономеров. Последовательность мономеров кодирует информацию, правила передачи которой описываются центральной догмой. Информация передаётся с высокой точностью, детерминистически и один биополимер используется как шаблон для сборки другого полимера с последовательностью, которая полностью определяется последовательностью первого полимера.
В живых организмах встречаются три вида гетерогенных, то есть состоящих из разных мономеров полимера — ДНК, РНК и белок. Передача информации между ними может осуществляться 3 × 3 = 9 способами. Центральная догма разделяет эти 9 типов передачи информации на три группы:
Общий — встречающиеся у большинства живых организмов;
Специальный — встречающиеся в виде исключения, у вирусов и у мобильных элементов генома или в условиях биологического эксперимента;
Неизвестные — не обнаружены.
Общие |
Специальные |
Неизвестные |
ДНК → ДНК |
РНК → ДНК |
белок → ДНК |
ДНК → РНК |
РНК → РНК |
белок → РНК |
РНК → белок |
ДНК → белок |
белок → белок |
Общие способы передачи информации
Репликация ДНК: ДНК → ДНК
ДНК — основной способ передачи информации между поколениями живых организмов, поэтому точное удвоение (репликация) ДНК очень важна. Репликация осуществляется комплексом белков, которые расплетают хроматин, затем двойную спираль. После этого ДНК полимераза и ассоциированные с ней белки, строят на каждой из двух цепочек идентичную копию.
Транскрипция: ДНК → РНК
Транскрипция — биологический процесс, в результате которого информация, содержащаяся в участке ДНК, копируется на синтезируемую молекулу мРНК. Транскрипцию осуществляют факторы транскрипции и РНК-полимераза. В эукариотической клетке первичный транскрипт (пре-мРНК) часто редактируется. Этот процесс называется процессингом РНК.
Трансляция: РНК → белок
Зрелая мРНК считывается рибосомами в процессе трансляции. В прокариотических клетках процесс транскрипции и трансляции не разделён пространственно, и эти процессы сопряжены. В эукариотических клетках место транскрипции — клеточное ядро — отделено от места трансляции (цитоплазмы) ядерной мембраной, поэтому иРНК транспортируется из ядра в цитоплазму. мРНК считывается рибосомой в виде трёхнуклеотидных «слов». Комплексы факторов инициации и факторов элонгации доставляют аминоацилированные транспортные РНК к комплексу мРНК-рибосома.
Специальные способы передачи информации
Обратная транскрипция: РНК → ДНК
Обратная транскрипция — перенос информации с РНК на ДНК, процесс, обратный нормальной транскрипции, осуществляемый ферментом обратной транскриптазой. Встречается у ретровирусов, например, ВИЧ и в случае ретротранспозонов.
Репликация РНК: РНК → РНК
Репликация РНК — копирование цепи РНК на комплементарную ей цепь РНК с помощью фермента РНК-зависимой РНК-полимеразы. Вирусы, содержащие одноцепочечную (например, пикорнавирусы, к которым относится вирус ящура) или двуцепочечную РНК реплицируются подобным способом.
Прямая трансляция белка на матрице ДНК: ДНК → белок
Прямая трансляция была продемонстрирована в клеточных экстрактах кишечной палочки, которые содержали рибосомы, но не иРНК. Такие экстракты синтезировали белки с введённых в систему ДНК, и антибиотик неомицин усиливал этот эффект.
Эпигенетические изменения — это изменения в проявлении генов, не обусловленные изменением генетической информации (мутациями). Эпигенетические изменения происходят в результате модификации уровня экспрессии генов, то есть их транскрипции и/или трансляции. Наиболее изученным видом эпигенетической регуляции является метилирование ДНК с помощью белков ДНК-метилтрансфераз, что приводит к временной, зависящей от условий жизни организма инактивации метилированного гена. Однако поскольку первичная структура молекулы ДНК при этом не изменяется, это исключение нельзя считать истинным примером передачи информации от белка к ДНК.
Основной фигурой матричных биосинтезов являются нуклеиновые кислоты РНК и ДНК. Они представляют собой полимерные молекулы, в состав которых входят азотистые основания пяти типов, пентозы двух типов и остатки фосфорной кислоты. Азотистые основания в нуклеиновых кислотах могут быть пуриновыми (аденин, гуанин) и пиримидиновыми (цитозин, урацил (только в РНК), тимин (только в ДНК)). В зависимости от строения углевода выделяют рибонуклеиновые кислоты – содержат рибозу (РНК), и дезоксирибонуклеиновые кислоты – содержат дезоксирибозу (ДНК).
Термин "матричные биосинтезы" подразумевает способность клетки синтезировать полимерные молекулы, таких как нуклеиновые кислоты и белки, на основе шаблона – матрицы. Это обеспечивает точную передачу сложнейшей структуры от уже существующих молекул к новосинтезируемым.
Основной постулат молекулярной биологии
В подавляющем большинстве случаев передача наследственной информации от материнской клетки к дочерней осуществляется при помощи ДНК (репликация). Для использования генетической информации самой клеткой необходимы РНК, образуемые на матрице ДНК (транскрипция). Далее РНК непосредственно участвуют на всех этапах синтеза белковых молекул (трансляция), обеспечивающих структуру и деятельность клетки.
На вышесказанном основана центральная догма молекулярной биологии, согласно которой перенос генетической информации осуществляется только от нуклеиновой кислоты (ДНК и РНК). Получателем информации может быть другая нуклеиновая кислота (ДНК или РНК) и белок.
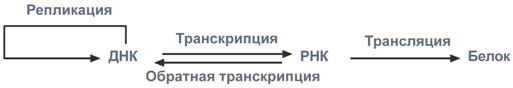
Центральная догма молекулярной биологии
3. Дезоксирибонуклеиновая кислота (ДНК) — универсальный носитель генетической информации и наследственных признаков у всех существующих на Земле организмов. Исключение составляют только некоторые микроорганизмы, например, вирусы — универсальным носителем генетической информации у них является РНК - одноцепочечная рибонуклеиновая кислота.
Открытие ДНК молекулы произошло в 1953 году. Френсис Крик и Джеймс Уотсон открыли структуру двойной спирали ДНК, их работа впоследствии была отмечена Нобелевской премией.
ДНК представляет собой двойную нить, скрученную в спираль. Каждая нить состоит из «кирпичиков» — из последовательно соединенных нуклеотидов. Каждый нуклеотид ДНК содержит одно из четырёх азотистых оснований — гуанин (G), аденин (A) (пурины), тимин (T) и цитозин (C) (пиримидины), связанное с дезоксирибозой, к последней, в свою очередь, присоединена фосфатная группа. Между собой соседние нуклеотиды соединены в цепи фосфодиэфирной связью, образованной 3’-гидроксильной (3’-ОН) и 5’-фосфатной группами (5’-РО3). Это свойство обуславливает наличие полярности в ДНК, т. е. противоположной направленности, а именно 5’- и 3’-концов: 5’-концу одной нити соответствует 3’-конец второй нити.
Первичная структура ДНК — это линейная последовательность нуклеотидов ДНК в цепи. Последовательность нуклеотидов в цепи ДНК записывают в виде буквенной формулы ДНК: например — AGTCATGCCAG, запись ведется с 5’- на 3’-конец цепи ДНК.
Вторичная структура ДНК образуется за счет взаимодействий нуклеотидов (в большей степени азотистых оснований) между собой, водородных связей. Классический пример вторичной структуры ДНК — двойная спираль ДНК. Двойная спираль ДНК — самая распространенная в природе форма ДНК, состоящая из двух полинуклеотидных цепей ДНК. Построение каждой новой цепи ДНК осуществляется по принципу комплементарности, т. е. каждому азотистому основанию одной цепи ДНК соответствует строго определенное основание другой цепи: в комплемнтарной паре напротив A стоит T, а напротив G располагается C и т.д.
Уникальным свойством ДНК является ее способность удваиваться (реплицироваться). В природе репликация ДНК происходит следующим образом: с помощью специальных ферментов (гираз), которые служат катализатором (веществами, ускоряющими реакцию), в клетке происходит расплетение спирали в том ее участке, где должна происходить репликация (удвоение ДНК). Далее водородные связи, которые связывают нити, разрываются и нити расходятся.
В построении новой цепи активным «строителем» выступает специальный фермент — ДНК-полимераза. Для удвоения ДНК необходим также стратовый блок или «фундамент», в качестве которого выступает небольшой двухцепочечный фрагмент ДНК. Этот стартовый блок, а точнее - комплементарный участок цепи родительской ДНК — взаимодействует с праймером — одноцепочечным фрагментом из 20—30 нуклеотидов. Происходит репликация или клонирование ДНК одновременно на обеих нитях. Из одной молекулы ДНК образуются две молекулы ДНК, в которых одна нить от материнской молекулы ДНК, а вторая, дочерняя, вновь синтезированная.
Таким образом, процесс репликации ДНК (удваивания) включает в себя три основных этапа:
Расплетение спирали ДНК и расхождение нитей
Присоединение праймеров
Образование новой цепи ДНК дочерней нити
Молекула ДНК человека — носитель генетической информации, которая записана в виде последовательности нуклеотидов с помощью генетического кода. В результате описанной выше репликации ДНК происходит передача генов ДНК от поколения к поколению.
Изменение последовательности нуклеотидов в ДНК (мутации) может приводить к генетическим нарушениям в организме.