

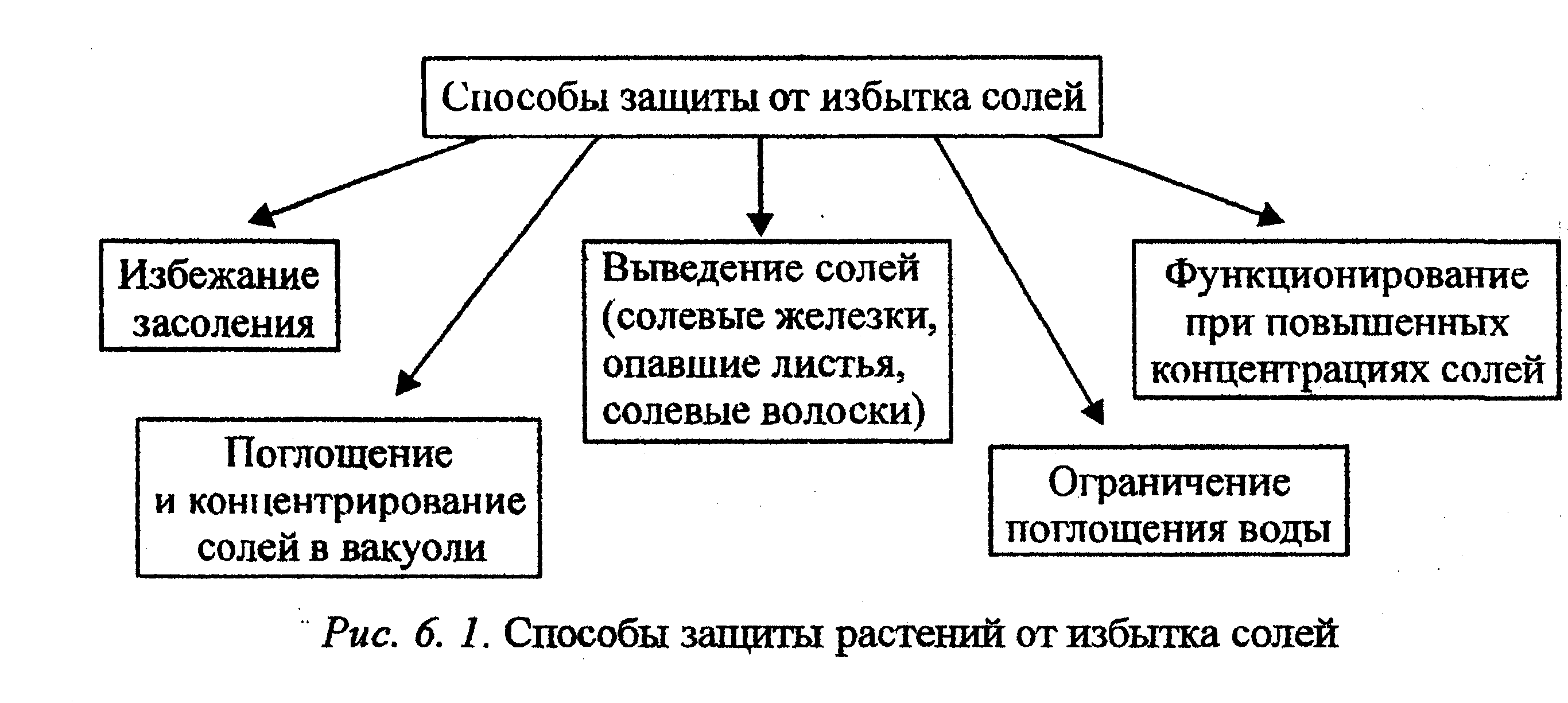
6. 1. 2. Особенности приспособления растений к условиям засоления
Концентрация солей выше 5% легальна для большинства растений. Однако существуют виды, адаптированные к произрастанию даже в условиях сильного засоления. При этом используется различная стратегия. Растения пытаются избежать накопления солей (даже при их интенсивном поступлении) (рис. 6. 1) или приспосабливаются к их высокой концентрации.
Избежание засоления
Для создания высокого осмотического давления и усиления поступления воды растения могут поглощать большое количество солей, однако для защиты цитоплазмы от избытка солей их необходимо концентрировать в вакуолярном соке.
Подобный способ увеличения осмотического давления достаточно эффективен в энергетическом отношении, так как высокое содержание ионов в почве при засолении создает пред-посылки для их пассивного поглощения по градиенту концентрации, т. е. без затрат АТФ.
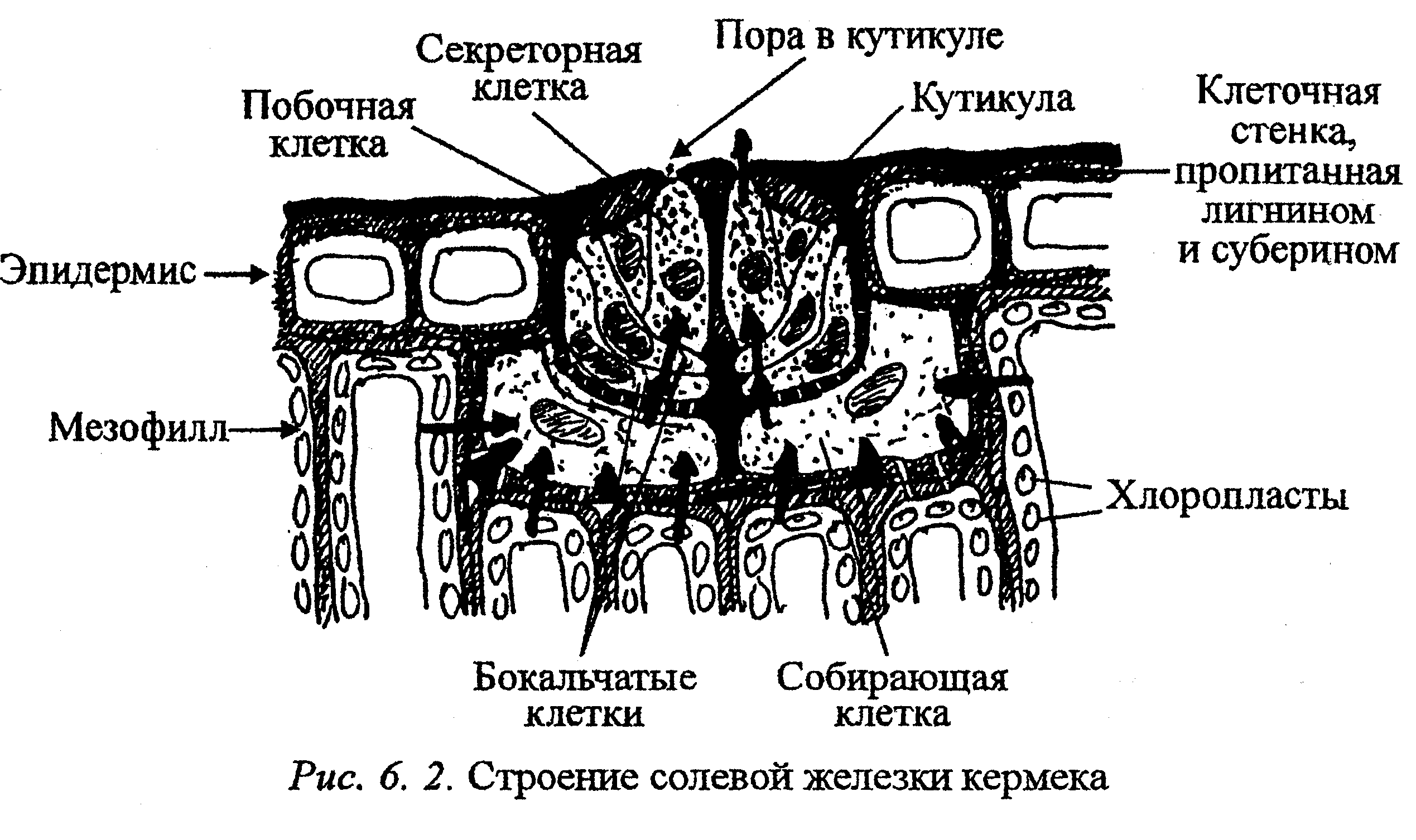
Поглощенные соли выводятся также из клеток с водой с помощью специализированных солевых железок, или избыток солей удаляется с опавшими листьями. Солевые железы листьев и стеблей растений выделяют избыток ионов при их росте в высокосолевой среде. Собирательные клетки железок мало чем отличаются от эпидермальных, но клеточная стенка у них значительно утолщена и лигнифицирована (рис. 6. 2). Эти клетки связаны многочисленными плазмодесмами с окружающими их клетками мезофилла и эпидермиса, а также с бокальчатыми клетками. Секреторные, побочные и бокальчатые клетки также связаны друг с другом через плазмодесмы, их клеточные стенки пропитаны не тольколигнином, но и суберином, что полностью исключает алопластный транспорт веществ. Кроме того, эти клетки железки находятся в своеобразной капсуле из лигнифицированной и суберинизированной клеточной стенки, которая таким образом изолируется от клеточных стенок окружающих клеток. Снаружи железы покрыты мощной кутикулой, которая в зоне секреторных клеток имеет поры. Через эти поры и происходит секреция солей. В секреторных клетках имеются многочисленные рибосомы, митохондрии, много мелких вакуолей, но нет хлоропластов и запасных веществ.
Соли поступают в корень, вместе с током воды движутся по сосудам ксилемы, а затем по апопласту тканей листа, что предохраняет цитоплазму клеток от избытка солей. В собирающих клетках при участии ионных насосов соли переходят в симпласг и до выделения наружу движутся по симпласту. В плазмалемме секреторных клеток активно функционируют Н+ и Сl-АТФазы. Благодаря деятельности этих и других насосов происходит активное выделение ионов хлора, натрия, кальция, магния, сульфата, нитрата. Помимо ионов через поры в кутикуле теряются значительные количества воды. В сухую погоду вода из выделенного раствора быстро испаряется и растение покрывается налетом солей, который затем сдувается ветром или смывается дождем.
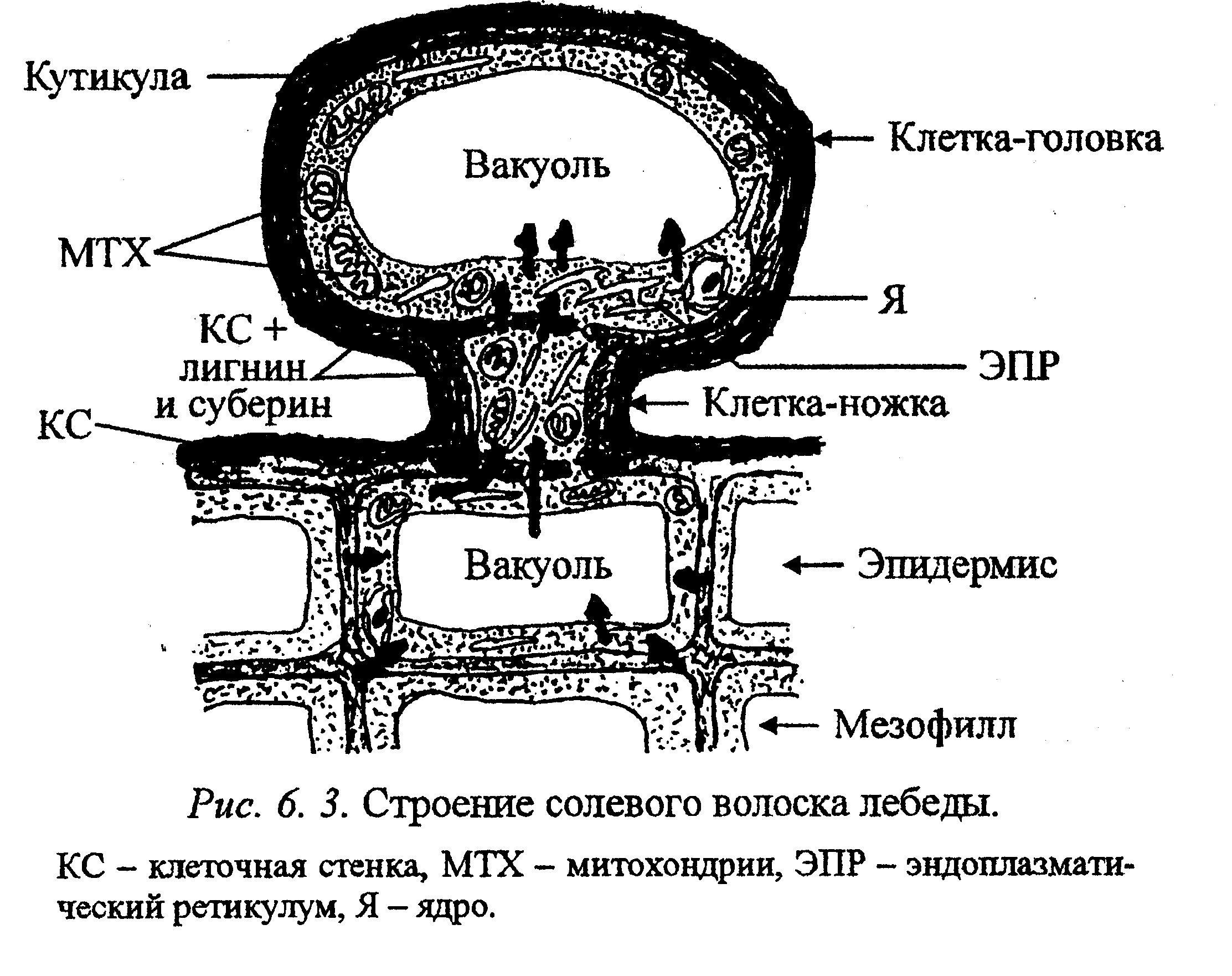
Поглощаемые соли могут концентрироваться и в вакуолях клеток головок специализированных волосков (рис. 6. 3). Когда в вакуолях го-ловки накапливаются соли, головка обрывается, на ее месте несколько раз в течение роста листа образуется новая головка.
Солевые волоски состоят из ножки и головки. В собирающей клетке-ножке, не имеющей центральной вакуоли, много митохондрий и цистерн ЭПР. В верхней секреторной клетке — головке, 95% объема цитоплазмы занимает центральная вакуоль. Ножка связана плазмодесмами с головкой и эпидермальными клетками. Клеточные стенки обеих клеток волоска утолщены и в значительной степени лигнифицированы, а также импрегнированы суберином, что препятствует передвижению солей из апопласта в симпласт. Снаружи волосок имеет толстую кутикулу, которая затрудняет испарение воды. При активном участии ионных насосов соли проникают в цитоплазму эпидермальных клеток, граничащих с клеткой-ножкой, перемещаются по симпласту в ножку и попадают в вакуоль клетки-головки. Накопление солей в клеточном соке происходит благодаря активному ионному транспорту на тонопласте. Концентрация солей в вакуоли головки может в 60 раз превышать таковую в клетках эпидермиса и эквивалентна 5-6 моль/л натрия и калия и 9-9,5 моль/л хлора. Вакуоль содержит также большое количество оксала-та. Кроме того, волоски выделяют катионы: Mg2+ и Са2+, и анионы: N03-, НСО3-, S042- Солевые волоски теряют очень мало воды, но эффективность выделения солей у них ниже, чем у солевых железок.
Уровень солей в цитоплазме уменьшается при сбрасывании старых листьев, через гидатоды вместе с секрецией воды (особенно если нет значительного водного дефицита). Локальное снижение уровня солей происходит и при выделении их в сосуды ксилемы и/или флоэмы и дальнейшем передвижении по растению.
Необходимо упомянуть и об ограниченном поглощении солей клетками корней вследствие соленепроницаемости мембран. Повышается специфичность ионных каналов, в основном калиевых, уменьшается их общее число. Кроме того, при засолении происходят структурные перестройки в липидах мембран, появляются пропитанные суберином эндодермальные клетки, которые тормозят поступление солей по апопласту. Однако мембраны не проницаемы для солей лишь при незначительном засолении, в случае сильного засоления соли проникают внутрь клеток. В этих условиях начинают действовать другие механизмы, направленные на активное выведение солей из цитоплазмы (в вакуоль или клеточную стенку).
Функционирование при повышенных концентрациях солей
Растения могут также приспосабливаться к высоким концентрациям солей. Такие растения, толерантные к засолению, называются гало-фитами (галос — соль, фитос — растение). Галофиты классифицируются следующим образом (рис. 6. 4).
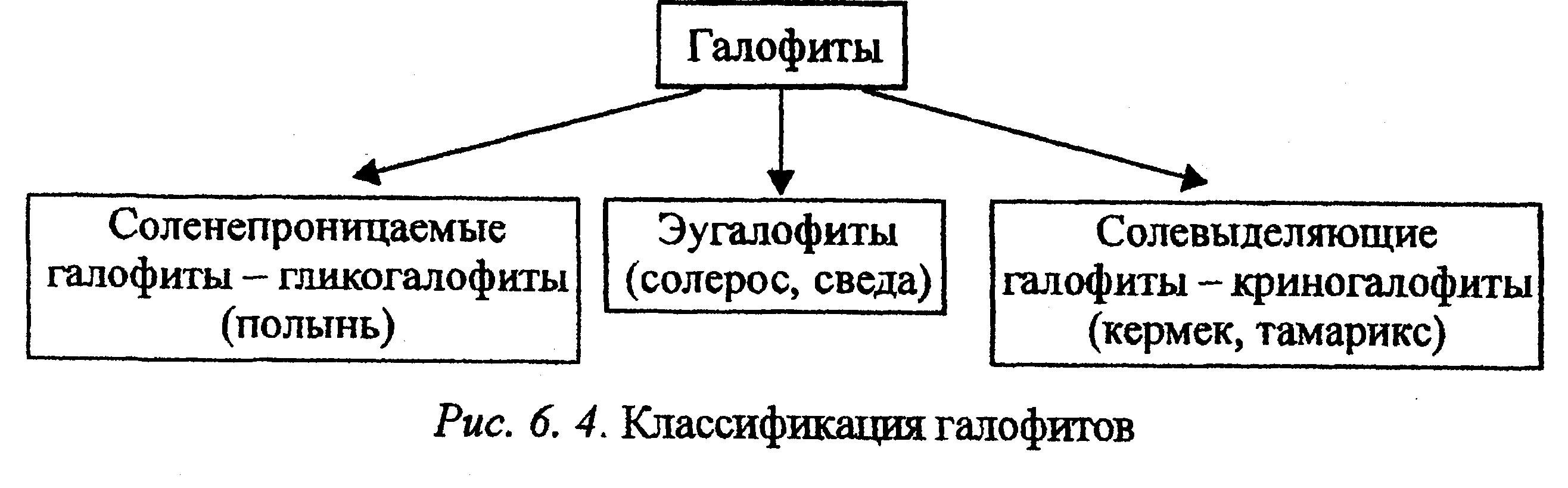
Эугалофиты (настоящие, или соленакапливающиё) — наиболее со-леустойчивые растения, накапливающие в вакуолях соли в значительных концентрациях. Растут на влажных засоленных почвах: на солончаках по морским побережьям и по берегам временных и постоянных соленых озер. Проницаемость мембран для солей у них повышена, и они накапливают соли до 10% (в несколько раз больше, чем в почве), т. е. представители этой группы галофигов нуждаются в высоком уровне солей для нормального развития. Вследствие высокого осмотического давления эугалофиты обладают большой сосущей силой и способны регулировать свое водоснабжение, поглощая воду из сильно засоленных почв. К эугалофитам относятся солерос (Salicornnia herbacea), сведа (Suaeda maritina).
Криногалофиты, или солевыводящие галофиты, благодаря хорошей проницаемости мембран поглощают соли, но накапливают их внутри тканей меньше, чем эугалофиты, так как способны выводить соли из клеток с помощью секреторных железок на листьях и стеблях. Выделение солей железками осуществляется с помощью ионных насосов и сопровождается транспортом большого количества воды. В сухую погоду криногалофиты покрываются налетом солей, который затем сдувается ветром или смывается дождем. Часть солей удаляется с опадающими листьями, т. е. приспособления этой группы галофигов включают и способы избежания засоления. В эту группу входят кермек (Statice gmelini), тамарикс (Tamarix speciosa) и др. Кроме того, к криногалофи-там относят и галофиты с мясистыми листьями — галосуккуленты. Привыращивании на незаселенных почвах галосуккулентность исчезает. Эугалофиты и криногалофигы называют солянками. У некоторых видов, например лебеды (Atriplex), при выделении солей в вакуоль клетки-головки волоска с последующим ее отбрасыванием и восстановлением избавление от избытка солей происходит без поглощения большого количества воды.
Гликогалофиты, или соленепроницаемые галофиты, растут на менее засоленных почвах, чем растения двух предыдущих групп. Высокое осмотическое давление в их клетках поддерживается за счет синтеза органических осмотиков, а мембраны клеток корня малопроницаемы для солей, т. е. значительного накопления солей в тканях гликогалофи-тов не происходит. К этой группе относятся различные виды полыни (Artemisia salina), обитающие в засоленных степях и полупустынях, а также некоторые другие виды.
Растения незасоленных водоемов и почв — гликофиты, в условиях засоления также обнаруживают определенную способность к перенесению избытка солей, и, следовательно, обладают возможностью приспосабливаться к повышенному уровню засоленности. Относительной устойчивостью обладают ячмень, сахарная свекла, хлопчатник, мягкая пшеница, в большей степени, чем твердая. Накапливаются соли в основном в старых тканях.
Увеличение концентрации солей в цитоплазме клеток служит сигналом к переходу обмена веществ на новый режим. Качественные изменения в метаболизме растений под влиянием солевого стресса однотипны у растений с разным уровнем устойчивости. Отмечаются лишь количественные различия скорости и амплитуды отклонения физиологических показателей от нормы. У приспособленных растений амплитуда изменений меньше, чем у неприспособленных.
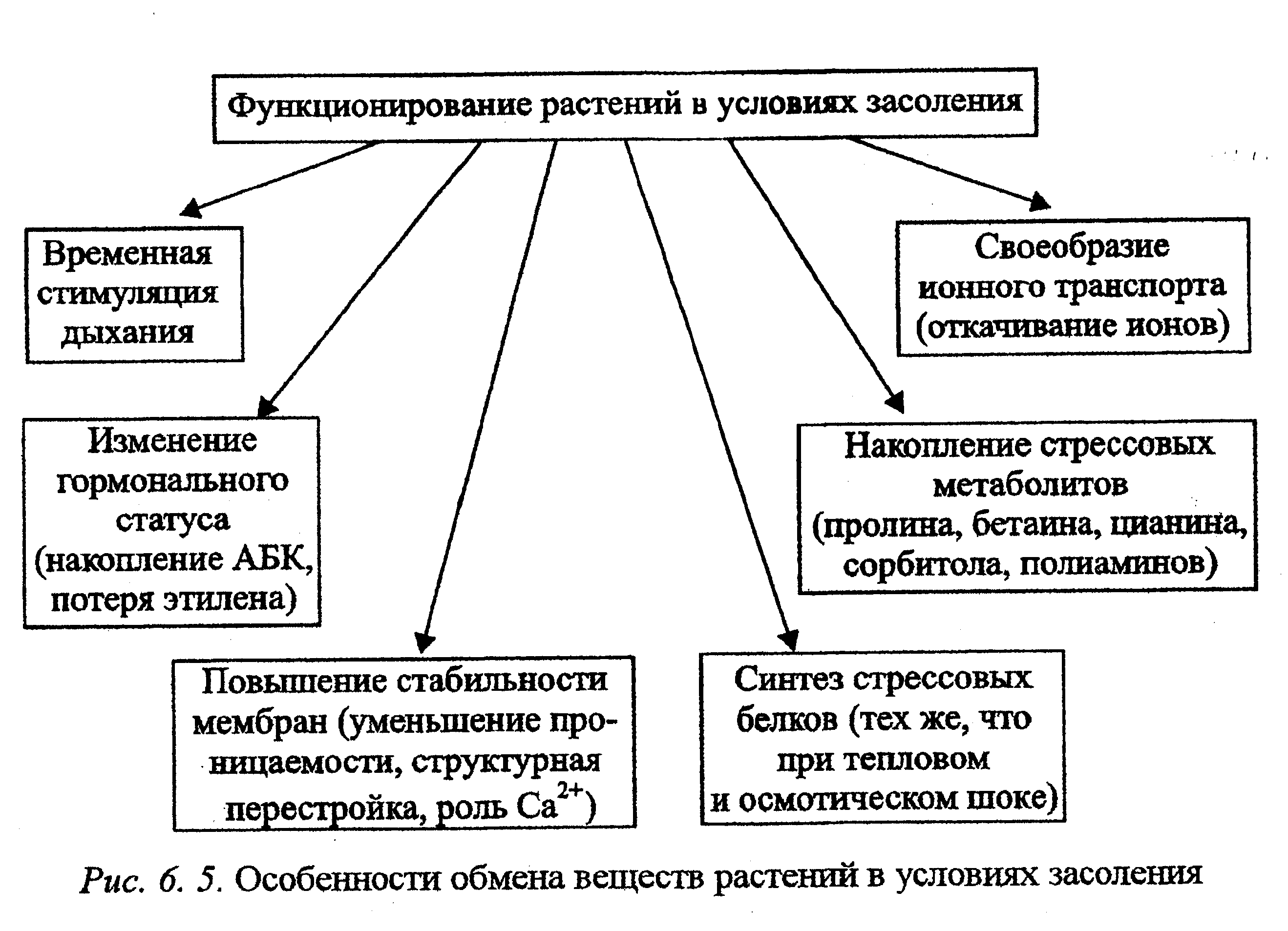
Особенности функционирования растений при избытке солей представлены на рис. 6.5. Изменение интенсивности дыхания при засолении выражается обычной параболической кривой, параметры которой зависят прежде всего от энергетических затрат на работу активного мембранного транспорта по выведению избытка ионов, а также на другие функциональные перестройки при солевом стрессе. Усиление дыхания необходимо и при недопущении поступления солей в клетку, поскольку структурные перестройки мембран, связанные с уменьшением проницаемости, изменением состояния белковых и липидных компонентов, также требуют затраты энергии. Помимо трансформации мембран энергия расходуется на образование анатомо-морфологических приспособлений, уплотнение покровов, суберинизацию эндодермы в корне, уменьшение транспирации листа.
Таким образом, реакция дыхания неоднозначна и изменяется в онтогенезе при различных концентрациях солей и в зависимости от типа приспособления галофитов. По уровню стимуляции дыхания основные ионы, определяющие засоление, можно расположить следующим образом: Na+ > К+ > S04 2-> Сl-. В присутствии солей меняется и дыхательный коэффициент (ДК). Поскольку в связывании ионов участвуют и органические кислоты, генерация которых возрастает, происходит увеличение отношения С02/02. Однако величина ДК может варьировать у разных растений.
Предохранению цитоплазмы от избытка солей способствует внутриклеточная компартментация ионов в вакоули, межклетниках. Создание такой разницы концентрации также требует серьезных затрат энергии. То же можно сказать о работе солевых волосков или железок, у которых интенсивность дыхания повышена, а также об образовании связывающих соли органических веществ — стрессовых метаболитов, если они синтезируются. У солеустойчивых растений интенсивность дыхания возрастает, а у менее устойчивых снижается в листьях и повышается в корнях и стеблях. В целом с солеустойчивостъю коррелирует уровень дыхания поддержания, отражающий способность организма к формированию защитных механизмов клеток.
В соответствии с накоплением стрессовых метаболитов — защитных веществ, галофиты группируют по признаку преобладания в них специфических продуктов нитратного обмена: пролина, бетаина и других метилированных аминокислот. При засолении у галофигов резко усиливается содержание именно тех соединений азотного метаболизма, которые они специфически накапливают. Содержание остальных групп азотных соединений при засолении меняется мало. Для галофигов характерно накопление полиаминов (в отличие от гликофитов, у которых содержание этих соединений снижается). Увеличение концентрации полиаминов связано с повышением активности декарбоксилаз и регулированием рН. Накопление осмопротекгоров коррелирует с засухо- и солеустойчивостью. Например, у хрустальной травки— Mesembryant-hemum crystallinnum, вида, обитающего в пустынях Намибии и обладающего устойчивостью к засухе, засолению и холоду, концентрация осмопротекгоров в цитоплазме и хлоропластах при стрессе значительно ниже, чем в специализированных эпидермальных клетках.
Накопление осмотиков происходит в результате стимуляции их синтеза, а также вовлечения в их образование продуктов распада полимеров. Роль осмотически активных соединений выполняют как неорганические соли, так и органические соединения. У эугалофитов минеральные соли могут обеспечивать накопление до 50-70% воды. Значительное содержание катионов в клетках поддерживается, в частности, за счет анионов органических кислот, которые образуются в цикле Кребса. Кроме того, органические кислоты участвуют в С4- или САМ-путях фотосинтеза, которые присущи многим солеустойчивым растениям.
В условиях солевого стресса большое значение приобретают особенности ионного транспорта. На уровне корня создаются мощные ион-транспортирующие барьеры. У соленепроницаемых галофигов такие барьеры хорошо выражены, поэтому галофиты этого типа испытывают только водный стресс, в то время как соленакапливающие— только солевой. Истинные галофиты, накапливающие соли, располагают молекулярными биологическими механизмами солеустойчивости и могут (особенно молодые растения) не иметь никаких ион-транспортирующих барьеров. Галофиты с другими типами солевого обмена обладают разными по мощности ионтранспортирующими барьерами и могут испытывать и водный, и солевой стресс. У гликофитов ион-транспортирующих барьеров нет. В их солеустойчивости решающую роль могут играть процессы водно-солевого обмена (рис. 6. 5).
Откачивание ионов из клетки в окружающую среду или в вакуоль осуществляется активно, с участием АТФаз. При избытке солей меняются свойства ионных каналов, определяющие степень солеустойчивости. У галофита проводимость канала больше, чем у гликофита. Под влиянием калия уменьшается Na-Mg-засоление.
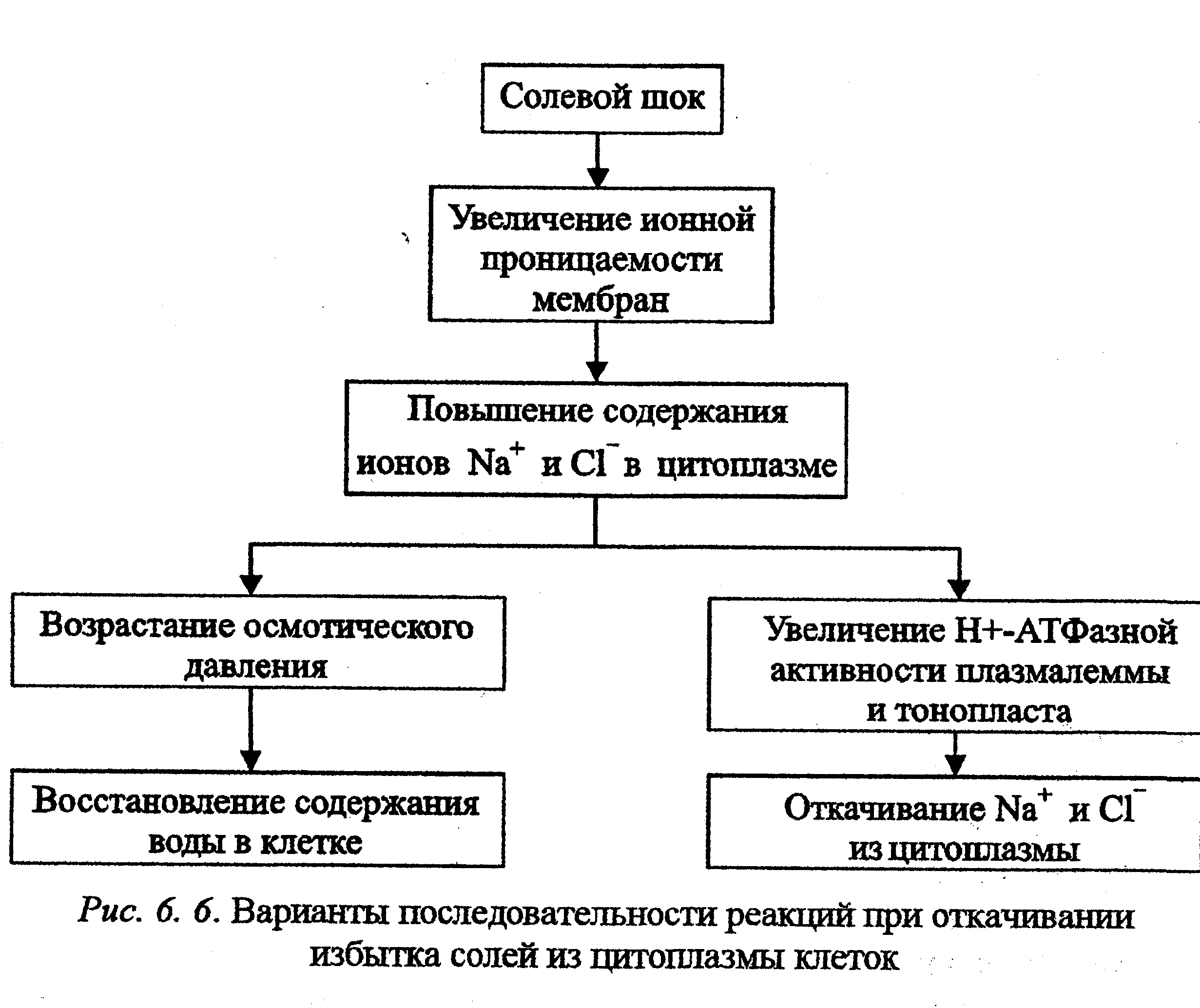
При засолении, особенно натрий-хлоридном, происходит значительное накопление соли в алопласте растительных клеток. Ионная проницаемость плазмалеммы повышается, поэтому хлор из клеточной стенки легко проникает в клетки через хлорные каналы и в симпорте с протоном, а натрий поступает через неспецифические катионные каналы, конкурируя с калием в калиевых каналах и на переносчиках. Уровень ионов хлора и особенно натрия в цитоплазме значительно увеличивается, что способствует активации систем ионного гомеостатирования. С одной стороны, повышается осмотическое давление, что ведет к восстановлению содержания воды в клетке. С другой стороны, возрастает активное протонных насосов плазмалеммы и тонопласта, в результате чего увеличивается вторично-активный обмен протонов на ионы натрия, которые откачиваются в вакуоль и в фазу клеточных стенок. На первых этапах засоления у эугалофитов может происходить пассивный транспорт натрия в вакуоль по градиенту концентрации. Кроме того, возрастает избирательность калиевых каналов и переносчиков для ионов калия, способствующая снижению поступления натрия (рис. 6. 6).
Система ионного гомеостаза у гликофитов при засолении работает достаточно эффективно. Так, клетки растений табака, выращенного на растворе хлористого натрия, 430 ммоль/л, содержали в клеточном соке в шесть-восемь раз больше ионов натрия и хлора, чем в цитоплазме. В ответ на засоление в цитоплазме значительно возрастает содержание не только натрия, но и кальция, являющегося вторичным посредником обмена веществ и участником трансдукции стрессового сигнала. Однако уровень кальция очень быстро снижается до исходного вследствие его закачивания в ЭПР и откачивания из клетки Са-насосами, а также в обмен на протоны.
У галофигов одна протонная помпа не может справиться с сильным засолением. В связи с этим представляют интерес данные, полученные на галотолерантных морских водорослях. На везикулах плазмалеммы этих водорослей обнаружено, что помимо Н+-АТФазы в плазмалемме функционирует еще один первичный натрий-транспортирующий насос — Na+-ATФaза. Транспорт ионов натрия сопровождается генерацией электропотенциала и встречным движением протонов. Na+-ATФaза работает как электрогенный унипортер, а выход Н+ из везикул осуществляется электрофоретически вследствие генерации потенциала на везикулярной мембране в ходе транслокации ионов натрия. Активность этих механизмов определяется солеустойчивостью организма и помимо величины электрохимического потенциала зависит от коэффициента проницаемости мембраны.
У галофитов (особенно у соленепроницаемых) при засолении повышается стабильность мембран, т. е. снижается их проницаемость. При обработке растений экзогенным кальцием ингибируется проницаемость мембран и возрастает активность АТФазы плазмалеммы, что играет защитную роль в стабилизации мембранных структур при засолении.
Один из механизмов солезащитного действия состоит в изменении гормонального статуса. Например, у растений, устойчивых к засолению, происходит блокирование ферментных систем образования этилена с одновременным увеличением содержания полиаминов. Поскольку, как отмечалось выше, предшественником биосинтеза и полиаминов, и этилена является 5-аденозилметионин, блокирование синтеза этилена приводит к активизации синтеза полиаминов. В результате устойчивость растения к засолению повышается (рис. 6. 5).
При действии высоких концентраций солей усиливается транскрипция генов низкомолекулярных стрессовых белков, тех же, что и при других стрессовых воздействиях, связанных с дефицитом воды. Так, в условиях засоления синтезируются белки-шапероны, LEA-белки, осмотин, аквалорин, различные гидролазы, в том числе и протеазы, ингибиторы протеаз, ферменты синтеза осмотиков, ферменты фиксации аммиака. Синтезируются ферменты антиоксидантной защиты. Галофи-ты отличаются высокой активностью пероксидазы и СОД.
Интенсификация работы протонных помп, ионных каналов и белков-переносчиков также происходит вследствие не только активации имеющихся систем, но и биосинтеза стрессовых белков — компонентов системы ионного гомеостаза: Н+-АТФазы, Nа+/Н+-антипортера и К+/Н+-симпортера ионных каналов. Отметим, что ферменты галофитных и гликофитных растений мало чем отличаются по солеустойчивости. От ингибирующего действия солей ферменты защищаются активной регуляцией ионного и водного гомеостаза клетки. Однако снижение активности ферментов при высоких концентрациях солей может быть компенсировано увеличением количества ферментного белка, что и происходит в результате синтеза стрессовых белков (рис. 6. 5).
В условиях засоления растений важно предотвратить не только накопление солей в цитоплазме, но и обезвоживание тканей. Поэтому га-лофиты должны обладать и приспособлениями к дефициту влаги. Как и в случае засухоустойчивых растений, у галофитов к таким приспособлениям относятся анатомо-морфологические изменения (ксероморф-ность): уменьшение размеров листьев, их опушенность; многослойный эпидермис; уменьшение количества устьиц; толстая кутикула; галосук-кулентность и т. п. Нормальная оводненность тканей достигается и за счет высокого содержания осмотически активных веществ, причем растениям каждого вида свойствен определенный преобладающий осмо-тик.
Таким образом, установлены многие общие механизмы устойчивости к избыточному засолению и засухе (а также и к высоким температурам). Поэтому устойчивость к одному из воздействий, вызывающих осмотический стресс, сопровождается резистентностью к другим подобным факторам среды. Действительно, изолированные солеустойчи-вые клетки табака имеют более высокую термотолерантность, а при температурном воздействии повышается устойчивость к повреждающим концентрациям хлористого натрия. О том же свидетельствуют сходные пути экспрессии синтеза белка при засухе и засолении (рис. 4. 9) или наборы стрессовых белков. Так, полипептид 24 кД для культуры
клеток табака и его целых растений, а также солеустойчивого сорта хлопчатника идентичен по молекулярной массе осмотину, который играет важную роль в адаптации растений не только к избыточному засолению, но и к засухе. Акватории, чей синтез активируется у арабидопсиса и при засухе, и при засолении, гомологичен у растений и животных. Введение мРНК аквалорина растений в ооциты Xenopis повышает их проницаемость для воды. Следовательно, у растений и животных существуют общие молекулярные механизмы поступления воды в клетки. Это не противоречит осморегуляторной функции вакуоли, в мембране которой также обнаружен акватории. Все сказанное свидетельствует о сходстве ответа клеток растений и других организмов на различные стрессовые воздействия.