


Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
МУЛЬТИСТАБИЛЬНОСТЬ В ПРОСТЕЙШИХ МОДЕЛЯХ ЭВОЛЮЦИИ ЕСТЕСТВЕННЫХ ПОПУЛЯЦИЙ
Жданова О.Л. 1,2, Фрисман Е.Я.2
1ФГБУН Институт автоматики и процессов управления ДВО РАН, Владивосток, Россия axanka@iacp.dvo.ru
2ФГБУН Институт комплексного анализа региональных проблем ДВО РАН, Биробиджан, Россия
frisman@mail.ru
Аннотация: Исследуется динамика численности и генетического состава однородной лимитированной популяции с плотностно-зависимым r-K отбором в одном диаллельном локусе. Проводится подробное изучение механизмов потери устойчивости рассматриваемой модели. Показано, что в достаточно обширных биологически содержательных областях параметров возможно одновременное существование нескольких различных предельных режимов, каждый из которых имеет свой бассейн притяжения.
1. Введение
Детальные исследования простейших моделей динамики численности, появившиеся в начале 70-х гг. прошлого века (Шапиро, 1972; May, 1975) показали, что динамика таких популяций, может быть неожиданно сложной. Обнаруженный недавно (Безручко и др., 2002; Кузнецов и др., 2012; Кулаков и др., 2014; Неверова, Фрисман, 2015; и др.) новый эффект нелинейной динамики - мультистабильность выражается в одновременном существовании в системе различных предельных режимов динамики, переход к которым определяется выбором начальных условий, привлек интерес исследователей, т.к. помимо удивительности самого феномена, позволяет объяснить случаи смены динамических режимов, наблюдаемые в реальных популяциях (напр., Kausrud et al., 2008; White, 2011).
2.Модель однородной популяции с r-K отбором по одному диаллельному локусу
Вработе исследуются результаты действия естественного отбора по одному адаптивному признаку в однородной популяции диплоидных организмов:
|
|
xn+1 |
= |
|
n xn |
|
|
|
|
|
|
w |
|
|
|
|
(1) |
||||
|
= qn (wAAqn |
+ 2wAa (1 |
|
|
|
|
||||
qn+1 |
− qn )) / wn |
|||||||||
где n – это |
номер |
поколения, |
|
n = wAAqn2 + 2wAa qn (1− qn ) + waa (1− qn )2 - средняя |
||||||
w |
||||||||||
приспособленность популяции в n-ом поколении, xn - численность n-ого поколения, qn – частота аллеля A в n-ом поколении, wAA(n), wAa(n), waa(n) – приспособленности генотипов AA, Aa и aa соответственно в n-м поколении. Предполагается экспоненциальный вид
зависимости приспособленности от численности популяции: wij = exp(Rij (1− x / Kij )) .
Мальтузианский параметр (Rij) характеризует репродуктивный потенциал популяции или скорость размножения популяции в пустоту (т.е. в отсутствие каких-либо ограничений по ресурсам). Ресурсный параметр (Kij) – это равновесное значение численности, которое было бы достигнуто популяцией, если бы она состояла только из особей с ij-м генотипом. Описаны результаты многих наблюдений r-и K-стратегий отбора в лабораторных и природных популяциях как низших, так и высших организмов (Long, Long, 1974; MacNaughton, 1975; Graham, 1977).
Аналитическое исследование модели (1) позволило получить условия существования и устойчивости всех ее нетривиальных стационарных точек (таблица 1), (Жданова, Фрисман, 2005). Анализ представленных результатов показывает, что хотя в общем и можно классифицировать динамику численности и генетического состава рассматриваемой
60

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
популяции на основе значений внутрипопуляционных параметров (Kij и Rij), немаловажную роль в определении направления эволюции и характера динамики популяции имеют начальные условия. Так в модели (1) возможен полиморфизм в параметрической области, где существуют только мономорфные равновесия и нет полиморфного; кроме того, возможно вытеснение оптимального по ресурсному параметру генотипа (при неблагоприятных для него начальных условиях).
Таблица 1 – Условия существования и устойчивости нетривиальных стационарных точек модели (1)
Стационарная точка |
|
|
существование |
|
устойчивость |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|||
Мономорфная 1: q = 0, x = Kaa |
|
|
|
|
|
Raa > 0 и Kaa > 0 |
0 |
< Raa < 2, Kaa > KAa |
|||||
Мономорфная 2: q = 1, x = KAA |
|
|
|
|
RAA > 0 и KAA > 0. |
0 |
< RAA < 2, KAA > |
||||||
|
|
|
|
|
|
|
|
|
|
KAа |
|||
(w |
Aa |
(x) −1)2 = |
(w |
AA |
(x) −1)(w |
(x) −1) |
KAa < min (КАA, |
KAa > max (КАA, Кaa) |
|||||
|
|
|
|
|
aa |
|
|||||||
Полиморфная: |
|
q = |
|
1 − wAa (x) |
|
|
Кaa) или KAa > |
− 2 < x |
|
x′ < 0 |
|||
|
|
|
|
|
|
|
|
max (КАA, Кaa) |
w |
||||
|
wAA (x) − wAa (x) |
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|||||
В работе исследуются механизмы потери устойчивости и характер сложного динамического поведения генетической структуры и численности популяции. Строятся бассейны притяжения одновременно существующих предельных динамических режимов; таким образом производится оценка влияния начальных условий на направление эволюции естественной популяции.
Благодарности
Работа выполнена при частичной поддержке комплексной программы фундаментальных исследований Дальневосточного отделения РАН "Дальний Восток" и
РФФИ (проект 15-29-02658).
Литература
Безручко Б.П., Прохоров М.Д., Селезнев Е.П. Виды колебаний, мультистабильность и бассейны притяжения аттракторов симметрично связанных систем с удвоением периода // ПНД. – 2002. Т. 10, №4. - С.47 – 67.
Жданова О.Л., Фрисман Е.Я. Динамические режимы в модели однолокусного плотностно-зависимого отбора // Генетика. - 2005. Т. 41, № 11. - С.1575 - 1584.
Кулаков М.П., Неверова Г.П., Фрисман Е.Я. Мультистабильность в моделях динамики миграционно-связанных популяций с возрастной структурой // Нелинейная динамика. – 2014. Т. 10, № 4. – С.407 - 425.
Кузнецов А.П., Савин А.В., Седова Ю. В., Тюрюкина Л. В. Бифуркации отображений. Саратов: Наука, 2012. - 196 с.
Неверова Г.П., Фрисман Е.Я. Сравнительный анализ влияния различных типов плотностной регуляции на динамику численности структурированных популяций // Информатика и системы управления. – 2015.
Т. 43, № 1. – С.41 - 53.
Шапиро А.П. К вопросу о циклах в возвратных последовательностях // Управление и информация. - 1972. Вып.
3. - С.96 - 118.
Graham J. Reproductiv effect and r- and K-selection in two species of Lacuna (Gastropods: Prosobranchia) // Mar. Biol.
– 1977. V.40, № 3. P.217 - 224.
Kausrud K. L., Mysterud A., Steen H., Vik J. O., Ostbye E., Cazelles B., Framstad E., Eikeset A. M., Mysterud I., Solhoy T., and Stenseth N.C. Linking climate change to lemming cycles // Nature. – 2008. V. 456. - P.93 – 97.
Long Th., Long G. The effects of r- and K-selection on components of variance for two quantitative traits // Genetics. – 1974. V. 76, № 3. P.567 - 573.
MacNaughton S.J. r- and K-selec. in Tipha // Amer. Natur. – 1975. V.109, № 961. P. 251 - 261.
May R.M. Biological population obeying difference equations: stable points, stable cycles, and chaos // J. Theor. Biol. - 1975. V.51, № 2. - P.511 - 524.
White T.C.R. What has stopped the cycles of sub-Arctic animal populations? Predators or food? // Basic Appl. Ecol. – 2011. V. 12. - P.481 – 487.
61
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
ДВА ПОДХОДА МАТЕМАТИЧЕСКОГО МОДЕЛИРОВАНИЯ КОНКУРЕНТНЫХ ОТНОШЕНИЙ ДЕРЕВЬЕВ ДУБА ЧЕРЕШЧАТОГО
Жиренко Н.Г.
Институт лесоведения РАН, Москва, Россия nzhirenko@mail.ru
Аннотация: на примере выявленных конкурентных отношений двух близко расположенных деревьев дуба черешчатого рассмотрены два подхода математического моделирования этого процесса.
Множество происходящих в природе процессов носит колебательный характер и подчиняется экспоненциальным законам. Рассмотрим два подхода, которые можно использовать при математическом описании означенных процессов, на примере выявленных нами конкурентных отношений двух деревьев дуба.
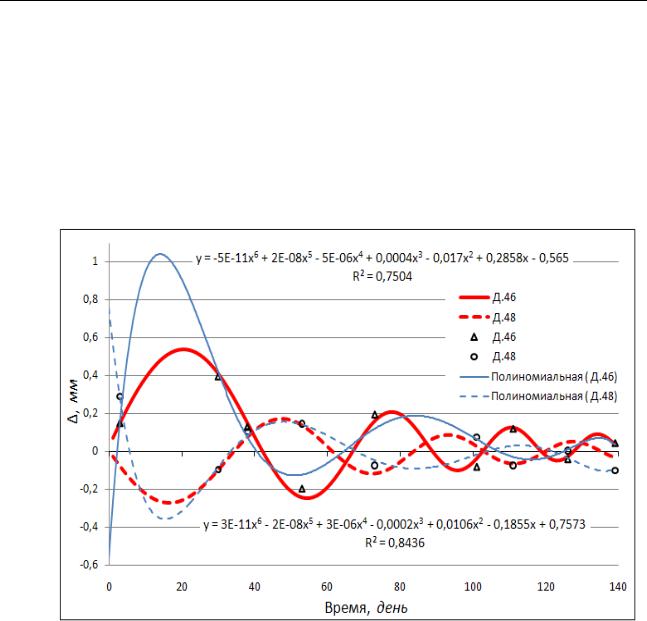
Исследования проводились в 2012 г. в течение вегетационного периода в Теллермановском лесном массиве (Воронежская обл.) и заключались в изучении радиального прироста ствола двух близко расположенных друг к другу деревьев дуба черешчатого IV класса возраста (деревья Д.46 и Д.48). Для этого, на стволы деревьев с южной стороны на высоте ~1.3 м были установлены разработанные нами дендрометры. Конструкция дендрометров и методика их установки позволяют определять изменения толщины заболонного слоя и вторичной коры. В свою очередь изменение толщины заболонного слоя и вторичной коры зависит от их прироста, температуры и влажности. Кроме этого, толщина заболонного слоя может меняться из-за различного сжатия водопроводящих сосудов, в результате возникающего в них отрицательного давления при транспирации. Таким образом, изменение обозначенной толщины может отображать текущее состояние дерева. По показаниям дендрометров были определены изменения обозначенной толщины (∆), которые показаны на рисунке 1 в виде соответствующих графических символов. Времяисчисление (t), в днях, начинается с 19.06.2012 г.
Одним из подходов при математическом описании полученных зависимостей, например, с использованием средств MS Excel, является аппроксимация экспериментальных данных путем построения их диаграммы с последующим подбором наиболее подходящей функции с помощью пункта программы "Добавить линию тренда". В результате этих действий на диаграмму можно вывести полученные уравнения, а также величину достоверности аппроксимации R2. На рисунке 1 результаты аппроксимации представлены полиноминальными уравнениями и их графиками. Уравнение для Д.46 расположено в верхней части диаграммы, а для Д.48 – в нижней. Коэффициенты детерминации в обоих случаях, с точностью до десятых, равны 0.8. Это говорит об удовлетворительной аппроксимации, т.е. формулы в целом адекватны эмпирическим зависимостям. Тем не менее, не смотря на достаточно приемлемые результаты, очевидна громоздкость полученных уравнений. Вследствие этого, проведение анализа, например, с целью выявления отличительных признаков каких-либо других аналогичных процессов, будет достаточно затруднительным.
Рассмотрим второй подход. Как видно из рисунка 1, полученные графические зависимости носят колебательный характер. Следовательно, можно предположить, что исследуемая величина изменяется со временем по закону синуса (или косинуса). Кроме этого, по характеру уменьшения амплитуды колебаний с течением времени, можно предположить, что процесс является затухающим. Как правило, затухание колебаний происходит по экспоненциальному закону и описывается уравнением:
∆=Аsin(wt+ψ)exp(-at) (1)
где А – начальная амплитуда колебания, ψ – начальная фаза колебания, а – коэффициент
62
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
затухания. И еще одним отличительным моментом является то, что частота колебаний полученных зависимостей с течением времени увеличивается. Предположим, что увеличение частоты также происходит по экспоненциальному закону. Введем этот параметр в уравнение 1. Окончательно получим:
∆=Аsin(wtexp(tk)+ψ)exp(-at) (2)
где k – коэффициент, определяющий изменение частоты.
Конкурентные отношения исследуемых деревьев, как видно из графиков (рисунок 1), проявляются в том, что колебательные процессы изменения обозначенной толщины у деревьев происходят в противофазе. Следовательно, колебательный процесс Д.48 будет отставать по фазе от такового Д.46 на π (3.14 рад).
Рисунок 1 - Зависимости изменений толщины заболони и первичной коры деревьев дуба черешчатого (пояснения в тексте)
Воспользовавшись приемами аппроксимации экспериментальных данных и пакетом анализа "Поиск решения" в среде MS Excel не сложно подобрать все параметры уравнения 2. В итоге для исследуемых деревьев Д.46 и Д.48 мы получили следующие уравнения:
∆46 = 0.77·sin(0.06·t·exp(t·0.007))·exp(-0.02·t)+0.03, R2=0.97 (3) ∆48 = 0.36·sin(0.08·t·exp(t·0.004)+π)·exp(-0.02·t), R2=0.83
(4)
Интерпретацию полученных результатов можно представить следующим образом. Введение в уравнение 4 значения π говорит о том, что взаимодействие определенных процессов у рассматриваемых деревьев, например, определяющих их состояние, находится в противофазе. Т.е. когда одно из деревьев находится в лучшем состоянии, то другое, наоборот, оказывается в худшем. Наибольшее значение амплитуды колебания наблюдается у Д.46. В первую очередь, это связано с большими приростами древесины у Д.46 в сравнении с Д.48. О лучшем состоянии Д.46 можно судить и по несколько меньшей частоте колебательного процесса (w46=0.06 рад/с) по сравнению с Д.48 (w48=0.08 рад/с). Это говорит о том, что Д.46 является более устойчивым к факторам окружающей среды. Увеличение же
63

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
частоты со временем у Д.46 (k46=0.007) в сравнении с Д.48 (k48=0.004) может быть связано с ухудшением состояния Д.46 в дальнейшем. По коэффициентам затухания а (коэффициенты равны) можно характеризовать естественное замедление физиологических процессов у растений к концу вегетационного сезона. Графики, построенные по уравнениям 3 и 4, показаны на рисунке 1 красным цветом.
Оба подхода приемлемы для целей математического моделирования. Однако, второй подход, в сравнении с первым, является намного информативнее. Кроме этого, наряду с возможностью сравнения сходных процессов, его применение дает возможность давать достаточно полные характеристики и отдельно взятого исследуемого процесса. Мы предполагаем, что в таком ракурсе взаимоотношение между конкурирующими растениями представляется впервые.
64