

Иннервация мочевого пузыря и расстройства мочеиспускания
В неврологической клинике нарушения функции тазовых органов (расстройства мочеиспускания, дефекации и половых органов) встречаются довольно часто.
Мочеиспускание осуществляется согласованной деятельностью двух мышечных групп: т. detrusor urinae и т. sphincter urinae. Сокращение мышечных волокон первой группы приводит к сжатию стенки мочевого пузыря, к выдавливанию его содержимого, что становится возможным при одновременном расслаблении второй мышцы. Происходит это в результате взаимодействия соматической и вегетативной нервной системы.
Мышцы, составляющие внутренний сфинктер мочевого пузыря и m. detrusor vesicae, состоят из гладких мышечных волокон, получающих вегетативную иннервацию. Наружный сфинктер мочеиспускательного канала образован поперечно-полосатыми мышечными волокнами и иннервируется соматическими нервами.
В акте произвольного мочеиспускания принимают участие и другие поперечно-полосатые мышцы, в частности мышцы передней брюшной стенки, диафрагмы дна таза. Мышцы брюшной стенки и диафрагмы при своем напряжении вызывают резкое повышение внутрибрюшного давления, что дополняет функцию m. detrusor vesicae.
Механизм регуляции деятельности отдельных мышечных образований, обеспечивающих функцию мочеиспускания, довольно сложен. С одной стороны, на уровне сегментарного аппарата спинного мозга существует вегетативная иннервация гладких волокон этих мышц; с другой стороны, у взрослого человека сегментарный аппарат подчиняется церебральной корковой зоне и этим осуществляется произвольный компонент регуляции мочеиспускания.
В акте мочеиспускания можно выделить два компонента: непроизвольно рефлекторный и произвольный.
Сегментарная рефлекторная дуга состоит из следующих нейронов (см рис.): афферентная часть — клетки межпозвоночного узла Si-Sm, дендриты заканчиваются в проприорецепторах стенки мочевого пузыря, входят в состав тазовых внутренностных нервов (nn. splanchnici pelvini), тазового нерва — nn. pelvici, аксоны идут в задних корешках и спинном мозге, контактируют с клетками
переднебоковой части серого вещества сегментов спинного мозга Si-Sm (спинальный центр парасимпатической иннервации мочевого пузыря).
Волокна этих нейронов вместе с передними корешками выходят из позвоночного канала и в составе тазового нерва (п. pelvicus) достигают стенки мочевого пузыря, где прерываются в клетках pi. vesicalis.
Постсинаптические волокна этих интрамуральных парасимпатических узлов иннервируют гладкие мышцы п. detrusor vesicae и частично внутренний сфинктер. Импульсы по этой рефлекторной дуге приводят к сокращению m. detrusor vesicae и расслаблению внутреннего сфинктера.
Схематически иннервацию мочевого пузыря можно изобразить следующим образом (см рис.1).
^-1
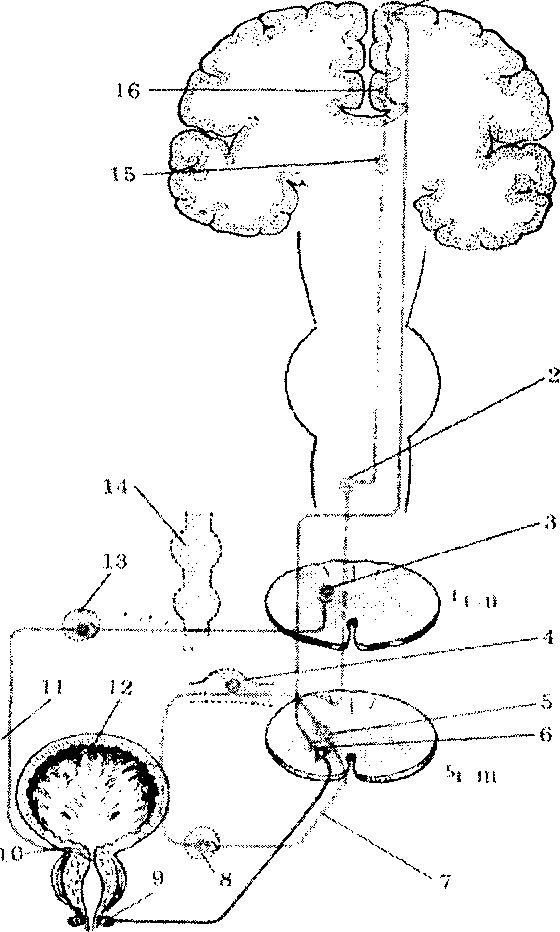
Рис. 1. Иннервация мочевого пузыря и его сфинктеров:
1 — пирамидная клетка коры парацентральной дольки; 2 — клетка ядра тонкого пучка; 3 — симпатическая клетка бокового рога Ьш; 4 — клетка спинномозгового узла; 5 — парасимпатическая клетка бокового рога Smii; 6 — периферический мотонейрон; 7 — половой нерв; 8 — пузырное сплетение; 9 — наружный сфинктер мочевого пузыря; 10—внутренний сфинктер мочевого пузыря; 11 — подчревный нерв; 12 — детрузор мочевого пузыря; 13 — нижний брыжеечный узел; 14 — симпатический ствол; 15 — клетка зрительного бугра; 16 — чувствительная клетка парацентральной дольки
Симпатические клетки, осуществляющие иннервацию мочевого пузыря, находятся на уровне Li-ц сегментов спинного мозга. Волокна этих симпатических нейронов вместе с передними корешками покидают позвоночный канал, затем отделяются в виде белой соединительной ветви и проходят, не прерываясь, через поясничные узлы симпатического ствола, в составе брыжеечных нервов достигают нижнего брыжеечного узла, где переключаются на следующий нейрон. Постсинаптические волокна в составе п. hypogastricus подходят к гладким мышцам мочевого пузыря. Автоматическое опорожнение мочевого пузыря обеспечивают две сегментарные рефлекторные дуги (парасимпатическая и соматическая). Раздражение от растяжения его стенок по афферентным волокнам тазового нерва передается в спинной мозг на парасимпатические клетки крестцовых сегментов спинного мозга, импульсы по эфферентным волокнам приводят к сокращению m. detrusor vesicae и расслаблению внутреннего сфинктера. Раскрытие внутреннего сфинктера и поступление мочи в начальные отделы мочеиспускательного канала включают еще
15
одну рефлекторную дугу для наружного (поперечно-полосатого) сфинктера, при расслаблении которого выделяется моча. Так функционирует мочевой пузырь у новорожденных детей. В дальнейшем в связи с созреванием надсегментарного аппарата вырабатываются и условные рефлексы, формируется ощущение позыва к мочеиспусканию. Обычно такой позыв появляется при повышении внутрипузырного давления на 5 мм рт. ст. Произвольный компонент акта мочеиспускания включает в себя управление наружным сфинктером уретры и вспомогательными мышцами (мышцы живота, диафрагма, диафрагма таза и др.).
Чувствительные нейроны заложены в межпозвоночных узлах Si-Sm. Дендриты проходят в составе срамного нерва и заканчиваются рецепторами как в стенке пузыря, так и в сфинктерах. Аксоны вместе с задними корешками достигают спинного мозга и в составе задних канатиков поднимаются до продолговатого мозга. Дальше эти пути следуют к gyrus fornicatus (сенсорная область мочеиспускания). По ассоциативным волокнам импульсы от этой зоны передаются на центральные двигательные нейроны, расположенные в коре парацентральной доли (двигательная зона мочевого пузыря расположена вблизи зоны стопы). Аксоны этих клеток в составе пирамидного тракта достигают клеток передних рогов крестцовых сегментов (Sn-Srv). Волокна периферических двигательных нейронов вместе с передними корешками покидают позвоночный канал, в полости малого таза образуют половое сплетение и в составе п. pudendus; подходят к наружному сфинктеру. При сокращении этого сфинктера удается произвольно удерживать мочу в мочевом пузыре.
При двустороннем нарушении связей церебральных (корковых) зон мочевого пузыря со спинальными его центрами (это бывает при поперечном поражении спинного мозга на уровне грудных и шейных сегментов) возникает нарушение функции мочеиспускания. Такой больной не чувствует ни позыва, ни прохождения мочи (или катетера) по мочеиспускательному каналу и не может произвольно управлять мочеиспусканием. При остро возникшем нарушении вначале наступает задержка мочи (retentio urinae); пузырь переполняется мочой и растягивается до больших размеров (дно его может доходить до пупка и выше); опорожнить его можно только при помощи катетера. В дальнейшем в связи с повышением рефлекторной возбудимости сегментарных аппаратов спинного мозга задержка мочи сменяется периодическим недержанием (incontinentio intermittens).
В более легких случаях наблюдаются императивные позывы на мочеиспускание.
При нарушении сегментарной вегетативной иннервации мочевого пузыря и сфинктеров возникают различные расстройства мочеиспускания.
Задержка мочи бывает при нарушении парасимпатической иннервации m. detrusor vesicae пузыря (сегменты спинного мозга Si-Siv, п. pelvicus).
Денервация внутреннего и наружного сфинктеров приводит к истинному недержанию мочи (incontinentia vera). Это возникает при поражении поясничных сегментов спинного мозга и корешков конского хвоста, п. hypogastricus и п. pudendus. В таких случаях больной не может удерживать мочу, она выделяется непроизвольно, либо периодически, либо непрерывно.
Встречается еще один тип расстройства мочеиспускания: парадоксальное недержание мочи (ischuria paradoxa), когда имеются элементы задержки мочи (пузырь постоянно переполнен, произвольно он не опорожняется) и недержания (моча все время вытекает по каплям вследствие механического перерастяжения сфинктера).