

6a50ifFCQC
.pdfРис. 26. Схема «зоны пустоты», возникающей вокруг хищника в стае рыб (Мантейфель, 1980).
Многие птицы и млекопитающие используют разного рода сигналы тревоги, которые будучи пущены в ход, предупреждают других животных о том, что по близости находится хищник. Луговые собачки (отр. Грызуны), суслики, многие приматы, а также птицы, кормящиеся стаями, например, вороны и галки, нередко выставляют «часовых», наблюдающих за окрестностями с удобной позиции и в случае необходимости оповещающих группу о приближении хищника. У птиц особенно распространен крик тревоги как реакция на появление ястребов и соколов. Интересно предупреждают об опасности газели Томсона (Gazella thomsoni) – как только одна из них обнаруживает поблизости хищника, она на всех четырех ногах подпрыгивает высоко вверх. При приземлении раздается стук копытец о землю. Этот сигнал настолько выразителен, что газели замечают его даже на расстоянии километра (Вагнер, Шнейдерова, 1987).
Необходимо отметить, что в сообществах разных широт в характере межвидовых отношений имеются некоторые различия. Так, сообщества более низких широт характеризуются большим напряжением отношений «хищник-жертва» и соответственно усиленным развитием защитных приспособлений. Есть различия и по полушариям. В северотихоокеанской фауне напряженность отношений «хищник-жертва», видимо, больше, чем в фауне североатлантической (Никольский, 1963).
Таким образом, можно выделить четыре фактора, способствующих стабилизации взаимоотношений хищника и жертвы: 1) низкая эффективность хищника (основная причина – вышеперечисленные эволюционные приспособления жертвы); 2) экологические ограничения численности хищника (динамика численности жертв и другие факторы); 3) возможность перехода на другую пищу; 4) ускорение отклика популяции хищника на изменение численности жертвы (Константинов, 1986).
Хищники лимитируют популяцию жертвы, но в сформировавшемся сообществе это воздействие вряд ли можно считать отрицательным. Вопервых, хищник, как правило, действует избирательно, уничтожая слабых и больных животных. Во-вторых, избежавшие гибели животные часто проявляют реакции, которые компенсируют потери популяции (основная компенсация для большинства жертв – высокая скорость размножения). Эффективность такой компенсации можно проследить на следующем примере. Численность морских птиц достигает примерно 3 млрд. особей, из расчета их рационов (баклан потребляет 500–700 г корма в сутки, чайки – 200–300, чистиковые – 150–200, моевки – 108–140, крачки – 35–65) общее изъятие птицами из океана планктона, бентоса, головоногих моллюсков и рыбы, вероятно, превышает 1,5 млрд. ц (Шунтов, 1972; Белопольский, Шунтов, 1980). Это довольно большая цифра. Но от годовой продукции зоопланктона, бентоса и нектона, которая приблизительно оценена В. Богоро-
61
вым (1974) в 562 млрд. ц, она составляет менее 0,3%. Другими видами компенсации является снижение межвидовой конкуренции и увеличение внутривидовой.
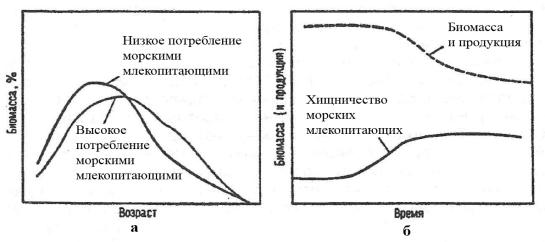
Хищник распространяет свое воздействие на трофическую нишу жертвы, уменьшая ее численность и создавая тем самым определенный избыток ресурсов. Этим он немало способствует активному размножению жертвы, изменению возрастной структуры популяции. Часто, истребляя избыточный молодняк, «отбивающий» корм у родителей, хищник поддерживает хороший физиологический статус особей, вынуждая их более активно двигаться, полнее использовать ресурсы трофической ниши (Рис. 27). Учитывая, что большинство хищников предпочитают охотиться на больных, старых или ослабленных особей, они выступают в роли своеобразных «санитаров». Сказанное, прежде всего, относится к так называемым расчетливым хищникам.
Рис. 27. Схема влияния хищничества морских млекопитающих на распределение биомассы вида-жертвы по возрастам (а) и на общую биомассу рыб (б) в экосистеме (По Левасту, Ларкинзу, 1987).
Ограничивая доступ вида-жертвы к части пищевых ресурсов и выбраковывая слабых, хищник сокращает пространственную нишу популяции (Рис. 28). Отсюда, вопреки устоявшемуся мнению (Бигон и др., 1989), хищничество увеличивает внутривидовую конкуренцию, т.к. способствует менее равномерному распределению жертв по обилию, т.е. «колоколизации» (сжиманию с боков) кривой Шелфорда.
62
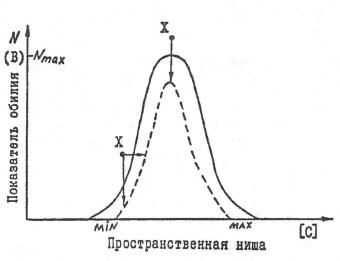
Рис. 28. Влияние хищничества (X) на изменение формы экологической ниши. Стрелками показано давление отбора.
Иными словами, выровненность представительства различных фенотипов (морфотипов) в популяциях под прессом хищников уменьшается. На примере короткоциклических видов можно хорошо проследить влияние хищничества на структуру популяции. Среди планктонных ракообразных озера Байкал весьма многочисленны так называемые шлемообразующие виды дафний (под/отр. Cladocera) Daphnia galeata и Daphnia cucullata. В их популяциях обычно встречаются два морфотипа – «нормальный» (или «типовой») и «шиповой» (Рис. 29). Шлемоносность рассматривают как эффективную защиту от мелких хищников – крупных кладоцер из рода Leptodora (Корзун, Питулько, 2010). Нормальный морфотип доминирует в популяциях при отсутствии хищников. Шип (вырост) на головном отделе кутикулы (панциря) дафний при отсутствии хищников только мешает рачкам, увеличивая их вес, затрудняя плавание, а значит, увеличивая энергетические затраты.
В обычной ситуации отбор действует в направлении подавления проявления данного признака. Однако при появлении в среде больших количеств хищной лептодоры происходит выедание мелких (до 1 мм) форм дафний, и в популяции начинают преобладать шлемоносный морфотип. Так, в Чивыркуйском заливе оз. Байкал при высокой плотности лептодоры (Leptodora kindti) частота встречаемости шипового морфотипа D. galeata составляла 87% у зрелых самок, 94% – у предзрелых и 78% – у ювенильных особей. При отсутствии лептодоры доля шипового морфотипа была равна соответственно 37, 45 и 64% (Корзун, Питулько, 2010). Из этих данных видно, что влияние хищника на популяцию дафний ведет к концентрации в популяции одного морфотипа, за счет избирательного выедания другого. При отсутствии хищника происходит выравнивание численности присутствия обоих морфотипов с преобладанием нормальных особей.
63

Рис. 29. Шиповой (а) и нормальный (б) морфотипы у D. galeata из оз. Байкал: Hgal – высота шлема, BL – длина тела.
Платой за хищничество часто является образование организованных ассоциаций и скоплений: стай, стад, колоний, табунов и т.п., являющихся эффективным механизмом противостояния хищнику, но характеризующихся напряженными внутривидовыми отношениями (Рис. 30).
Рис. 30. В большой колонии птиц гнездование (Гн) происходит более согласованно; в результате этого период Гн сокращается, а период воздействия хищника на наиболее уязвимую часть популяции (птенцов) уменьшается. Сокращение времени Гн аналогично сужению пространственной ниши под давлением хищника. С учетом повышения выживаемости птенцов вследствие уменьшения, как длительности, так и интенсивности воздействия хищника (защита птенцов всей колонией) общая численности птиц может расти, если позволяет трофическая ниша (Из Дре, 1976).
В сложных сообществах хищники поддерживают высокое видовое разнообразие: избирательно выедая наиболее многочисленные виды, они снижают межвидовую конкуренцию. Таким образом, хищничество и конкуренция часто оказываются комплементарными (Джиллер, 1988), т.е. взаимоуравновешивающими типами взаимодействия между организмами, совместно обеспечивающими стабильное существование сообщества.
64
Популяции самих хищников более подвержены действию внутривидовой конкуренции, поэтому многим из них так свойственно территориальное поведение (большинство кошачьих, медведи, хищные птицы). Когда в дело вмешиваются сильные межвидовые трения, например среди львов, гиен и гиеновых собак в африканской саванне, картина усложняется образованием ассоциаций даже у таких крупных кошачьих, как львы (прайды).
65

Множество паразитов обитает и на других животных. Многочисленны паразиты у рыб. На плотве (Rutilus rutilus) из озера Селигер было обнаружено 59 видов паразитов (Шульман, Чернышева, 1969), на окуне (Perca fluviatilis) из Ладожского озера – 35 видов (Румянцев и др., 1993), на молоди семги (Salmo salar) – 28 видов (Митенев, 1993), на трехлетней треске (Gadus morhua) – 31 вид паразитов из восьми систематических групп (Шульман и др., 1993) и т.д. Наибольшее значение, как паразиты рыб, имеют, пожалуй, веслоногие и ракообразные (отр. Copepoda). В фауне СССР насчи-ты- валось около 180 видов паразитических веслоногих (Маркевич, 1956). Из пресноводных веслоногих можно указать Ergasilus sieboldi, выдерживающего и небольшое осолонение воды. В Невской губе, по данным В.А. Догеля, этим рачком заражено около 13% щук и 6,5% плотвы. На одной рыбе зарегистрировано до 213 особей паразитов (Цит. по Никольскому, 1963).
Паразиты и патогены (болезнетворные агенты) являются крайне важной группой организмов. Каждый год миллионы людей гибнут от различных инфекций. Если сюда добавить болезни домашних животных и культурных растений, то урон с точки зрения человеческих страданий и экономических потерь становится неисчислимым. Конечно, люди, образуя плотные и скученные популяции, и создавая такие же условия для домашних животных и культурных растений, создают для паразитов и патогенов благоприятную обстановку. Есть основания считать, что вообще все животные и растения страдают и в огромных количествах гибнут от паразитов и болезней. Паразиты, безусловно, являются важным фактором смертности и снижения плодовитости во многих (может быть, в большинстве) природных популяциях, часто затрагивая все стороны их функционирования (Рис. 32).
При изучении влияния плероцеркоидов ленточного червя Ligula intestinalis на карповых рыб также показано, что паразит, развивающийся в полости тела рыб, влияет практические на все биологические функции и параметры организма хозяина (Извекова, 2010). Так, в результате влияния плероцеркоидов деформируется чешуя рыб, изменяются морфофизиологические показатели, происходит деградация печени, изменяется ее биохимический состав, наблюдается замедление в развитии гонад, приводящее к бесплодию, происходят значительные изменения в составе крови, в результате чего нарушается гуморальная регуляция, изменяется активность пищеварительных ферментов и интенсивность потребления кислорода. Все перечисленные функциональные нарушения приводят к серьезному изменению поведения рыб. Последнее сильно влияет на выживаемость зараженной части популяции, т.к. во многих случаях увеличивают доступность зараженных особей для хищников. Это механизм является эволюционно сложившимся и способствует проникновению паразита в организм окончательного (дефинитивного) хозяина, которым и являются хищники, поедающие жертву – промежуточного хозяина паразита.
67

Рис. 32. Зависимости между зараженностью клопа-водомерки Hydrometra myrae паразитическим клещом Hydraphantes tenuabilis и выживаемостью (А); продолжительностью жизни (Б); созреванием (В); плодовитостью (Г); скоростью роста популяции хозяина (Д) (Из Бигона и др., 1989).
Паразитизм и хищничество часто идут вместе и не всегда просто отделить одно явление от другого. Характерно в этом отношении высказывание Камерона (Cameron, 1958): «Хищники – паразиты расы, а паразиты – хищники особей. Они оба служат для сохранения баланса в природе» (Цит. по Краснощекову, 1995). Как мы уже условились ранее, многих кровососущих животных следует отнести к группе хищников (Дажо, 1976; Бигон и др., 1989): они достаточно самостоятельно регулируют свой обмен с окружающей средой. Многие из них являются переносчиками настоящих паразитов. Жизнь паразита, все его процессы и изменения, какие в нем происходят, в первую очередь обусловлены и зависят от организма хозяина. Эта особенность и является наиболее характерной и общей чертой всех истинных паразитов, отличающей их от всех других организмов и позволяющая объединить их в одну естественную группу (Догель, 1962). Однако явление паразитизма весьма разнообразно, и не всегда возможно провести резкую грань между категорией паразитических организмов и организмов, ведущих иной образ жизни (комменсалов, аменсалов и т.п.). Кроме того, существуют и многочисленные переходные формы.
В определенные периоды своего цикла паразит может вести совершенно свободную жизнь. Так, личинки блох (отр. Aphaniptera) относятся к сапрофагам, так как питаются различными органическими остатками поблизости от своих будущих хозяев, но очень редко на них самих. Такой паразитизм называют периодическим. Если же паразит связан с тем или
68

иным хозяином всю свою жизнь, и во внешней среде ни в какой стадии существовать не может, то такой паразитизм называют постоянным (Догель, 1962). Среди паразитов различают также эндопаразитов, живущих в теле своего хозяина и питающихся его тканями, соками или содержимым пищеварительного тракта (сосальщики-трематоды, цепни, двуустки, аскариды, трипаносомы, раффлезия и др.). Они гораздо лучше соответствуют обычным представлениям о паразитах, чем эктопаразиты, живущие, как правило, на поверхности хозяина и обладающие достаточной подвижностью, чтобы переходить от одного хозяина к другому (блохи, вши, пухоеды, большинство растений-паразитов и др.). Все ракообразные, паразитирующие на рыбах, также являются наружными паразитами, поражающими кожу или жабры рыб (Рис. 33).
Рис. 33. Ракообразные – паразиты рыб (слева направо): Argulus (отp. Branchiura); Caligus (отр. Copepoda); Aega (отр. Isopoda) (По Никольскому, 1963).
Паразитирующие организмы, могут быть факультативными (необязательными) паразитами, т.е. они могут при определенных условиях отказываться от паразитизма. Типичным факультативным паразитом является круглый червь Aloionema appendiculatum. Обыкновенно он является свободным обитателем почвы. Однако иногда личинки Aloionema проникают в слизня Arion ater, где и живут некоторое время в кишечнике, продолжая расти и достигая вдвое большей величины по сравнению с нормой. Впрочем, половозрелого состояния достигнуть в слизне они не могут и должны для этого выйти в окружающую среду, где и размножаются. В конечном счете факультативные паразиты составляют основу переходных форм. Напротив, облигатные паразиты, обязательно имеют хотя бы одну паразитирующую стадию в своем жизненном цикле.
Периодические паразиты. Периодический паразитизм может варьировать в зависимости от того, какой характер носит периодичность. В общем, периодический паразитизм встречается чаще постоянного (Догель, 1947).
69
Моногенетические сосальщики (класс Monogenoidea) – эктопаразитические плоские черви, питающиеся на коже или жабрах в первую очередь рыб, а также земноводных, пресмыкающихся, китообразных и головоногих моллюсков. Они удерживаются на теле хозяина с помощью специального прикрепительного органа на заднем конце тела. Свободноплавающие непаразитические личинки моногеней или сами взрослые особи активно разыскивают новых хозяев. Моногенея Gyrodactylus pterygialis – паразит, обладающий широкой экологической валентностью, что вообще мало свойственно для паразитов; его находили на разных рыбах (треска, навага, сельдь), обитающих в различных экологических условиях. Отмечено, что степень зараженности паразитом сильно зависит от условий обитания рыбы. При неблагоприятных условиях, близким к экстремальным (низкая соленость), степень зараженности трески из оз. Могильное (о. Кильдин), например, может достигать 93,1% особей, а количество паразитов до 200 штук на одну рыбу (Карасев и др., 1993). Такое явление можно связать с общим обеднением паразитофауны реликтовой кильдинской трески и снижением конкуренции среди паразитов (Догель, 1962).
Среди веслоногих ракообразных имеется своеобразное семейство Monstillidae. Во взрослом состоянии они быстро плавают в морском планктоне при помощи сильных гребных ног. Кишечник взрослых монстиллид атрофирован настолько, что они совсем не питаются. Из яиц оплодотворенных самок вылупляются личинки – науплиусы, лишенные пищеварительного канала. Они плавают, отыскивают определенные виды многощетинковых червей (Polychaeta) и проникают в них. В теле червя они проникают в спинной кровеносный сосуд, где и паразитируют, постепенно превращаясь во взрослого рачка, который затем покидает хозяина (Догель, 1962).
В других случаях паразит продолжительное время находится в покоящейся стадии, «поджидая» нового хозяина, и попадает в него с поглощением пищи и воды. Так это бывает с простейшим Entamoeba hystolytica, вызывающим амебную дизентерию. Подобным образом покоящиеся споры гриба Plasmodiophora brassicae могут, по-видимому, сохранять жизнеспособность в течение десятков лет. В присутствии подходящего хозяина они прорастают и проникают в корневые волоски капусты, после чего начинается цикл роста и размножения, заканчивающийся образованием на корнях наростов (Бигон и др., 1989).
Споры многих паразитических грибов также способны длительное время сохраняться на воздухе и в почве. Заселение листьев грибом (например, в случае мильдью на пшенице) требует контакта между спорами (обычно переносимыми ветром) и эпидермой, после чего гриб проникает в клетки хозяина или в межклеточные пространства, где начинает рост. Споры некоторых паразитических грибов, например спорыньи (Claviceps), могут переноситься насекомыми, привлекаемыми каплями сахаристой жид-
70