

Енергетика сперміїв
Джерелом енергії для руху сперміїв є три біохімічні процеси: дихання, гліколіз та розпад АТФ. Ці процеси взаємозв'язані: внаслідок перших двох виділяється енергія, а
124
Сперма. Фізіологія та біохімія сперми
третій зводиться до перетворення цієї енергії в таку форму, яка може діяти на руховий апарат спермія. Названі три процеси екзотермічні, тобто супроводжуються виділенням енергії, їх можна зобразити такими рівняннями:
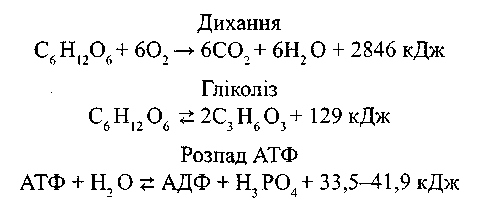
Друга і третя реакції - зворотні і можуть йти як зліва направо, так і справа наліво за рахунок енергії, виділеної у першій реакції. При цьому АДФ, що утворилася під час руху спермія, переходить у АТФ за рахунок енергії дихання чи гліколізу, а накопичена під час гліколізу молочна кислота може назад відновлюватись за рахунок енергії дихання до фруктози.
Акумулятором утворюваної енергії служить АТФ, молекула якого складається з аденозину та трьох частин фосфорної кислоти. Остання частка фосфорної кислоти приєднана за допомогою макроергічного зв'язку; при розриві цього зв'язку виділяється енергія (33,5-41,9 кДж/моль) і АТФ перетворюється у АДФ. Таким же чином остання може перетворюватися у АМФ.
В. А. Енгельгарт і С. А. Бурнашова виділили з сперміїв бугая скоротливий білок епермозин, що володіє властивістю ферменту аденозинтрифосфатази. Спермозин зосереджений переважно у хвостовій частині спермія, його зовнішніх дев'яти фібрилах. При його дії на АТФ руйнується макроергічний зв'язок, а виділювана енергія, що передається спермозину, перетворюється з хімічної у механічну роботу, що проявляється рухом спермія.
Коли макроергічні зв'язки у АТФ та АДФ вичерпуються, скорочення волокенець спермія та його рух припиняються. Щоб рух сперміїв продовжувався АТФ і АДФ повинні відновитися. Насправді так і є, ці реакції обратимі. Для ресинтезу АТФ з АДФ або АДФ в АТФ потрібен органічний фосфат та енергія, що виділяється у процесі дихання та гліколізу. Ця енергія захоплюється молекулою АТФ у вигляді макроергічних зв'язків, які розриваються під впливом спермозину, і вивільнювана енергія передається скоротливому білку, який при цьому скорочується і обумовлює рух спермія. Завдяки диханню та гліколізу вміст АТФ у сперміях утримується приблизно на одному рівні. Якщо позбавити спермії цих процесів (помістивши у безкисневі умови та безцукрове середовище), то концентрація АТФ у них швидко впаде і рух сперміїв припиниться.
Головними субстратами для дихання та гліколізу є цукри, оскільки вони легко проникають крізь оболонку сперміїв і піддаються розщепленню з виділенням енергії. Одержання енергії для життєвих процесів шляхом розщеплення цукру у безкисневих
125
Розділ 4
умовах називається гліколізом. Оскільки головним цукром сперми є фруктоза, то цей процес ще називають фруктолізом; інтенсивність гліколізу (фруктолізу) залежить від концентрації цукру.
У спермі бугая та барана міститься значно більше цукру, ніж у спермі кнура та жеребця, тому його розщеплення тут набагато інтенсивніше. У спермі ж кнура та жеребця переважає дихання, а розклад цукру тут мало помітний. Враховуючи це М. П. Шер-гін розділив сперму сільськогосподарських тварин на два типи. До першого належить сперма з високим вмістом цукру та добре вираженим гліколізом (фруктолізом). Це -сперма бугая, барана, цапа, оленя. Характерною рисою сперми цього типу є малий об'єм еякуляту, висока концентрація сперміїв, кисла реакція та виражена здатність сперміїв впадати у анабіоз. До другого типу належить сперма кнура та жеребця, яка характеризується великим об'ємом еякуляту, низькою концентрацією у ній сперміїв, малим вмістом цукру, лужною реакцією і слабкою здатністю впадати в анабіоз.
Майже всі проміжні реакції гліколізу супроводжуються звільненням невеликої кількості енергії, що разом складає 129 кДж на ммоль фруктози. Якщо би така кількість енергії звільнялася в результаті однієї реакції, то це викликало б теплове пошкодження структур сперміїв і їх загибель. Поступове ж виділення енергії захищає клітини від перегрівання.
Усі проміжні реакції гліколізу супроводжуються фосфорилюванням, яке охороняє виділювану при цьому енергію від розсіювання. Без фосфатів вона виділялась би у вигляді теплової енергії, нездатної викликати скорочення джгутика.
Гліколіз може відбуватися як у анаеробних, так і в аеробних умовах, проте на значно нижчому рівні. Кінцевим продуктом гліколізу є молочна кислота, яка, залежно від її концентрації, діє по-різному на спермії. Спочатку вона гальмує гліколіз і викликає у сперміїв стан неглибокого анабіозу, збільшуючи при цьому їх виживання. Під мікроскопом такі спермії (в стані неглибокого анабіозу) виглядають нерухомими, але, якщо помістити їх на обігрівальний столик, то вони починають рухатися. Тому при оцінці сперми користуються мікроскопом з обігрівальним столиком. Лри дальшому накопиченні молочної кислоти наступає друга фаза анабіозу, коли одного підвищення температури вже недостатньо для активування сперміїв: необхідно додати до краплі сперми на предметному склі краплю лугу. Нарешті, при значному накопиченні молочної кислоти і зміщенні рН сперми нижче 5,7 більшість сперміїв гине.
Перебуваючи у стані анабіозу, спермії наче зберігають енергію, їх протоплазма змінюється мало, їх живучість збільшується. Проте подібний анабіоз не настільки глибокий, щоб припинити шкідливу дію на протоплазму сперміїв молочної кислоти, і з часом все менший процент сперміїв може відновлювати рухливість (на відміну від низькотемпературного анабіозу).
Важливу роль у життєдіяльності сперміїв відіграють аеробні процеси. Під окисленням розуміють, звичайно, або приєднання кисню, або втрату двох атомів водню, або ж перенесення електронів даною сполукою. При диханні внаслідок більш глибокого розщеплення субстрату спермії одержують значно більше енергії, ніж при гліко-
126
Сперма. Фізіологія та біохімія сперми
лізі і тому затрати поживного матеріалу, і виділення продуктів розпаду відносно малі. Для одержання рівної кількості енергії при гліколізі повинно розщепитися майже у 29 разів більше енергетичної речовини, ніж при диханні.
Але, як і при гліколізі, не вся енергія, що звільнюється при диханні, перетворюється у енергію макроергічних зв'язків АТФ; можливе і "вільне окислення", при якому енергія, що звільнюється0 при диханні, не передається на фосфорилюючі системи, а виділяється у вигляді тепла.
Субстратом дихання можуть бути цукри, ліпіди, білки, проте в першу чергу окислюються прості цукри (моноцукри - фруктоза та глюкоза); можуть також використовуватися галактоза, сахароза, мальтоза, арабіноза, лактоза, сорбіт, а також молочна кислота, що утворюється при гліколізі.
При відсутності цукрів у аеробних умовах спермії можуть використовувати для дихання внутрішньоклітинні субстрати, головним чином фосфоліпіди і у меншій мірі білки та жири. Окислення субстрату до вуглекислого газу і води відбувається за участю ферментних систем, що засвоюють кисень з повітря. Такими системами у спермі с флавопротеїни та залізовмісні цитохроми.
Дихання і гліколіз взаємозв'язані між собою, перші їх стадії навіть є спільними. Тому збільшення окислювальних процесів (при доступі кисню) гальмує гліколіз, і навпаки. Це явище назване ефектом Кребтрі.
ь
Вище відмічалось, що в анаеробних умовах спермії бугая та барана швидко впадають у стан анабіозу і тому довше зберігають свою життєздатність, тоді як у аеробних умовах вони активно рухаються і тому швидко гинуть. Спермії можуть також використовувати для окислення фосфоліпіди, жирні кислоти, гліцерин і навіть амінокислоти (Ь-триптофан, Ь-фенілалалін та Ь-тирозин); останні окислюються до аміаку.